
MiRNAs Action and Impact on Mitochondria Function, Metabolic Reprogramming and Chemoresistance of Cancer Cells: A Systematic Review

 , and
, and
Abstract
:1. Introduction
2. Method
2.1. Data Sources and Search Strategy
2.2. Study Selection and Eligibility Criteria
2.3. Data Extraction
2.4. Quality and Bias Evaluation
3. Results and Discussion
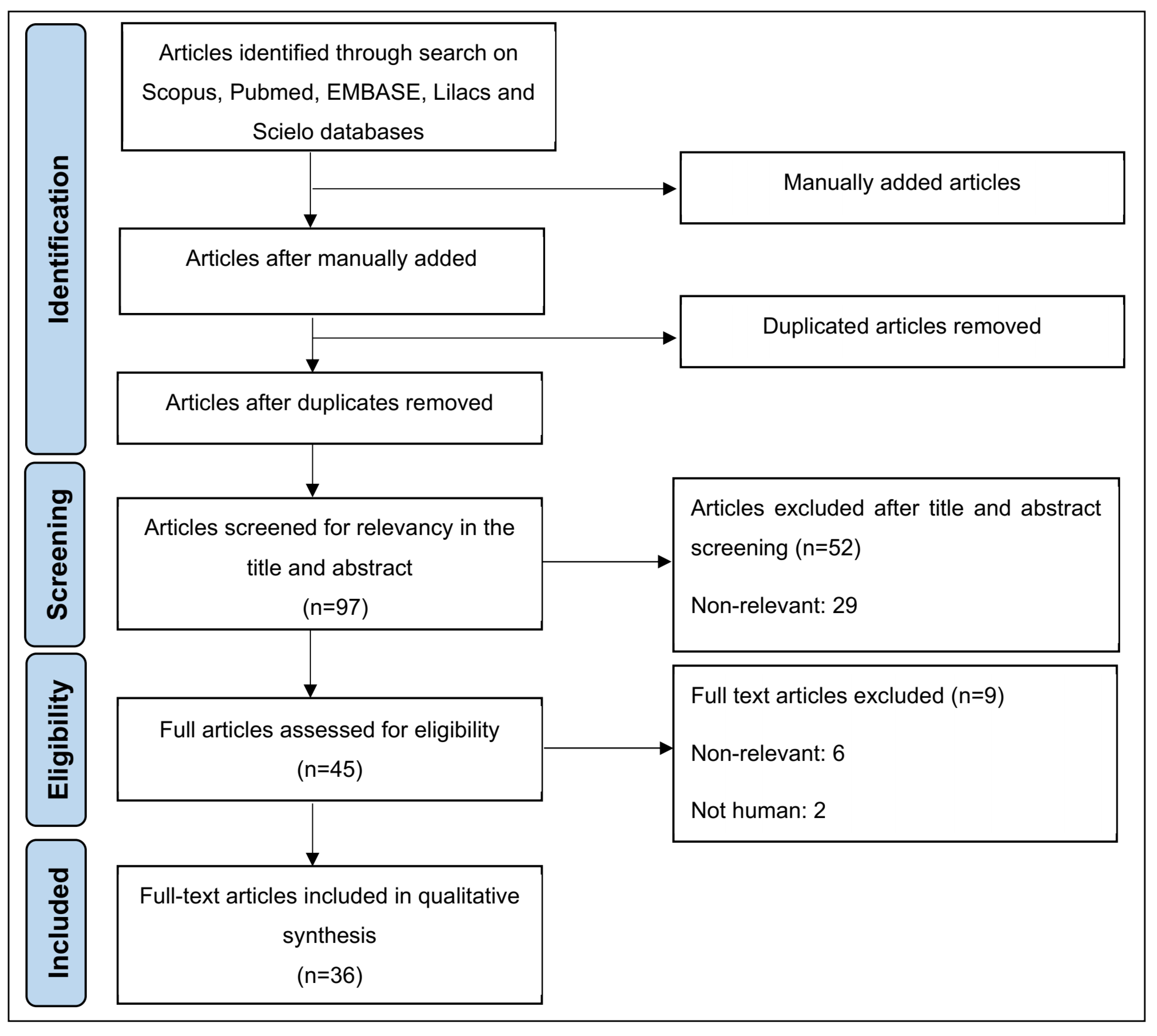
3.1. Thirty-Six Articles Were Identified and Selected Following the PRISMA Guidelines
3.2. Most of the Articles Selected Presented a Low Risk of Bias
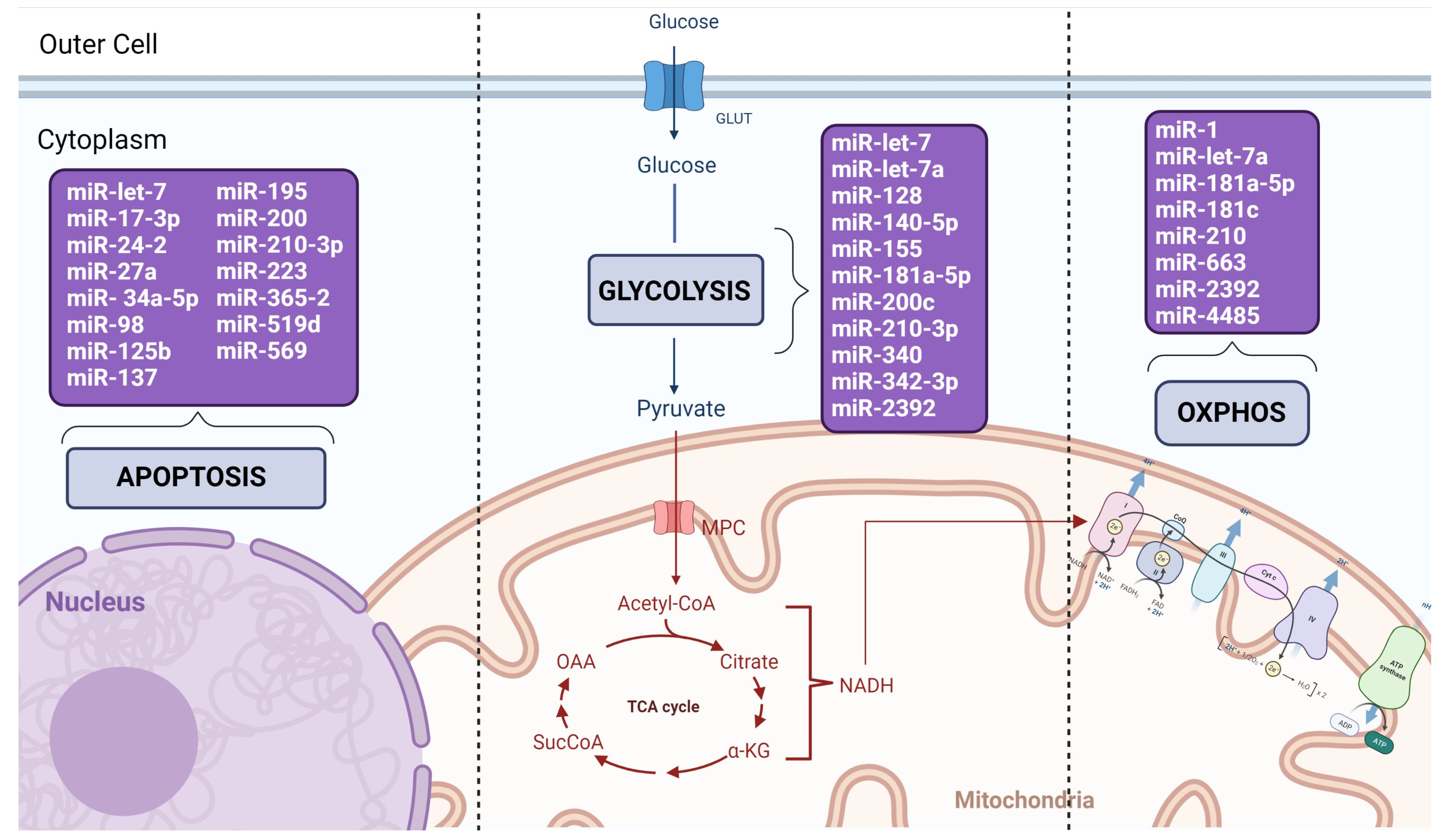
3.3. Fifty-One miRNAs Were Described in 12 Types of Cancer
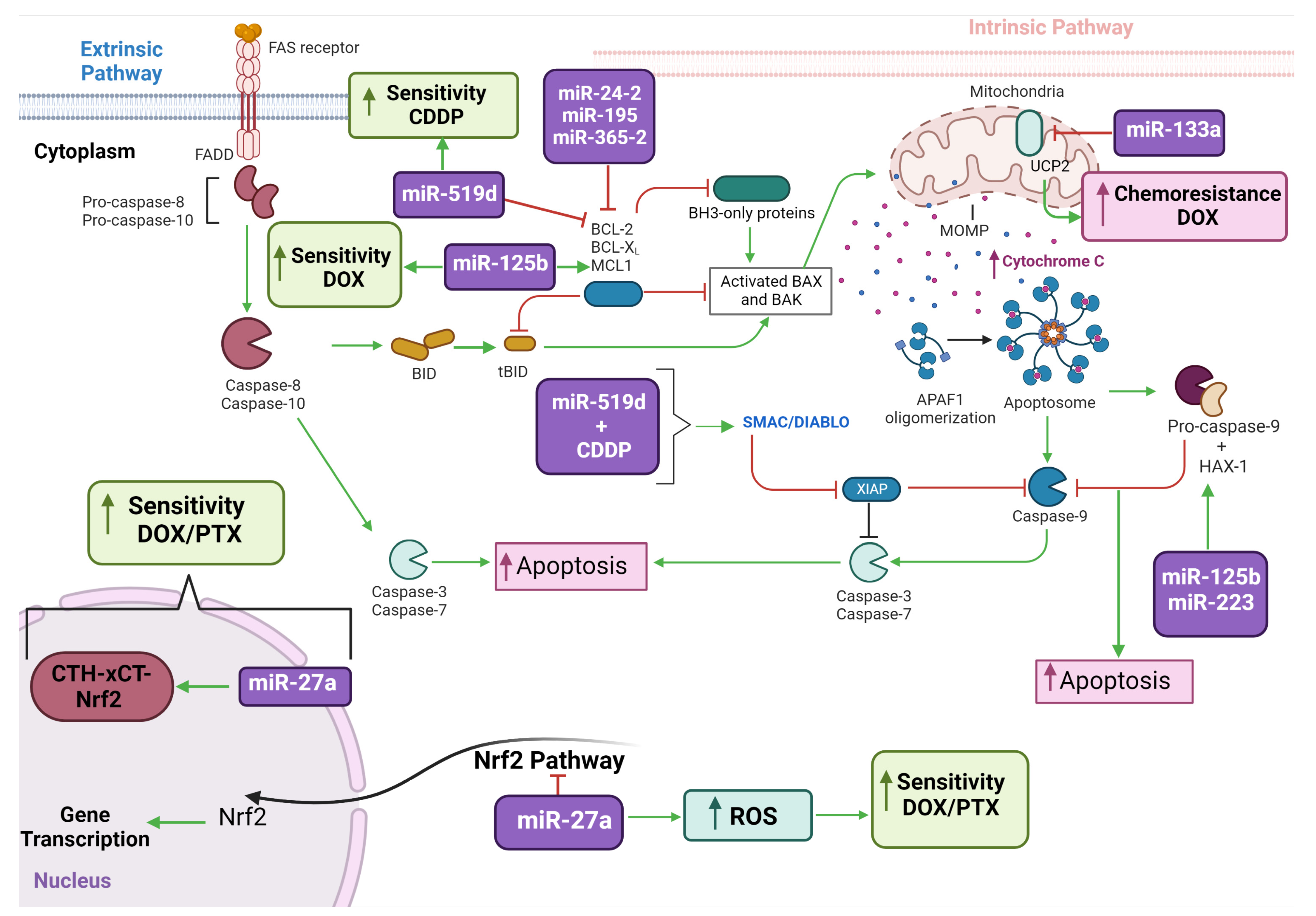
3.4. Thirty-One miRNAs Were Described with Oncogenic Mode of Action Mediating Metabolic Reprogramming and Mitochondria Related Functions
3.5. Twenty-One miRNAs Were Described with Suppressive Mode of Action Mediating Metabolic Reprogramming and Mitochondria Related Functions
3.6. Nine miRNAs Were Identified Acting on Tumor Chemoresistance Mediating Metabolic Reprogramming and Mitochondria Related Functions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, I.; Ajay, S.S.; Yook, J.I.; Kim, H.S.; Hong, S.H.; Kim, N.H.; Dhanasekaran, S.M.; Chinnaiyan, A.M.; Athey, B.D. New class of microRNA targets containing simultaneous 5′-UTR and 3′-UTR interaction sites. Genome Res. 2009, 19, 1175–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, M.; Li, J.; Li, W.; Wang, Y.; Wu, F.; Xi, Y.; Zhang, L.; Ding, C.; Luo, H.; Li, Y.; et al. MicroRNAs activate gene transcription epigenetically as an enhancer trigger. RNA Biol. 2017, 14, 1326–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheu-Gruttadauria, J.; Xiao, Y.; Gebert, L.F.; MacRae, I.J. Beyond the seed: Structural basis for supplementary microRNA targeting by human Argonaute2. EMBO J. 2019, 38, e101153. [Google Scholar] [CrossRef]
- Tie, J.; Fan, D. Big roles of microRNAs in tumorigenesis and tumor development. Histol. Histopathol. 2011, 26, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhang, L.; Li, Z.; Li, Y.; Huang, C.; Lu, X. MicroRNAs in DNA Damage Response, Carcinogenesis, and Chemoresistance. Int. Rev. Cell Mol. Biol. 2017, 333, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Madhumita, M.; Paul, S. A review on methods for predicting miRNA-mRNA regulatory modules. J. Integr. Bioinform. 2022, 19. [Google Scholar] [CrossRef]
- Porporato, P.E.; Filigheddu, N.; Pedro, J.M.B.; Kroemer, G.; Galluzzi, L. Mitochondrial metabolism and cancer. Cell Res. 2018, 28, 265–280. [Google Scholar] [CrossRef]
- Lin, H.Y.; Chu, P.Y. Advances in Understanding Mitochondrial MicroRNAs (mitomiRs) on the Pathogenesis of Triple-Negative Breast Cancer (TNBC). Oxid. Med. Cell. Longev. 2021, 2021, 5517777. [Google Scholar] [CrossRef]
- Purohit, P.K.; Saini, N. Mitochondrial microRNA (MitomiRs) in cancer and complex mitochondrial diseases: Current status and future perspectives. Cell. Mol. Life Sci. 2021, 78, 1405–1421. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, M.; Newport, E.; Morten, K.J. The Warburg effect: 80 years on. Biochem. Soc. Trans. 2016, 44, 1499–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Palm, W. Metabolic plasticity allows cancer cells to thrive under nutrient starvation. Proc. Natl. Acad. Sci. USA 2021, 118, e2102057118. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Xiang, K.; Jendrossek, V.; Matschke, J. Oncometabolites and the response to radiotherapy. Radiat. Oncol. 2020, 15, 197. [Google Scholar] [CrossRef]
- Kamphorst, J.J.; Cross, J.R.; Fan, J.; de Stanchina, E.; Mathew, R.; White, E.P.; Thompson, C.B.; Rabinowitz, J.D. Hypoxic and Ras-transformed cells support growth by scavenging unsaturated fatty acids from lysophospholipids. Proc. Natl. Acad. Sci. USA 2013, 110, 8882–8887. [Google Scholar] [CrossRef] [Green Version]
- Rabinowitz, J.D.; White, E. Autophagy and metabolism. Science 2010, 330, 1344–1348. [Google Scholar] [CrossRef] [Green Version]
- Vyas, S.; Zaganjor, E.; Haigis, M.C. Mitochondria and Cancer. Cell 2016, 166, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Faubert, B.; Solmonson, A. Metabolic reprogramming and cancer progression. Science 2020, 368, eaaw5473. [Google Scholar] [CrossRef]
- Missiroli, S.; Genovese, I.; Perrone, M.; Vezzani, B.; Vitto, V.A.M.; Giorgi, C. The Role of Mitochondria in Inflammation: From Cancer to Neurodegenerative Disorders. J. Clin. Med. 2020, 9, 740. [Google Scholar] [CrossRef] [Green Version]
- Barrey, E.; Saint-Auret, G.; Bonnamy, B.; Damas, D.; Boyer, O.; Gidrol, X. Pre-microRNA and mature microRNA in human mitochondria. PLoS ONE 2011, 6, e20220. [Google Scholar] [CrossRef]
- Bandiera, S.; Rüberg, S.; Girard, M.; Cagnard, N.; Hanein, S.; Chrétien, D.; Munnich, A.; Lyonnet, S.; Henrion-Caude, A. Nuclear outsourcing of RNA interference components to human mitochondria. PLoS ONE 2011, 6, e20746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandiera, S.; Matégot, R.; Girard, M.; Demongeot, J.; Henrion-Caude, A. MitomiRs delineating the intracellular localization of microRNAs at mitochondria. Free. Radic. Biol. Med. 2013, 64, 12–19. [Google Scholar] [CrossRef]
- Macgregor-Das, A.M.; Das, S. A microRNA’s journey to the center of the mitochondria. Am. J. Physiol. Circ. Physiol. 2018, 315, H206–H215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, D.L.; Hathaway, Q.A.; Pinti, M.V.; Nichols, C.E.; Durr, A.J.; Sreekumar, S.; Hughes, K.M.; Stine, S.M.; Martinez, I.; Hollander, J.M. Exploring the mitochondrial microRNA import pathway through Polynucleotide Phosphorylase (PNPase). J. Mol. Cell. Cardiol. 2017, 110, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latronico, M.V.; Condorelli, G. The might of microRNA in mitochondria. Circ. Res. 2012, 110, 1540–1542. [Google Scholar] [CrossRef] [Green Version]
- Sripada, L.; Tomar, D.; Singh, R. Mitochondria: One of the destinations of miRNAs. Mitochondrion 2012, 12, 593–599. [Google Scholar] [CrossRef]
- Simão, A.L.; Borralho, P.M.; Steer, C.J.; Castro, R.E.; Rodrigues, C.M.P. Isolation of Mitochondria from Liver and Extraction of Total RNA and Protein: Analyses of miRNA and Protein Expression. Methods Mol. Biol. 2021, 2310, 1–15. [Google Scholar] [CrossRef]
- Borralho, P.M.; Rodrigues, C.M.; Steer, C.J. MicroRNAs in Mitochondria: An Unexplored Niche. Adv. Exp. Med. Biol. 2015, 887, 31–51. [Google Scholar] [CrossRef]
- Duarte, F.V.; Palmeira, C.M.; Rolo, A.P. The Role of microRNAs in Mitochondria: Small Players Acting Wide. Genes 2014, 5, 865–886. [Google Scholar] [CrossRef] [Green Version]
- Geiger, J.; Dalgaard, L.T. Interplay of mitochondrial metabolism and microRNAs. Cell. Mol. Life Sci. 2017, 74, 631–646. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Bharat; Dogra, N.; Singh, S. Small Regulatory Molecules Acting Big in Cancer: Potential Role of Mito-miRs in Cancer. Curr. Mol. Med. 2019, 19, 621–631. [Google Scholar] [CrossRef]
- Rencelj, A.; Gvozdenovic, N.; Cemazar, M. MitomiRs: Their roles in mitochondria and importance in cancer cell metabolism. Radiol. Oncol. 2021, 55, 379–392. [Google Scholar] [CrossRef]
- Kuthethur, R.; Shukla, V.; Mallya, S.; Adiga, D.; Kabekkodu, S.P.; Ramachandra, L.; Saxena, P.U.P.; Satyamoorthy, K.; Chakrabarty, S. Expression analysis and function of mitochondrial genome-encoded microRNAs. J. Cell Sci. 2022, 135, jcs258937. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Tian, T.; Chen, W.; Lv, X.; Lei, X.; Zhang, H.; Sun, S.; Cai, L.; Pan, G.; He, L.; et al. Mitochondrial miRNA Determines Chemoresistance by Reprogramming Metabolism and Regulating Mitochondrial Transcription. Cancer Res. 2019, 79, 1069–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khot, V.M.; Salunkhe, A.B.; Pricl, S.; Bauer, J.; Thorat, N.D.; Townley, H. Nanomedicine-driven molecular targeting, drug delivery, and therapeutic approaches to cancer chemoresistance. Drug Discov. Today 2021, 26, 724–739. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Kosnacova, H. Mitochondrial Genetic and Epigenetic Regulations in Cancer: Therapeutic Potential. Int. J. Mol. Sci. 2022, 23, 7897. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Hayden, J.A.; Côté, P.; Bombardier, C. Evaluation of the quality of prognosis studies in systematic reviews. Ann. Intern. Med. 2006, 144, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Serguienko, A.; Grad, I.; Wennerstrøm, A.B.; Meza-Zepeda, L.A.; Thiede, B.; Stratford, E.W.; Myklebost, O.; Munthe, E. Metabolic reprogramming of metastatic breast cancer and melanoma by let-7a microRNA. Oncotarget 2015, 6, 2451–2465. [Google Scholar] [CrossRef] [Green Version]
- Ueda, S.; Takanashi, M.; Sudo, K.; Kanekura, K.; Kuroda, M. miR-27a ameliorates chemoresistance of breast cancer cells by disruption of reactive oxygen species homeostasis and impairment of autophagy. Lab. Investig. J. Tech. Methods Pathol. 2020, 100, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Zhao, X.; Wang, J.; Lv, L.; Wang, C.; Feng, L.; Shen, L.; Ren, W. miR-125b regulates the drug-resistance of breast cancer cells to doxorubicin by targeting HAX-1. Oncol. Lett. 2018, 15, 1621–1629. [Google Scholar] [CrossRef]
- Xie, X.; Hu, Y.; Xu, L.; Fu, Y.; Tu, J.; Zhao, H.; Zhang, S.; Hong, R.; Gu, X. The role of miR-125b-mitochondria-caspase-3 pathway in doxorubicin resistance and therapy in human breast cancer. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2015, 36, 7185–7194. [Google Scholar] [CrossRef]
- Yuan, Y.; Yao, Y.F.; Hu, S.N.; Gao, J.; Zhang, L.L. MiR-133a Is Functionally Involved in Doxorubicin-Resistance in Breast Cancer Cells MCF-7 via Its Regulation of the Expression of Uncoupling Protein 2. PLoS ONE 2015, 10, e0129843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Cordoba, S.L.; Rodriguez-Cuevas, S.; Bautista-Pina, V.; Maffuz-Aziz, A.; D’Ippolito, E.; Cosentino, G.; Baroni, S.; Iorio, M.V.; Hidalgo-Miranda, A. Loss of function of miR-342-3p results in MCT1 over-expression and contributes to oncogenic metabolic reprogramming in triple negative breast cancer. Sci. Rep. 2018, 8, 12252. [Google Scholar] [CrossRef] [Green Version]
- Luan, T.; Fu, S.; Huang, L.; Zuo, Y.; Ding, M.; Li, N.; Chen, J.; Wang, H.; Wang, J. MicroRNA-98 promotes drug resistance and regulates mitochondrial dynamics by targeting LASS2 in bladder cancer cells. Exp. Cell Res. 2018, 373, 188–197. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, Q.; Zheng, S.; Lin, K.; You, J.; Zhang, X. miR-27a regulates the sensitivity of breast cancer cells to cisplatin treatment via BAK-SMAC/DIABLO-XIAP axis. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2016, 37, 6837–6845. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, Y.; Zhang, H.; Huang, P.; Luthra, R. Hypoxia-regulated microRNA-210 modulates mitochondrial function and decreases ISCU and COX10 expression. Oncogene 2010, 29, 4362–4368. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Liu, B.; Sun, L.; Lv, X.B.; Lin, Z.; Chen, W.; Chen, W.; Tang, Q.; Wang, Y.; Su, Y.; et al. Mitochondrial fission determines cisplatin sensitivity in tongue squamous cell carcinoma through the BRCA1-miR-593-5p-MFF axis. Oncotarget 2015, 6, 14885–14904. [Google Scholar] [CrossRef] [Green Version]
- Wallace, L.; Aikhionbare, K.; Banerjee, S.; Peagler, K.; Pitts, M.; Yao, X.; Aikhionbare, F. Differential Expression Profiles of Mitogenome Associated MicroRNAs Among Colorectal Adenomatous Polyps. Cancer Res. J. 2021, 9, 23–33. [Google Scholar] [CrossRef]
- Sharma, P.; Sharma, V.; Ahluwalia, T.S.; Dogra, N.; Kumar, S.; Singh, S. Let-7a induces metabolic reprogramming in breast cancer cells via targeting mitochondrial encoded ND4. Cancer Cell Int. 2021, 21, 629. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, C.; Zhang, X. Mitochondrial Damage Mediated by miR-1 Overexpression in Cancer Stem Cells. Mol. Ther. Nucleic Acids 2019, 18, 938–953. [Google Scholar] [CrossRef] [Green Version]
- Purohit, P.K.; Edwards, R.; Tokatlidis, K.; Saini, N. MiR-195 regulates mitochondrial function by targeting mitofusin-2 in breast cancer cells. RNA Biol. 2019, 16, 918–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Zhou, E.; Wang, Y.; Xu, F.; Zhang, D.; Zhong, D. microRNA-200a inhibits cell proliferation by targeting mitochondrial transcription factor A in breast cancer. DNA Cell Biol. 2014, 33, 291–300. [Google Scholar] [CrossRef]
- Carden, T.; Singh, B.; Mooga, V.; Bajpai, P.; Singh, K.K. Epigenetic modification of miR-663 controls mitochondria-to-nucleus retrograde signaling and tumor progression. J. Biol. Chem. 2017, 292, 20694–20706. [Google Scholar] [CrossRef] [Green Version]
- Sripada, L.; Singh, K.; Lipatova, A.V.; Singh, A.; Prajapati, P.; Tomar, D.; Bhatelia, K.; Roy, M.; Singh, R.; Godbole, M.M.; et al. hsa-miR-4485 regulates mitochondrial functions and inhibits the tumorigenicity of breast cancer cells. J. Mol. Med. 2017, 95, 641–651. [Google Scholar] [CrossRef]
- Hu, Q.; Yuan, Y.; Wu, Y.; Huang, Y.; Zhao, Z.; Xiao, C. MicroRNA-137 exerts protective effects on hypoxia-induced cell injury by inhibiting autophagy/mitophagy and maintaining mitochondrial function in breast cancer stem-like cells. Oncol. Rep. 2020, 44, 1627–1637. [Google Scholar] [CrossRef]
- Jung, K.A.; Lee, S.; Kwak, M.K. NFE2L2/NRF2 Activity Is Linked to Mitochondria and AMP-Activated Protein Kinase Signaling in Cancers Through miR-181c/Mitochondria-Encoded Cytochrome c Oxidase Regulation. Antioxid. Redox Signal. 2017, 27, 945–961. [Google Scholar] [CrossRef]
- Yi, W.R.; Tu, M.J.; Yu, A.X.; Lin, J.; Yu, A.M. Bioengineered miR-34a modulates mitochondrial inner membrane protein 17 like 2 (MPV17L2) expression toward the control of cancer cell mitochondrial functions. Bioengineered 2022, 13, 12489–12503. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Lou, C.; Xiao, H.; Yang, Y.; Cai, X.; Li, C.; Jia, S.; Huang, Y. MiR-128 regulation of glucose metabolism and cell proliferation in triple-negative breast cancer. Br. J. Surg. 2018, 105, 75–85. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Deng, F.; Zhao, S.; Zhong, S.; Zhao, J.; Wang, D.; Chen, X.; Zhang, J.; Hou, J.; Zhang, W.; et al. Analysis of miRNA-mRNA network reveals miR-140-5p as a suppressor of breast cancer glycolysis via targeting GLUT1. Epigenomics 2019, 11, 1021–1036. [Google Scholar] [CrossRef]
- Kim, S.; Lee, E.; Jung, J.; Lee, J.W.; Kim, H.J.; Kim, J.; Yoo, H.J.; Lee, H.J.; Chae, S.Y.; Jeon, S.M.; et al. MicroRNA-155 positively regulates glucose metabolism via PIK3R1-FOXO3a-cMYC axis in breast cancer. Oncogene 2018, 37, 2982–2991. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Wei, N.; Ma, R.; Jiang, S.; Song, D. A miR-210-3p regulon that controls the Warburg effect by modulating HIF-1α and p53 activity in triple-negative breast cancer. Cell Death Dis. 2020, 11, 731. [Google Scholar] [CrossRef]
- Zhuang, X.; Chen, Y.; Wu, Z.; Xu, Q.; Chen, M.; Shao, M.; Cao, X.; Zhou, Y.; Xie, M.; Shi, Y.; et al. Mitochondrial miR-181a-5p promotes glucose metabolism reprogramming in liver cancer by regulating the electron transport chain. Carcinogenesis 2020, 41, 972–983. [Google Scholar] [CrossRef]
- Sripada, L.; Tomar, D.; Prajapati, P.; Singh, R.; Singh, A.K.; Singh, R. Systematic analysis of small RNAs associated with human mitochondria by deep sequencing: Detailed analysis of mitochondrial associated miRNA. PLoS ONE 2012, 7, e44873. [Google Scholar] [CrossRef]
- Singh, R.; Yadav, V.; Kumar, S.; Saini, N. MicroRNA-195 inhibits proliferation, invasion and metastasis in breast cancer cells by targeting FASN, HMGCR, ACACA and CYP27B1. Sci. Rep. 2015, 5, 17454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.; Wang, Y.; Peng, J.; Shen, Q.; Chen, M.; Tang, W.; Li, X.; Cai, C.; Wang, B.; Cai, S.; et al. Mitochondrial calcium uniporter as a target of microRNA-340 and promoter of metastasis via enhancing the Warburg effect. Oncotarget 2017, 8, 83831–83844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhou, Y.; Han, F.; Zhao, Y.; Tu, M.; Wang, Y.; Huang, C.; Fan, S.; Chen, P.; Yao, X.; et al. A novel miR-1291-ERRα-CPT1C axis modulates tumor cell proliferation, metabolism and tumorigenesis. Theranostics 2020, 10, 7193–7210. [Google Scholar] [CrossRef]
- Xu, Y.; Fang, F.; Zhang, J.; Josson, S.; St. Clair, W.H.; St. Clair, D.K. miR-17* suppresses tumorigenicity of prostate cancer by inhibiting mitochondrial antioxidant enzymes. PLoS ONE 2010, 5, e14356. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, Y.; Ding, J.; Hu, W.; Tan, C.; Wang, M.; Tang, J.; Xu, Y. miR-17-3p Downregulates Mitochondrial Antioxidant Enzymes and Enhances the Radiosensitivity of Prostate Cancer Cells. Mol. Therapy. Nucleic Acids 2018, 13, 64–77. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Li, Y.; Zheng, M.; Zuo, W.; Zheng, W. MicroRNA-223 Increases the Sensitivity of Triple-Negative Breast Cancer Stem Cells to TRAIL-Induced Apoptosis by Targeting HAX-1. PLoS ONE 2016, 11, e0162754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Q.; Wang, S.; Zhao, Y.; Zhang, Z.; Qin, C.; Yang, X. MiR-519d impedes cisplatin-resistance in breast cancer stem cells by down-regulating the expression of MCL-1. Oncotarget 2017, 8, 22003–22013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Saini, N. Downregulation of BCL2 by miRNAs augments drug-induced apoptosis-a combined computational and experimental approach. J. Cell Sci. 2012, 125, 1568–1578. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.Q.; Wang, S.Q.; Chen, Y.; Fu, L.Y.; Xu, Y.N.; Li, L.; Tao, L.; Shen, X.C. MicroRNAs Regulating Mitochondrial Function in Cardiac Diseases. Front. Pharmacol. 2021, 12, 663322. [Google Scholar] [CrossRef]
- O’Bryan, S.; Dong, S.; Mathis, J.M.; Alahari, S.K. The roles of oncogenic miRNAs and their therapeutic importance in breast cancer. Eur. J. Cancer 2017, 72, 1–11. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| MiRNA | Target Gene | Cancer Type | Mechanism of Action | Biological Impact | Reference |
|---|---|---|---|---|---|
| miR-98 | LASS2 | Bladder cancer | ↑miR-98 ↓LASS2→↓Mitochondrial fusion ↑Mitochondrial membrane potential | ↓Apoptosis ↑Chemoresistance | [50] |
| miR-27a | BAK-SMAC/DIABLO/XIAP | Breast cancer | ↑miR-27a→↓BAK→↓SMAC/DIABLO/XIAP | ↑Colony formation ↓Chemosensitivity ↑Metastasis ↓ Apoptosis | [51] |
| miR-137 | FUNDCc1 | Breast cancer | ↑miR-137→ ↓FUNDC1→↓ROS | ↓Apoptosis ↓Autophagy/Mitophagy ↑Cell cycle | [61] |
| miR-155 | PIK3R1-FOXO3a-cMYC axis; p85α-FOXO3a-cMYC | Breast cancer | ↑miR-155→↓PIK3R1-FOXO3a-cMYC ↑miR-155→↓p85α-FOXO3a-cMYC | ↑Glucose and glycolysis ↑Tumor growth | [66] |
| miR-210-3p | CYGB GPD1L HIF-1α and p53 activity via GPD1L and CYGB | Breast cancer | ↑miR-210-3p ↓CYGB ↓GPD1L ↑HIF-1α↓p53→↑Aerobic glycolysis ↓Apoptosis | ↑Colony formation, ↑Extracellular acidification rate ↑Glucose uptake ↑Lactate production ↓Serum starvation-induced cell apoptosis. | [67] |
| miR-let-7i miR-21 miR-23a miR-29a miR-30a miR-31 miR-181b miR-452 | Not described | Cervical cell line (HeLa) | ↑miR-let-7i ↑miR-21 ↑miR-23a ↑miR-29a ↑miR-30a ↑miR-31 ↑miR-181b ↑miR-452 | Systematic analysis shows a significant overexpression of these miRNAs and their enrichment in the mitochondrial RNA fraction | [69] |
| miR-328 miR-494 miR-513a-5p miR-638 miR-1201 miR-1246 miR-1275 miR-1908 miR-1972 | Not described | Cervical cell line (HeLa) | ↑miR-328 ↑miR-494 ↑miR-513a-5p ↑miR-638 ↑miR-1201 ↑miR-1246 ↑miR-1275 ↑miR-1908 ↑miR-1972 | Systematic analysis shows a significant overexpression of these miRNAs and their enrichment in the mitochondrial RNA fraction | [25] |
| miR-181c | AMPKa mt-CO1 | Colon cancer | ↑miR-181c ↓mt-CO1 ↑MMP disturbance and ETC dysfunction→↓levels of ATP ↑AMPKa | ↓Mitochondrial OCR - OXPHOS (NFE2L2/NRF2- knockdown colon cancer cells) | [62] |
| miR-210 | COX10 ISCU | Colon cancer | ↑ miR-210 ↓COX10/ISCU ↑ROS | ↑Cell survival under hypoxic microenvironment | [52] |
| miR-21 miR-24 miR-181 miR-210 miR-378 | Not described | Colorectal cancer | ↑ miR-21 ↑miR-24 ↑miR-181 ↑miR-210 ↑miR-378 | ↑Transformation of precancerous polyps to CRC adenocarcinoma | [54] |
| miR-181a-5p | mt-CO2 mt-CYB | Liver cancer | ↑miR-181a-5p ↓mt-CYB ↓mt-CO2 →ETC remodeling →↓OXPHOS ↑GLUT1 ↑HK2 | ↑Cell survival under hypoxic microenvironment ↑Glucose consumption ↑Lactic acid secretion ↑LDH | [68] |
| miR-593-5p | MFF | Tongue cancer | ↑BRCA1→↑miR-593-5p→ ↓MFF | ↓Cisplatin sensitivity ↑Mitochondrial fission | [53] |
| miR-2392 | AGO2 | Tongue cancer | ↑miR-2392 ↑AGO2 →↓OXPHOS ↑Glycolysis | ↑Chemoresistance ↑Glycolysis | [38] |
| MiRNA | Target Gene | Cancer Type | Mechanism of Action | Biological Impact | Reference |
|---|---|---|---|---|---|
| let-7a | ND4 | Breast cancer | ↑miR-let-7a ↓ND4 → ↓OXPHOS | ↓Adipogenesis ↑Lactate concentration (glycolysis) | [55] |
| let-7a | AASDHPPT BACH1 FASN G6PD HMOX1 IMPDH2 SCD | Breast cancer and melanoma | ↑miR-let-7a → ↓AASDHPPT ↓BACH1 ↓FASN ↓G6PD ↓SCD ↓IMPDH2 ↑HMOX1 → ↑ROS ↑OXPHOS and glycolysis | ↑Chemosensitivity ↓Proliferation | [44] |
| miR-1 | ATP6 COX1 GPD2 LRPPRC MINOS1 ND1 ND4 | Breast cancer and melanoma | ↑miR-1 ↓ ATP6 ↓COX1, ↓ GPD2 ↓ LRPPRC ↓MINOS1 ↓ND1 ↓ ND4 | ↑Mitochondrial cristae organization and morphology ↓Tumorigenicity | [56] |
| miR-24-2 miR-195 miR-365-2 | BCL2 | Breast cancer | ↑miR-24-2 ↑miR-195 ↑miR-365-2 ↓BCL2 ↓ΔΨm ↑cytochrome c protein (in the cytosol) | ↑Apoptosis | [77] |
| miR-27a | CTH NFE2L2 SLC7A11 | Breast cancer | ↑miR-27a ↓CTH ↓xCT ↓NRF2↑ROS | ↓Autophagy ↓Chemoresistance | [45] |
| miR-125b | HAX-1 | Breast cancer | ↑ miR-125b + DOX → ↓HAX-1 ↑caspase-9 ↑ROS | ↑Apoptosis ↑Cell death ↑ Chemosensitivity ↑Mitochondrial damage | [46] |
| miR-125b | MCL-1 | Breast cancer | ↑miR-125b ↓MCL-1→ ↑caspase-3 | ↑Apoptosis ↓Doxorubicin resistance | [47] |
| miR-128 | INSR IRS1 | Breast cancer | ↑miR-128 ↓INSR ↓IRS1→↓mtDNA | ↓Glucose metabolism (glycolysis) ↓Proliferation | [64] |
| miR-133a | UCP-2 | Breast cancer | ↑miR 133a → ↓UCP-2 | ↓Chemoresistance | [48] |
| miR-140-5p | GLUT1 | Breast cancer | ↑miR-140-5p ↓GLUT1→↓glycolysis ↑mitochondrial respiration | ↓Proliferation (glycolysis) | [65] |
| miR-195 | ACACA CYP27B1 FASN HMGCR | Breast cancer | ↑miR-195 ↓ ACACA →↓biosynthesis of fatty acid ↓ FASN ↓CYP27B1 → ↓mitochondrial calcium levels ↓HMGCR →↓mevalonate pathway | ↓Fatty acid (via de novo lipogenesis) | [70] |
| miR-195 | MFN2 | Breast cancer | ↑miR-195 ↓MFN2→↓mitochondrial dynamics ↓mitochondrial calcium homeostasis | ↑Apoptosis | [57] |
| miR-200a | TFAM | Breast cancer | ↑miR-200a ↓TFAM→↓mtDNA copy number | ↓Cell proliferation | [58] |
| miR-223 | TRAIL/HAX-1 | Breast cancer | ↑miR-223 ↓TRAIL → ↓HAX-1 | ↑Apoptosis ↑Chemosensitivity | [75] |
| miR-340 | MCU | Breast cancer | ↑miR-340 ↓MCU→↓glycolysis | ↓Metastasis (glycolysis) | [71] |
| miR-342-3p | MCT-1 | Breast cancer | ↑miR-342-3p ↓ MCT-1 → ↓ extracellular lactate ↑glucose consumption | ↓ Lactate transportation to inside cell | [49] |
| miR-519d | MCL-1 | Breast cancer | ↑miR-519d ↑cisplatin → ↓MCL-1 ↑cytochrome C ↑SMAC/DIABLO | ↑Apoptosis ↑ Cell death ↑Chemosensitivity | [76] |
| miR-663 | OXPHOS genes | Breast cancer | ↑miR-663 ↑OXPHOS | ↓Tumorigenesis | [59] |
| miR-1291 | CPT1C ERRα | Breast cancer and pancreatic cancer | ↑miR-1291 ↓ERRα → ↓CPT1C | ↓Invasion ↑Mitochondrial dysfunction ↓ Proliferation ↓Tumorigenesis ↓ Cell metabolism | [72] |
| miR-4485 | 16S rRNA | Breast cancer | ↑miR-4485 ↓ ΔΨm ↓16S rRNA →↓respiratory complex I →↑ROS | ↑Cell death | [60] |
| miR-34a-5p | MPV17L2 | Lung cancer and osteosarcoma | ↓ND1 expression ↓mitochondrial respiration ↓ATP production ↑ROS accumulation | ↑ Apoptotic cell death | [63] |
| miR-17* | GPX2 MnSOD TRXR2 | Prostate cancer | ↑miR-17 ↓Gpx2 ↓MnSOD ↓TRXR2 → ↑ROS | ↑Cytotoxicity ↑Cell death ↓Tumorigenicity | [73] |
| miR-17-3p | GPX2 MnSOD TRXR2 | Prostate cancer | ↑miR-17-3p ↓Gpx2 ↓MnSOD ↓TRXR2 → ↑ROS ↓mitochondrial respiration | ↑Cell death ↑Ionizing radiation ↑Radiosensitivity | [74] |
| MiRNAs | Enzyme Precursor | Metabolic Pathway/Cell Compartment | Reference |
|---|---|---|---|
| let-7a | ND4 | OXPHOS/mitochondria | [55] |
| let-7a | AASDHPPT FASN G6PD HMOX1 IMPDH2 SCD | Pantothenate and CoA biosynthesis/cytosol Fatty acid biosynthesis/cytosol, mitochondria and nucleus Transport reactions/cytosol, extracellular and nucleus Pentose phosphate pathway/cytosol and endoplasmic reticulum Heme degradation/cytosol Porphyrin metabolism/cytosol Purine metabolism/cytosol Fatty acid desaturation/cytosol Fatty acid biosynthesis/cytosol and nucleus Fatty acid desaturation/cytosol Transport reactions—cytosol and nucleus | [44] |
| miR-1 | ATP6 COX1 GPD2 ND1 ND4 | OXPHOS/mitochondria OXPHOS/mitochondria Glycerophospholipid metabolism/cytosol and mitochondria Acylglycerides metabolism/cytosol and mitochondria Glycolysis/Gluconeogenesis/cytosol and mitochondria OXPHOS/ mitochondria OXPHOS/ mitochondria | [56] |
| miR-17* | GPX2 MnSOD TRXR2 | Arachidonic acid metabolism/cytosol, endoplasmic reticulum, extracellular, mitochondria, and peroxisome Glutathione metabolism/cytosol, extracellular and mitochondria Linoleate metabolism/cytosol and endoplasmic reticulum ROS detoxification/cytosol and mitochondria Metabolism of amino acids/cytosol Pyrimidine metabolism/cytosol and mitochondria Cysteine and methionine metabolism/cytosol and mitochondria Nucleotide metabolism/cytosol and mitochondria | [73] |
| miR-17-3p | GPX2 MnSOD TRXR2 | Same as miR-17* | [74] |
| miR-27a | CTH SLC7A11 | Cysteine and methionine metabolism/cytosol Metabolism of other amino acids/cytosol Transport reactions/cytosol and extracellular | [45] |
| miR-98 | LASS2 | Sphingolipid metabolism/cytosol | [50] |
| miR-133a | UCP-2 | Transport reactions/cytosol and mitochondria | [48] |
| miR-140-5p | GLUT1 | Glycosphingolipid metabolism/cytosol, endoplasmic reticulum, and golgi apparatus | [65] |
| miR-155 | PIK3R1 p85α | Inositol phosphate metabolism/cytosol p85Inositol phosphate metabolism/cytosol | [66] |
| miR-181a-5p | mt-CO2 mt-CYB | OXPHOS/mitochondria | [68] |
| miR-181c | mt-CO1 | OXPHOS/mitochondria | [62] |
| miR-195 | ACACA CYP27B1 HMGCR | Biotin metabolism/cytosol and nucleus Transport reactions/cytosol and nucleus Vitamin D metabolism/cytosol and mitochondria Cholesterol metabolism/cytosol, endoplasmic reticulum and peroxisome Cholesterol biosynthesis 1 (Bloch pathway)/cytosol Transport reactions/cytosol, endoplasmic reticulum and peroxisome | [70] |
| miR-210 | COX10 | Histidine metabolism/cytosol | [52] |
| miR-210-3p | GPD1L HIF-1α activity | Glycerophospholipid metabolism/cytosol and peroxisome Transport reactions/cytosol and peroxisome | [67] |
| miR-1291 | CPT1C | Fatty acid oxidation/cytosol and peroxisome Carnitine shuttle/cytosol, endoplasmic reticulum and peroxisome | [72] |
| miR-4485 | 16S rRNA | Phenylalanine, tyrosine and tryptophan biosynthesis/cytosol | [60] |
| MiRNA | Function | Cancer Type | Mechanisms of Action/Cytotoxicity Response | Reference |
|---|---|---|---|---|
| miR-98 | OncomiR | Bladder cancer | ↑miR-98 ↑ resistance to cisplatin and doxorubicin in T24 bladder cancer cells | [50] |
| miR-27a | Suppressor | Breast cancer | ↑miR-27a ↑ROS ↑cytotoxicity to doxorubicin and paclitaxel in MCF-7 and MDA-MB-231 cells | [45] |
| miR-27a | OncomiR | Breast cancer | ↑ miR-27a ↑resistance to cisplatin in T-47D cells | [51] |
| miR-125b | Suppressor | Breast cancer | ↑miR-125b ↑ROS ↑ cytotoxicity to doxorubicin in MCF-7/R cells | [46] |
| miR-125b | Suppressor | Breast cancer | ↓miR-125b ↓cytotoxicity to doxorubicin in MCF-7 cells | [47] |
| miR-133a | Suppressor | Breast cancer | ↓miR-133a ↑UCP-2 ↑ doxorubicin-resistant to doxorubicin in MCF-7/DOX cells | [48] |
| miR-223 | Suppressor | Breast cancer | ↑ miR-223 ↑cytotoxicity to doxorubicin or cisplatin in MDA-MB-231 cells | [75] |
| miR-519d | Suppressor | Breast cancer | ↑ miR-519d ↑ cytotoxicity to cisplatin in T-47D-cancer stem cells. | [76] |
| miR-let-7a | Suppressor | Breast cancer Melanoma | ↑miR-let-7a ↑Cytotoxicity to doxorubicin in MDA-MB-231 cells | [44] |
| miR-593-5p | OncomiR | Tongue cancer | ↑miR-593-5p ↓ MFF ↑ resistance to cisplatin in Cal-27 and SCC-9 cells | [53] |
| miR-2392 | OncomiR | Tongue cancer | ↑miR-2392 ↑ glycolysis ↑ resistance to cisplatin in CAL-27 and SCC-9 cells | [38] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosolen, D.; Nunes-Souza, E.; Marchi, R.; Tofolo, M.V.; Antunes, V.C.; Berti, F.C.B.; Fonseca, A.S.; Cavalli, L.R. MiRNAs Action and Impact on Mitochondria Function, Metabolic Reprogramming and Chemoresistance of Cancer Cells: A Systematic Review. Biomedicines 2023, 11, 693. https://doi.org/10.3390/biomedicines11030693
Rosolen D, Nunes-Souza E, Marchi R, Tofolo MV, Antunes VC, Berti FCB, Fonseca AS, Cavalli LR. MiRNAs Action and Impact on Mitochondria Function, Metabolic Reprogramming and Chemoresistance of Cancer Cells: A Systematic Review. Biomedicines. 2023; 11(3):693. https://doi.org/10.3390/biomedicines11030693
Chicago/Turabian StyleRosolen, Daiane, Emanuelle Nunes-Souza, Rafael Marchi, Maria Vitoria Tofolo, Valquíria C. Antunes, Fernanda C. B. Berti, Aline S. Fonseca, and Luciane R. Cavalli. 2023. "MiRNAs Action and Impact on Mitochondria Function, Metabolic Reprogramming and Chemoresistance of Cancer Cells: A Systematic Review" Biomedicines 11, no. 3: 693. https://doi.org/10.3390/biomedicines11030693