
Sperm Mitochondrial Content and Mitochondrial DNA to Nuclear DNA Ratio Are Associated with Body Mass Index and Progressive Motility
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants’ Characteristics and Semen Sample Collection
2.2. Sperm DNA Extraction and Quantification
2.3. Real-Time Quantitive PCR–qPCR
- (a)
- By deducting the mean mitochondrial Ct value from the mean nuclear Ct value for each sample as indicated ΔCt = mtDNA − nDNA;
- (b)
- The average ΔCt value for the control group, which in this instance consisted of men with normal body mass index (18.5 to 24.9 kg/m2) and normal semen characteristics;
- (c)
- The ΔΔCt for each sample by subtracting the ΔCt of the control group from the sample’s mean ΔCt, that is, ΔΔCt = ΔCt of a particular sample minus the ΔCt of the control group [17];
- (d)
- By applying the formula 2−ΔΔCt, the fold difference.
2.4. Statistical Analysis
2.5. Inclusion and Exclusion Criteria
3. Results
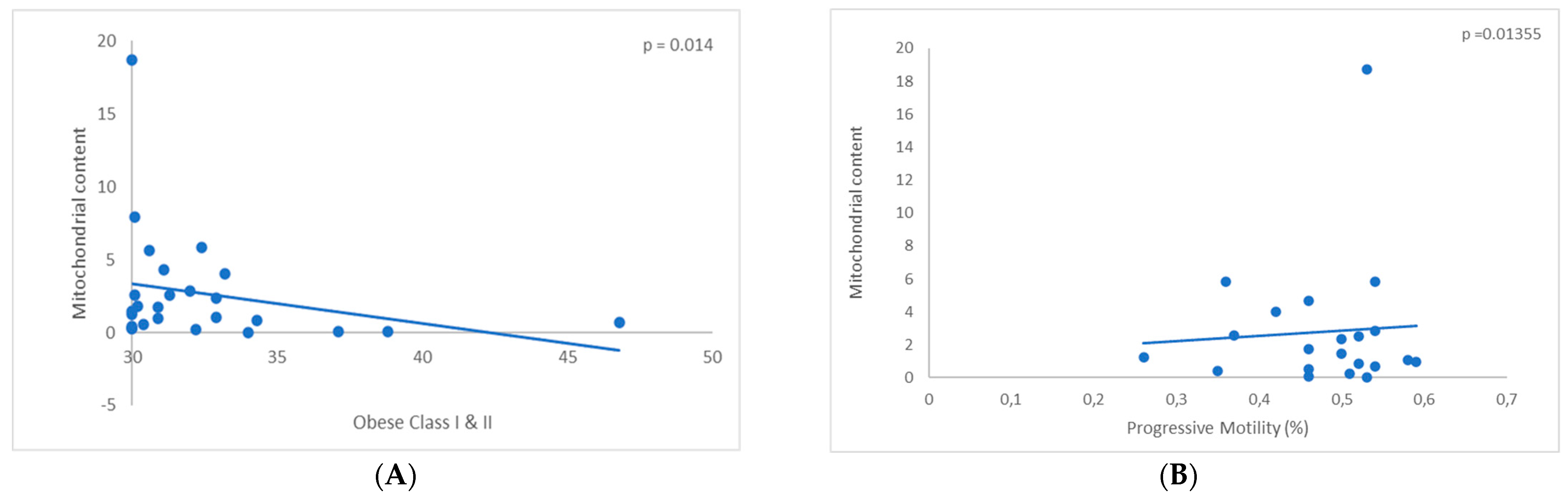
3.1. Assessment of Mitochondrial DNA Content
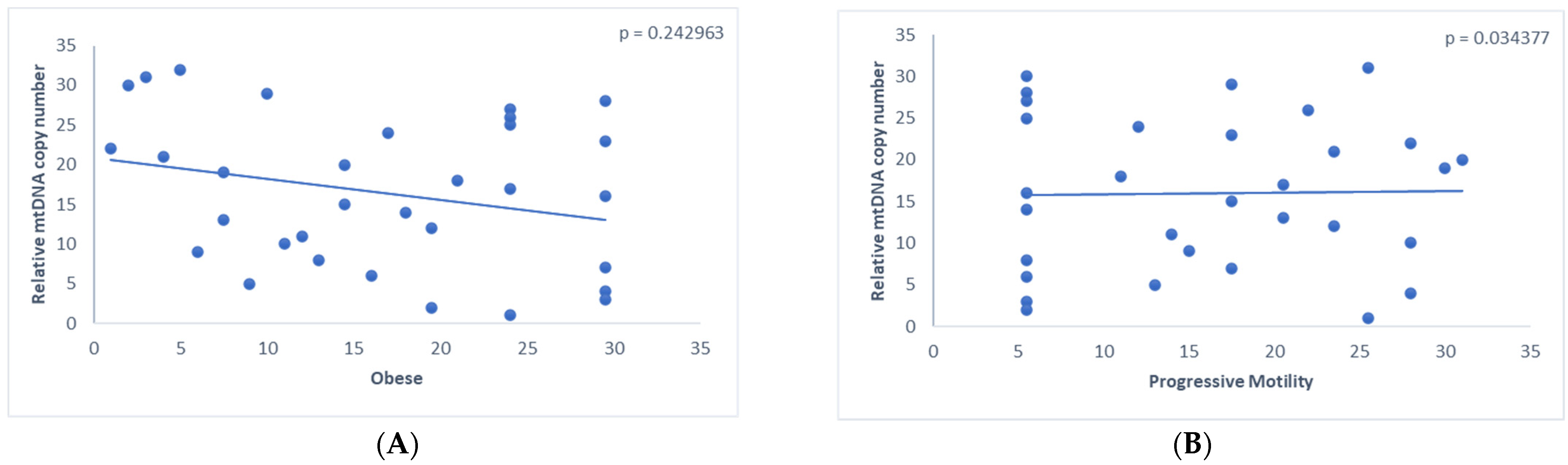
3.2. Assessment of the Relative mtDNA Copy Number
3.3. Assessment of the Ratio mtDNA to Nuclear DNA
4. Discussion
5. Conclusions
6. Limitations of the Study
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hirata, S.; Hoshi, K.; Shoda, T.; Mabuchi, T. Spermatozoon and mitochondrial DNA: Spermatozoon and mitochondrial DNA. Reprod. Med. Biol. 2002, 1, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.; Braga, P.C.; Rebelo, I.; Oliveira, P.F.; Alves, M.G. Mitochondria Quality Control and Male Fertility. Biology 2023, 12, 827. [Google Scholar] [CrossRef]
- Park, Y.-J.; Pang, M.-G. Mitochondrial Functionality in Male Fertility: From Spermatogenesis to Fertilization. Antioxidants 2021, 10, 98. [Google Scholar] [CrossRef]
- Wu, H.; Whitcomb, B.W.; Huffman, A.; Brandon, N.; Labrie, S.; Tougias, E.; Lynch, K.; Rahil, T.; Sites, C.K.; Pilsner, J.R. Associations of sperm mitochondrial DNA copy number and deletion rate with fertilization and embryo development in a clinical setting. Hum. Reprod. 2019, 34, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Rong, Z.; Tu, P.; Xu, P.; Sun, Y.; Yu, F.; Tu, N.; Guo, L.; Yang, Y. The Mitochondrial Response to DNA Damage. Front. Cell Dev. Biol. 2021, 9, 669379. [Google Scholar] [CrossRef] [PubMed]
- Zuidema, D.; Jones, A.; Song, W.-H.; Zigo, M.; Sutovsky, P. Identification of candidate mitochondrial inheritance determinants using the mammalian cell-free system. eLife 2023, 12, RP85596. [Google Scholar] [CrossRef] [PubMed]
- Amor, H.; Hammadeh, M.E. A Systematic Review of the Impact of Mitochondrial Variations on Male Infertility. Genes 2022, 13, 1182. [Google Scholar] [CrossRef]
- Wang, J.-J.; Wang, S.-X.; Tehmina; Feng, Y.; Zhang, R.-F.; Li, X.-Y.; Sun, Q.; Ding, J. Age-Related Decline of Male Fertility: Mitochondrial Dysfunction and the Antioxidant Interventions. Pharmaceuticals 2022, 15, 519. [Google Scholar] [CrossRef] [PubMed]
- Durairajanayagam, D.; Singh, D.; Agarwal, A.; Henkel, R. Causes and consequences of sperm mitochondrial dysfunction. Andrologia 2021, 53, e13666. [Google Scholar] [CrossRef]
- Gualtieri, R.; Kalthur, G.; Barbato, V.; Di Nardo, M.; Adiga, S.K.; Talevi, R. Mitochondrial Dysfunction and Oxidative Stress Caused by Cryopreservation in Reproductive Cells. Antioxidants 2021, 10, 337. [Google Scholar] [CrossRef]
- Chianese, R.; Pierantoni, R. Mitochondrial Reactive Oxygen Species (ROS) Production Alters Sperm Quality. Antioxidants 2021, 10, 92. [Google Scholar] [CrossRef]
- Ramaraju, G.A.; Teppala, S.; Prathigudupu, K.; Kalagara, M.; Thota, S.; Kota, M.; Cheemakurthi, R. Association between obesity and sperm quality. Andrologia 2018, 50, e12888. [Google Scholar] [CrossRef]
- Palmer, N.O.; Bakos, H.W.; Fullston, T.; Lane, M. Impact of obesity on male fertility, sperm function and molecular composition. Spermatogenesis 2012, 2, 253–263. [Google Scholar] [CrossRef]
- Ide, T.; Tsutsui, H.; Hayashidani, S.; Kang, D.; Suematsu, N.; Nakamura, K.; Utsumi, H.; Hamasaki, N.; Takeshita, A. Mitochondrial DNA Damage and Dysfunction Associated with Oxidative Stress in Failing Hearts After Myocardial Infarction. Circ. Res. 2001, 88, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Check, J.H.; Adelson, H.G.; Schubert, B.R.; Bollendorf, A. Evaluation of Sperm Morphology Using Kruger’s Strict Criteria. Arch. Androl. 1992, 28, 15–17. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Quiros, P.M.; Goyal, A.; Jha, P.; Auwerx, J. Analysis of mtDNA/nDNA Ratio in Mice. Curr. Protoc. Mouse Biol. 2017, 7, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Moustakli, E.; Zikopoulos, A.; Sakaloglou, P.; Bouba, I.; Sofikitis, N.; Georgiou, I. Functional association between telomeres, oxidation and mitochondria. Front. Reprod. Health 2023, 5, 1107215. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Peng, Y.; Fan, W.; Han, S.; Peng, Q.; Xue, C.; Qin, X.; Liu, Y.; Ding, Z. Obesity-induced oxidative stress and mitochondrial dysfunction negatively affect sperm quality. FEBS Open Bio 2023, 13, 763–778. [Google Scholar] [CrossRef] [PubMed]
- WHO. Obesity and Overweight. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 5 September 2023).
- Levine, H.; Jørgensen, N.; Martino-Andrade, A.; Mendiola, J.; Weksler-Derri, D.; Mindlis, I.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.; Kunstmann, J.M.; Czyglik, F.; Jouannet, P. Decline in semen quality among fertile men in Paris during the past 20 years. N. Engl. J. Med. 1995, 332, 281–285. [Google Scholar] [CrossRef]
- Skuratovskaia, D.; Litvinova, L.; Vulf, M.; Zatolokin, P.; Popadin, K.; Mazunin, I. From Normal to Obesity and Back: The Associations between Mitochondrial DNA Copy Number, Gender, and Body Mass Index. Cells 2019, 8, 430. [Google Scholar] [CrossRef]
- Ankel-Simons, F.; Cummins, J.M. Misconceptions about mitochondria and mammalian fertilization: Implications for theories on human evolution. Proc. Natl. Acad. Sci. USA 1996, 93, 13859–13863. [Google Scholar] [CrossRef]
- Reynier, P.; Chrétien, M.F.; Savagner, F.; Larcher, G.; Rohmer, V.; Barrière, P.; Malthièry, Y. Long PCR analysis of human gamete mtDNA suggests defective mitochondrial maintenance in spermatozoa and supports the bottleneck theory for oocytes. Biochem. Biophys. Res. Commun. 1998, 252, 373–377. [Google Scholar] [CrossRef]
- Cummins, J. Mitochondrial DNA in mammalian reproduction. Rev. Reprod. 1998, 3, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Song, G.J.; Lewis, V. Mitochondrial DNA integrity and copy number in sperm from infertile men. Fertil. Steril. 2008, 90, 2238–2244. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C.; Schon, E.A. Eliminating mitochondrial DNA from sperm. Dev. Cell 2012, 22, 469–470. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Huffman, A.M.; Whitcomb, B.W.; Josyula, S.; Labrie, S.; Tougias, E.; Rahil, T.; Sites, C.K.; Pilsner, J.R. Sperm mitochondrial DNA measures and semen parameters among men undergoing fertility treatment. Reprod. Biomed. Online 2019, 38, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Bao, H.; Martin, F.L.; Zhang, J.; Liu, L.; Huang, Q.; Shen, H. Association of DNA methylation and mitochondrial DNA copy number with human semen quality. Biol. Reprod. 2014, 91, 101. [Google Scholar] [CrossRef] [PubMed]
- May-Panloup, P.; Chrétien, M.-F.; Savagner, F.; Vasseur, C.; Jean, M.; Malthièry, Y.; Reynier, P. Increased sperm mitochondrial DNA content in male infertility. Hum. Reprod. Oxf. Engl. 2003, 18, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Díez-Sánchez, C.; Ruiz-Pesini, E.; Lapeña, A.C.; Montoya, J.; Pérez-Martos, A.; Enríquez, J.A.; López-Pérez, M.J. Mitochondrial DNA content of human spermatozoa. Biol. Reprod. 2003, 68, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Swerdloff, R.S. Limitations of semen analysis as a test of male fertility and anticipated needs from newer tests. Fertil. Steril. 2014, 102, 1502–1507. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Cardel, M.; Donahoo, W.T. Social and Environmental Factors Influencing Obesity. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. Available online: http://www.ncbi.nlm.nih.gov/books/NBK278977/ (accessed on 23 October 2023).
- The impact of body mass index on reproductive hormones, testosterone/estradiol ratio and semen parameters. Cent. Eur. J. Urol. 2020, 73, 226–230. [CrossRef]
- Bakos, H.W.; Henshaw, R.C.; Mitchell, M.; Lane, M. Paternal body mass index is associated with decreased blastocyst development and reduced live birth rates following assisted reproductive technology. Fertil. Steril. 2011, 95, 1700–1704. [Google Scholar] [CrossRef] [PubMed]
- Rahban, R.; Nef, S. Regional difference in semen quality of young men: A review on the implication of environmental and lifestyle factors during fetal life and adulthood. Basic Clin. Androl. 2020, 30, 16. [Google Scholar] [CrossRef] [PubMed]
- Irigoyen, P.; Pintos-Polasky, P.; Rosa-Villagran, L.; Skowronek, M.F.; Cassina, A.; Sapiro, R. Mitochondrial metabolism determines the functional status of human sperm and correlates with semen parameters. Front. Cell Dev. Biol. 2022, 10, 926684. [Google Scholar] [CrossRef]
- Todosenko, N.; Khaziakhmatova, O.; Malashchenko, V.; Yurova, K.; Bograya, M.; Beletskaya, M.; Vulf, M.; Gazatova, N.; Litvinova, L. Mitochondrial Dysfunction Associated with mtDNA in Metabolic Syndrome and Obesity. Int. J. Mol. Sci. 2023, 24, 12012. [Google Scholar] [CrossRef]
- Kozopas, N.M.; Chornenka, O.I.; Vorobets, M.Z.; Lapovets, L.Y.; Maksymyuk, H.V. Body Mass Index and Sperm Quality: Is there a Relationship? J. Hum. Reprod. Sci. 2020, 13, 110–113. [Google Scholar] [CrossRef]
- Hildebrandt, X.; Ibrahim, M.; Peltzer, N. Cell death and inflammation during obesity: “Know my methods, WAT(son)”. Cell Death Differ. 2023, 30, 279–292. [Google Scholar] [CrossRef]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef]
- Jung, U.; Choi, M.-S. Obesity and Its Metabolic Complications: The Role of Adipokines and the Relationship between Obesity, Inflammation, Insulin Resistance, Dyslipidemia and Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.-Y.; Huang, Y.; Du, Q.-Y.; Yao, G.-D.; Sun, Y.-P. Body mass index effects sperm quality: A retrospective study in Northern China. Asian J. Androl. 2017, 19, 234. [Google Scholar] [CrossRef] [PubMed]
- Darand, M.; Salimi, Z.; Ghorbani, M.; Sadeghi, N.; Babaie, S.; Hosseinzadeh, M. Obesity is associated with quality of sperm parameters in men with infertility: A cross-sectional study. Reprod. Health 2023, 20, 134. [Google Scholar] [CrossRef]
- Hussain, T.; Kandeel, M.; Metwally, E.; Murtaza, G.; Kalhoro, D.H.; Yin, Y.; Tan, B.; Chughtai, M.I.; Yaseen, A.; Afzal, A.; et al. Unraveling the harmful effect of oxidative stress on male fertility: A mechanistic insight. Front. Endocrinol. 2023, 14, 1070692. [Google Scholar] [CrossRef]
- Shamsi, M.; Kumar, R.; Bhatt, A.; Bamezai, R.N.K.; Kumar, R.; Gupta, N.; Das, T.; Dada, R. Mitochondrial DNA mutations in etiopathogenesis of male infertility. Indian J. Urol. 2008, 24, 150. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Pair | Sequence (Forward/Reverse) | Amplicon Base Pair (bp) | Tm |
|---|---|---|---|
| Nuclear region (gene RPE) | Forward 5′-ATAGGAAGCCAGAGAAGAGAGACT-3′ Reverse 5′-TCTATCTCTGCGGACTTTGAGCAT-3′ | 200 | 60 °C |
| Mitochondrial region | Forward 5′-TAGAGGAGCCTGTTCTGTAATCG-3′ Reverse 5′-TAAGGGCTATCGTAGTTTTCTGG-3′ | 205 | 59 °C |
| Inclusion Criteria | Exclusion Criteria |
|---|---|
| Males of reproductive age | Males without informed consent |
| IVF/ICSI participants | Urological conditions (e.g., varicocele) |
| Lifestyle (smoking, alcohol, drugs etc.) | Abnormal Karyotype |
| AZF deletions |
| Semen Characteristics | Normal | Overweight | Obese | p-Values |
|---|---|---|---|---|
| No of participants (total = 100) | 30 | 38 | 32 | |
| Male age, CI 95% | 32.5 ± 4.93 | 36.73 ± 5.614 | 30 ± 3.099 | 2.39 × 10−24 |
| Progressive motility (%), CI 95% | 43.93 ± 6.1 | 40.33 ± 5.0 | 43.00 ± 3.6 | 5.26 × 10−12 |
| Sperm count, million/mL, CI 95% | 36.68 ± 1.421 | 25.04 ± 1.1137 | 89 ± 2.2742 | 0.0068 |
| Sperm morphology (%), CI 95% | 13.38 ± 4.6 | 10.14 ± 6.8 | 10.00 ± 5.0 | 2.32 × 10−10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moustakli, E.; Zikopoulos, A.; Skentou, C.; Bouba, I.; Tsirka, G.; Stavros, S.; Vrachnis, D.; Vrachnis, N.; Potiris, A.; Georgiou, I.; et al. Sperm Mitochondrial Content and Mitochondrial DNA to Nuclear DNA Ratio Are Associated with Body Mass Index and Progressive Motility. Biomedicines 2023, 11, 3014. https://doi.org/10.3390/biomedicines11113014
Moustakli E, Zikopoulos A, Skentou C, Bouba I, Tsirka G, Stavros S, Vrachnis D, Vrachnis N, Potiris A, Georgiou I, et al. Sperm Mitochondrial Content and Mitochondrial DNA to Nuclear DNA Ratio Are Associated with Body Mass Index and Progressive Motility. Biomedicines. 2023; 11(11):3014. https://doi.org/10.3390/biomedicines11113014
Chicago/Turabian StyleMoustakli, Efthalia, Athanasios Zikopoulos, Charikleia Skentou, Ioanna Bouba, Georgia Tsirka, Sofoklis Stavros, Dionysios Vrachnis, Nikolaos Vrachnis, Anastasios Potiris, Ioannis Georgiou, and et al. 2023. "Sperm Mitochondrial Content and Mitochondrial DNA to Nuclear DNA Ratio Are Associated with Body Mass Index and Progressive Motility" Biomedicines 11, no. 11: 3014. https://doi.org/10.3390/biomedicines11113014