
Evaluation of Current Studies to Elucidate Processes in Dental Follicle Cells Driving Osteogenic Differentiation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Discovery of the BMP2/DLX3 Signaling Pathway
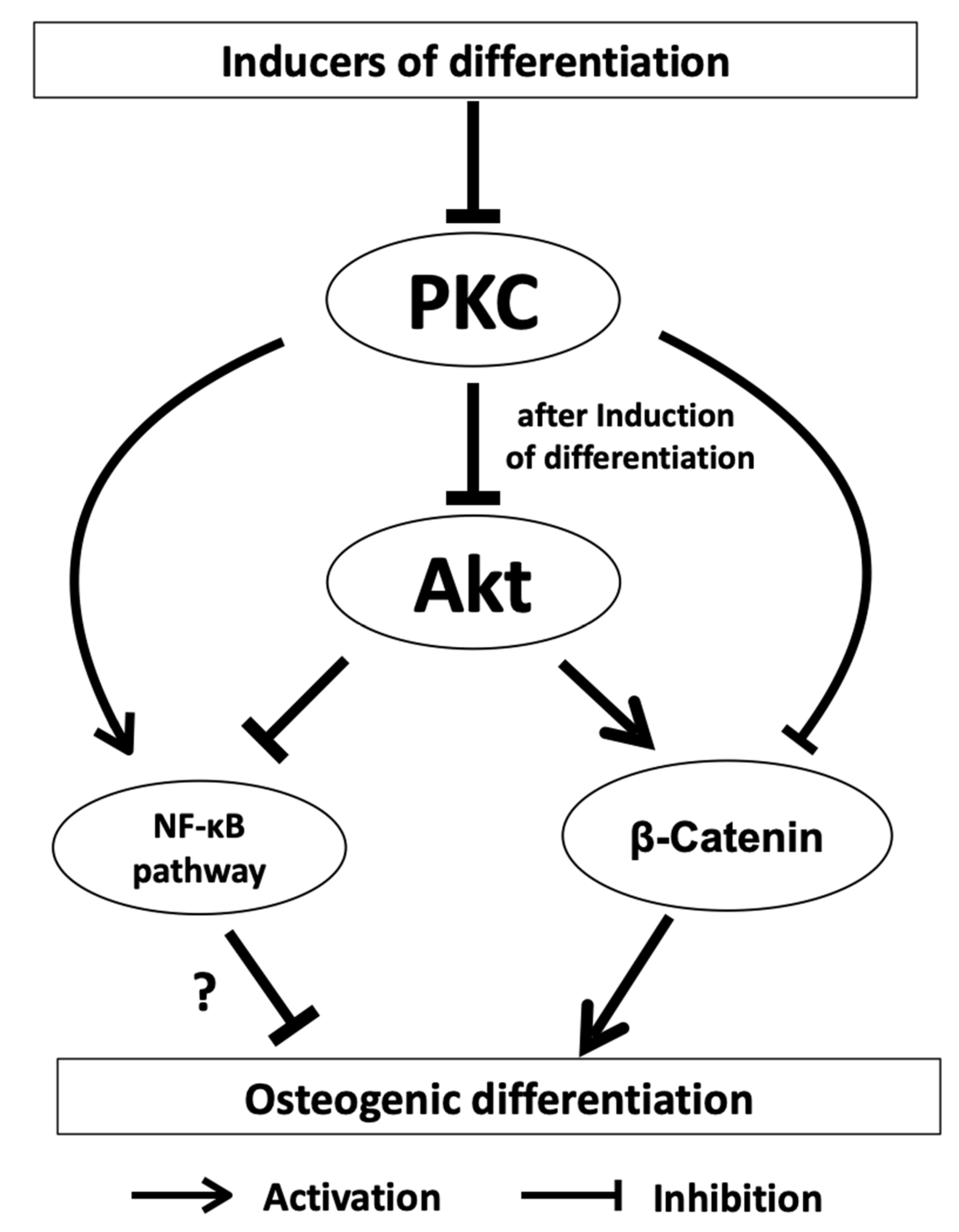
2. Protein Kinase C (PKC) Signaling and Biomineralization
2.1. PKC in Osteogenic Progenitor Cells
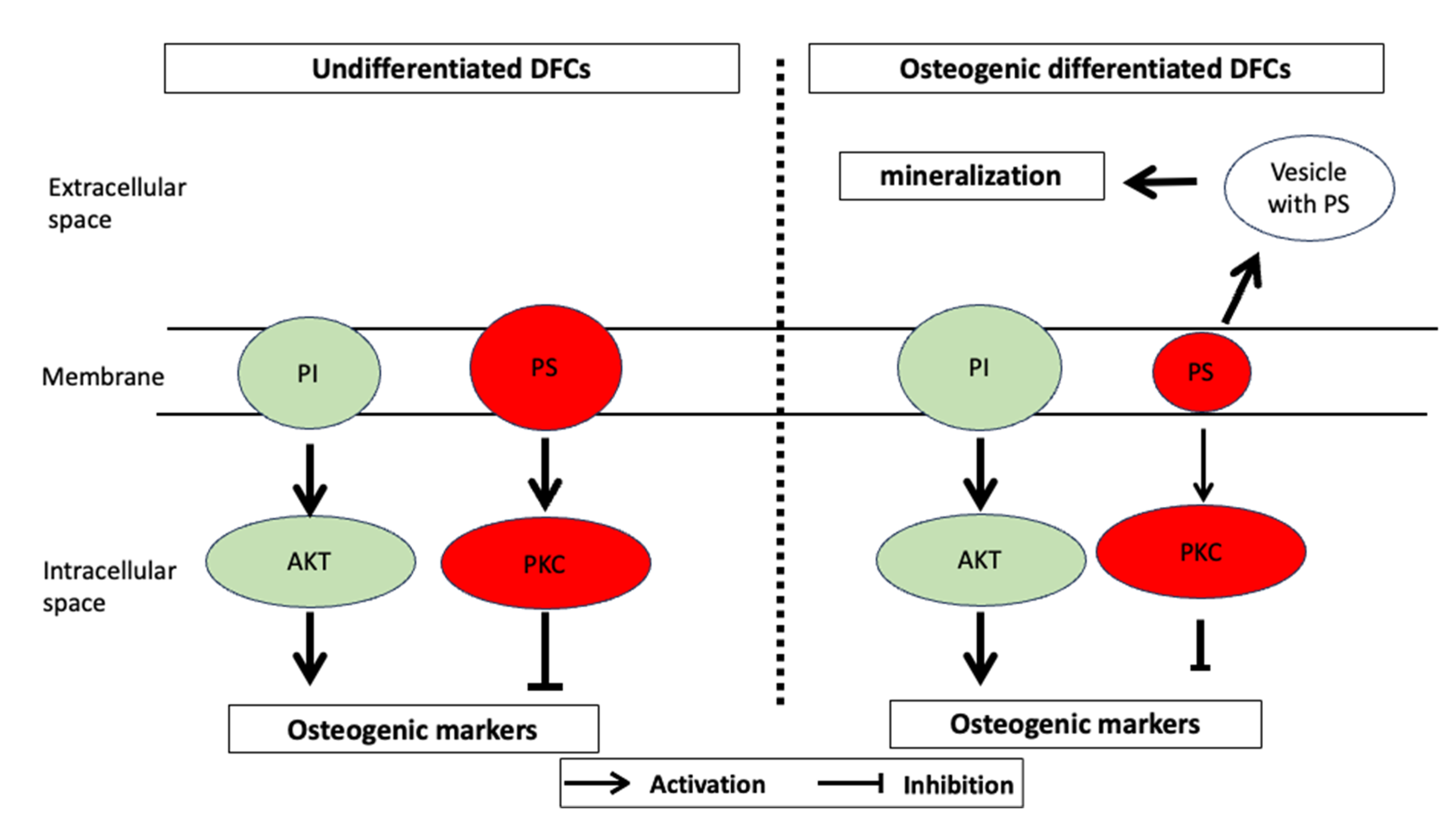
2.2. PKC Signaling in DFCs
3. Fatty Acid Synthesis and the Lipidome during Osteogenic Differentiation
3.1. Fatty Acid Synthesis
3.2. Lipidome
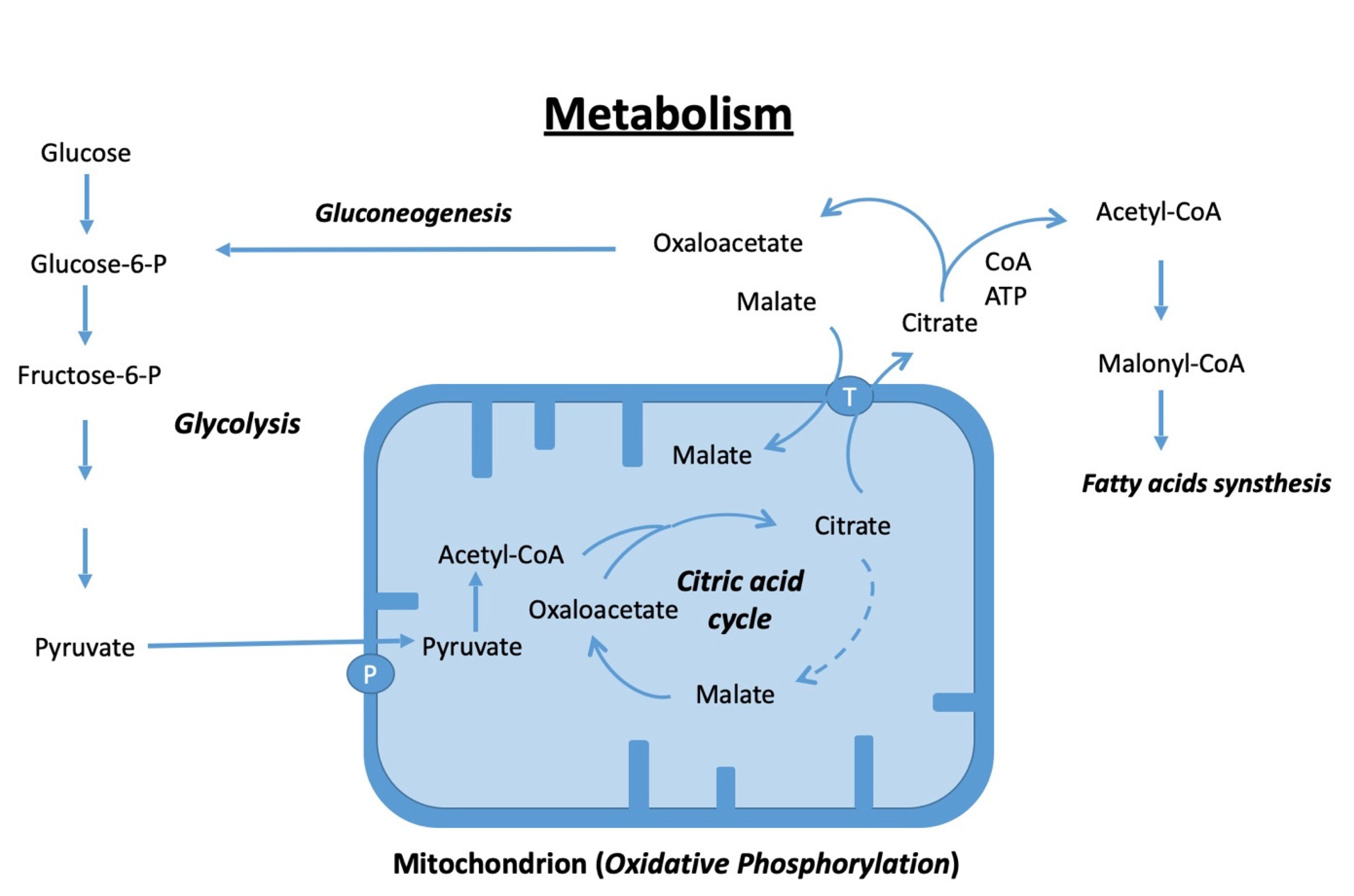
4. Energetic Metabolism during Osteogenic Differentiation
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fu, Y.; Miyazaki, K.; Chiba, Y.; Funada, K.; Yuta, T.; Tian, T.; Mizuta, K.; Kawahara, J.; Zhang, L.; Martin, D.; et al. Identification of GPI-anchored protein LYPD1 as an essential factor for odontoblast differentiation in tooth development. J. Biol. Chem. 2023, 299, 104638. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Fu, J.; Chen, Z.; Yang, G.; Yuan, G. Dlx3 Ubiquitination by Nuclear Mdm2 Is Essential for Dentinogenesis in Mice. J. Dent. Res. 2022, 101, 1064–1074. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yang, S.; Jiang, L.; Liu, J.; He, Y.; Sheng, X.; Chen, H.; Kang, J.; Jia, S.; Fan, W.; et al. Melatonin-mediated malic enzyme 2 orchestrates mitochondrial fusion and respiratory functions to promote odontoblastic differentiation during tooth development. J. Pineal Res. 2023, 74, e12865. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.S.; Liu, J.C.; Feng, Z.P.; Guo, S.L.; Wang, M.Z.; Wang, Z.; Li, Z.; Li, H.J.; Sui, L. N-acetylcysteine regulates dental follicle stem cell osteogenesis and alveolar bone repair via ROS scavenging. Stem Cell Res. Ther. 2022, 13, 466. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Zhao, W.; Liu, A.; Wu, M.; Shuai, Y.; Li, B.; Huang, X.; Liu, X.; Yang, X.; Guo, X.; et al. SHED promote angiogenesis in stem cell-mediated dental pulp regeneration. Biochem. Biophys. Res. Commun. 2020, 529, 1158–1164. [Google Scholar] [CrossRef]
- Morsczeck, C. Dental stem cells for tooth regeneration: How far have we come and where next? Expert. Opin. Biol. Ther. 2023, 23, 527–537. [Google Scholar] [CrossRef]
- Hermans, F.; Hemeryck, L.; Bueds, C.; Pereiro, M.T.; Hasevoets, S.; Kobayashi, H.; Lambrechts, D.; Lambrichts, I.; Bronckaers, A.; Vankelecom, H. Organoids from mouse molar and incisor as new tools to study tooth-specific biology and development. Stem Cell Rep. 2023, 18, 1166–1181. [Google Scholar] [CrossRef]
- Li, Z.; Yue, M.X.; Liu, Y.S.; Zhang, P.; Qing, J.; Liu, H.; Zhou, Y.S. Advances of Engineered Hydrogel Organoids within the Stem Cell Field: A Systematic Review. Gels 2022, 8, 379. [Google Scholar] [CrossRef]
- Morsczeck, C. Mechanisms during Osteogenic Differentiation in Human Dental Follicle Cells. Int. J. Mol. Sci. 2022, 23, 5945. [Google Scholar] [CrossRef]
- Guo, H.; Bai, X.Y.; Wang, X.L.; Qiang, J.B.; Sha, T.; Shi, Y.; Zheng, K.J.; Yang, Z.M.; Shi, C. Development and regeneration of periodontal supporting tissues. Genesis 2022, 60, e23491. [Google Scholar] [CrossRef]
- Nagata, M.; Ono, N.; Ono, W. Mesenchymal Progenitor Regulation of Tooth Eruption: A View from PTHrP. J. Dent. Res. 2020, 99, 10. [Google Scholar] [CrossRef]
- Morsczeck, C. Effects of Cellular Senescence on Dental Follicle Cells. Pharmacology 2021, 106, 137–142. [Google Scholar] [CrossRef]
- Morsczeck, C.; Moehl, C.; Gotz, W.; Heredia, A.; Schaffer, T.E.; Eckstein, N.; Sippel, C.; Hoffmann, K.H. In vitro differentiation of human dental follicle cells with dexamethasone and insulin. Cell Biol. Int. 2005, 29, 567–575. [Google Scholar] [CrossRef]
- Kemoun, P.; Narayanan, A.S.; Brunel, G.; Salles, J.P.; Laurencin-Dalicieux, S.; Rue, J.; Farges, J.C.; Gennero, I.; Conte-Auriol, F.; Briand-Mesange, F.; et al. Human dental follicle cells acquire cementoblast features under stimulation by BMP-2/-7 and enamel matrix derivatives (EMD) in vitro. Cell Tissue Res. 2007, 329, 283–294. [Google Scholar] [CrossRef]
- Honda, M.J.; Imaizumi, M.; Suzuki, H.; Ohshima, S.; Tsuchiya, S.; Satomura, K. Stem Cells Isolated from Human Dental Follicles Have Osteogenic Potential. Oral. Surg. Oral. Med. Oral. Pathol. Oral. Radiol. Endod. 2011, 111, 700–708. Available online: http://www.ncbi.nlm.nih.gov/pubmed/21147007 (accessed on 6 September 2023). [CrossRef] [PubMed]
- Popowics, T.; Foster, B.L.; Swanson, E.C.; Fong, H.; Somerman, M.J. Defining the Roots of Cementum Formation. Cells Tissues Organs 2005, 181, 248–257. Available online: http://www.ncbi.nlm.nih.gov/pubmed/16612090 (accessed on 6 September 2023). [CrossRef] [PubMed]
- Wise, G.E.; Frazier-Bowers, S.; D’Souza, R.N. Cellular, molecular, and genetic determinants of tooth eruption. Crit. Rev. Oral Biol. Med. An. Off. Publ. Am. Assoc. Oral. Biol. 2002, 13, 323–334. [Google Scholar] [CrossRef]
- Diekwisch, T.G. The Developmental Biology of Cementum. Int. J. Dev. Biol. 2001, 45, 695–706. Available online: http://www.ncbi.nlm.nih.gov/pubmed/11669371 (accessed on 6 September 2023). [PubMed]
- Morsczeck, C.; Gotz, W.; Schierholz, J.; Zeilhofer, F.; Kuhn, U.; Mohl, C.; Sippel, C.; Hoffmann, K.H. Isolation of precursor cells (PCs) from human dental follicle of wisdom teeth. Matrix Biol. 2005, 24, 155–165. [Google Scholar] [CrossRef]
- Morsczeck, C.; Frerich, B.; Driemel, O. Dental Stem Cell Patents. Recent. Pat. DNA Gene Seq. 2009, 3, 39–43. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19149737 (accessed on 6 September 2023). [CrossRef]
- Friedenstein, A.; Kuralesova, A.I. Osteogenic precursor cells of bone marrow in radiation chimeras. Transplantation 1971, 12, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef]
- Brunel, G.; Kémoun, P.; Laurencin-Dalicieux, S.; Rue, J.; Vaysse, F.; Roméas, A.; Arzate, H.; Conte-Auriol, F.; Farges, J.C.; Salles, J.P. Localization of STRO-1, BMP-2/-3/-7, BMP Receptors and Phosphorylated Smad-1 during the Formation of Mouse Periodontium. Tissue Cell 2007, 39, 257–266. Available online: http://www.ncbi.nlm.nih.gov/pubmed/17662325 (accessed on 6 September 2023).
- Völlner, F.; Ernst, W.; Driemel, O.; Morsczeck, C. A Two-Step Strategy for Neuronal Differentiation In Vitro of Human Dental Follicle Cells. Differ. Res. Biol. Divers. 2009, 77, 433–441. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19394129 (accessed on 6 September 2023). [CrossRef] [PubMed]
- Huang, Y.L.; Liu, L.; Liu, Q.; Huo, F.J.; Hu, X.Y.; Guo, S.J.; Tian, W.D. Dental follicle cells-derived small extracellular vesicles inhibit pathogenicity of Porphyromonas gingivalis. Oral. Diseases 2022, 29, 2297–2309. [Google Scholar] [CrossRef]
- Zhang, J.; Lan, T.; Han, X.; Xu, Y.; Liao, L.; Xie, L.; Yang, B.; Tian, W.; Guo, W. Improvement of ECM-based bioroot regeneration via N-acetylcysteine-induced antioxidative effects. Stem Cell Res. Ther. 2021, 12, 202. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yang, X.T.; Luo, X.Y.; Chen, G.; Chen, J.L.; Huo, F.J.; Zhu, Z.L.; Tian, Y.; Guo, W.H.; Tian, W.D. DFCs/TDM based artificial bio-root to obtain long-term functional root regeneration in non-human primate. Chem. Eng. J. 2023, 451, 138738. [Google Scholar] [CrossRef]
- Raju, R.; Oshima, M.; Inoue, M.; Morita, T.; Huijiao, Y.; Waskitho, A.; Baba, O.; Inoue, M.; Matsuka, Y. Three-dimensional periodontal tissue regeneration using a bone-ligament complex cell sheet. Sci. Rep. 2020, 10, 1656. [Google Scholar] [CrossRef]
- Tu, R.; Tang, X.A.; Xu, R.; Ping, Z.; Yu, Z.; Xie, T. Gap junction-transported cAMP from the niche controls stem cell progeny differentiation. Proc. Natl. Acad. Sci. USA 2023, 120, e2304168120. [Google Scholar] [CrossRef]
- Kopecny, L.R.; Lee, B.W.H.; Coroneo, M.T. A systematic review on the effects of ROCK inhibitors on proliferation and/or differentiation in human somatic stem cells: A hypothesis that ROCK inhibitors support corneal endothelial healing via acting on the limbal stem cell niche. Ocul. Surf. 2023, 27, 16–29. [Google Scholar] [CrossRef]
- Della Sala, F.; di Gennaro, M.; Lista, G.; Messina, F.; Valente, T.; Borzacchiello, A. Effect of Composition of Lung Biomimetic Niche on the Mesenchymal Stem Cell Differentiation toward Alveolar Type II Pneumocytes. Macromol. Biosci. 2023, 23, e2300035. [Google Scholar] [CrossRef]
- Lee, C.; Hong, S.N.; Kim, E.R.; Chang, D.K.; Kim, Y.H. Depletion of Intestinal Stem Cell Niche Factors Contributes to the Alteration of Epithelial Differentiation in SAMP1/YitFcsJ Mice With Crohn Disease-Like Ileitis. Inflamm. Bowel Dis. 2021, 27, 667–676. [Google Scholar] [CrossRef]
- Chua, J.S.; Muruganandam, G.; Sung, D.; Saijoh, Y.; Balagurunathan, K. Applications of Xylosides in the Manipulation of Stem Cell Niche to Regulate Human Neural Stem Cell Differentiation and Neurite Outgrowth. Methods Mol. Biol. 2022, 2303, 779–788. [Google Scholar] [CrossRef]
- Clemot, M.; Senos Demarco, R.; Jones, D.L. Lipid Mediated Regulation of Adult Stem Cell Behavior. Front. Cell Dev. Biol. 2020, 8, 115. [Google Scholar] [CrossRef]
- Saugspier, M.; Felthaus, O.; Viale-Bouroncle, S.; Driemel, O.; Reichert, T.E.; Schmalz, G.; Morsczeck, C. The Differentiation and Gene Expression Profile of Human Dental Follicle cells. Stem Cells Dev. 2010, 19, 707–717. Available online: http://www.ncbi.nlm.nih.gov/pubmed/20491563 (accessed on 6 September 2023). [CrossRef] [PubMed]
- Silvério, K.G.; Davidson, K.C.; James, R.G.; Adams, A.M.; Foster, B.L.; Nociti, F.H., Jr.; Somerman, M.J.; Moon, R.T. Wnt/β-Catenin Pathway Regulates Bone Morphogenetic Protein (BMP2)-Mediated Differentiation of Dental Follicle Cells. J. Periodontal Res. 2012, 47, 309–319. Available online: http://www.ncbi.nlm.nih.gov/pubmed/22150562 (accessed on 6 September 2023). [CrossRef] [PubMed]
- Viale-Bouroncle, S.; Gosau, M.; Morsczeck, C. Laminin regulates the osteogenic differentiation of dental follicle cells via integrin-α2/-β1 and the activation of the FAK/ERK signaling pathway. Cell Tissue Res. 2014, 357, 345–354. [Google Scholar] [CrossRef]
- Viale-Bouroncle, S.; Gosau, M.; Morsczeck, C. Collagen I induces the expression of alkaline phosphatase and osteopontin via independent activations of FAK and ERK signalling pathways. Arch. Oral. Biol. 2014, 59, 1249–1255. [Google Scholar] [CrossRef]
- Aonuma, H.; Ogura, N.; Takahashi, K.; Fujimoto, Y.; Iwai, S.; Hashimoto, H.; Ito, K.; Kamino, Y.; Kondoh, T. Characteristics and osteogenic differentiation of stem/progenitor cells in the human dental follicle analyzed by gene expression profiling. Cell Tissue Res. 2012, 350, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Morsczeck, C.; Petersen, J.; Völlner, F.; Driemel, O.; Reichert, T.; Beck, H.C. Proteomic Analysis of Osteogenic Differentiation of Dental Follicle Precursor Cells. Electrophoresis 2009, 30, 1175–1184. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19288589 (accessed on 6 September 2023). [CrossRef] [PubMed]
- Viale-Bouroncle, S.; Felthaus, O.; Schmalz, G.; Brockhoff, G.; Reichert, T.E.; Morsczeck, C. The Transcription Factor DLX3 Regulates the Osteogenic Differentiation of Human Dental Follicle Precursor Cells. Stem Cells Dev. 2012, 21, 1936–1947. Available online: http://www.ncbi.nlm.nih.gov/pubmed/22107079 (accessed on 6 September 2023). [CrossRef] [PubMed]
- Morsczeck, C.; Reichert, T.E. The dexamethasone induced osteogenic differentiation of dental follicle cells. Histol. Histopathol. 2017, 32, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Morsczeck, C.; Schmalz, G.; Reichert, T.E.; Völlner, F.; Saugspier, M.; Viale-Bouroncle, S.; Driemel, O. Gene Expression Profiles of Dental Follicle Cells before and after Osteogenic Differentiation In Vitro. Clin. Oral. Investig. 2009, 13, 383–391. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19252934 (accessed on 6 September 2023). [CrossRef] [PubMed]
- Choi, S.J.; Song, I.S.; Feng, J.Q.; Gao, T.; Haruyama, N.; Gautam, P.; Robey, P.G.; Hart, T.C. Mutant DLX 3 disrupts odontoblast polarization and dentin formation. Dev. Biol. 2010, 344, 682–692. [Google Scholar] [CrossRef]
- Hassan, M.Q.; Javed, A.; Morasso, M.I.; Karlin, J.; Montecino, M.; van Wijnen, A.J.; Stein, G.S.; Stein, J.L.; Lian, J.B. Dlx3 Transcriptional Regulation of Osteoblast Differentiation: Temporal Recruitment of Msx2, Dlx3, and Dlx5 Homeodomain Proteins to Chromatin of the Osteocalcin Gene. Mol. Cell Biol. 2004, 24, 9248–9261. Available online: http://www.ncbi.nlm.nih.gov/pubmed/15456894 (accessed on 6 September 2023). [CrossRef]
- Hassan, M.Q.; Tare, R.S.; Lee, S.H.; Mandeville, M.; Morasso, M.I.; Javed, A.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. BMP2 commitment to the osteogenic lineage involves activation of Runx2 by DLX3 and a homeodomain transcriptional network. J. Biol. Chem. 2006, 281, 40515–40526. [Google Scholar] [CrossRef]
- Viale-Bouroncle, S.; Klingelhöffer, C.; Ettl, T.; Reichert, T.E.; Morsczeck, C. A protein kinase A (PKA)/β-catenin pathway sustains the BMP2/DLX3-induced osteogenic differentiation in dental follicle cells (DFCs). Cell Signal. 2015, 27, 598–605. [Google Scholar] [CrossRef]
- Viale-Bouroncle, S.; Klingelhöffer, C.; Ettl, T.; Morsczeck, C. The AKT signaling pathway sustains the osteogenic differentiation in human dental follicle cells. Mol. Cell. Biochem. 2015, 406, 199–204. [Google Scholar] [CrossRef]
- Klingelhöffer, C.; Reck, A.; Ettl, T.; Morsczeck, C. The Parathyroid Hormone-Related Protein is Secreted during the Osteogenic Differentiation of Human Dental Follicle Cells and Inhibits the Alkaline Phosphatase Activity and the Expression of DLX3. Tissue Cell 2016, 48, 334–339. Available online: http://www.sciencedirect.com/science/article/pii/S0040816616300593 (accessed on 6 September 2023). [CrossRef]
- Pieles, O.; Reichert, T.E.; Morsczeck, C. Classical isoforms of protein kinase C (PKC) and Akt regulate the osteogenic differentiation of human dental follicle cells via both beta-catenin and NF-kappa B. Stem Cell Res. Ther. 2021, 12, 242. [Google Scholar] [CrossRef]
- Miraoui, H.; Oudina, K.; Petite, H.; Tanimoto, Y.; Moriyama, K.; Marie, P.J. Fibroblast growth factor receptor 2 promotes osteogenic differentiation in mesenchymal cells via ERK1/2 and protein kinase C signaling. J. Biol. Chem. 2009, 284, 4897–4904. [Google Scholar] [CrossRef] [PubMed]
- Nakura, A.; Higuchi, C.; Yoshida, K.; Yoshikawa, H. PKCalpha suppresses osteoblastic differentiation. Bone 2011, 48, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.M.; Jin, Y.H.; Choi, Y.H.; Yum, J.; Choi, J.K.; Yeo, C.Y.; Lee, K.Y. PKC signaling inhibits osteogenic differentiation through the regulation of Msx2 function. Bba-Mol. Cell Res. 2012, 1823, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Hu, C.; Li, J.W.; Liu, L.; Jing, W.; Tang, W.; Tian, W.D.; Long, J. Effect of miR-26a-5p on the Wnt/Ca2+ Pathway and Osteogenic Differentiation of Mouse Adipose-Derived Mesenchymal Stem Cells. Calcif. Tissue Int. 2016, 99, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Lotz, E.M.; Berger, M.B.; Boyan, B.D.; Schwartz, Z. Regulation of mesenchymal stem cell differentiation on microstructured titanium surfaces by semaphorin 3A. Bone 2020, 134, 115260. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, H.; He, J.; Lin, P.; Tong, Q.; Yang, J. Myeloma cells shift osteoblastogenesis to adipogenesis by inhibiting the ubiquitin ligase MURF1 in mesenchymal stem cells. Sci. Signal 2020, 13, eaay8203. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, X.; Zhang, L.; Wang, B.; Xu, B.; Zhang, J. GLP-1 inhibits PKCbeta2 phosphorylation to improve the osteogenic differentiation potential of hPDLSCs in the AGE microenvironment. J. Diabetes Complicat. 2020, 34, 107495. [Google Scholar] [CrossRef]
- Nemoto, E.; Sakisaka, Y.; Tsuchiya, M.; Tamura, M.; Nakamura, T.; Kanaya, S.; Shimonishi, M.; Shimauchi, H. Wnt3a signaling induces murine dental follicle cells to differentiate into cementoblastic/osteoblastic cells via an osterix-dependent pathway. J. Periodontal Res. 2015, 51, 164–174. [Google Scholar] [CrossRef]
- Nemoto, E.; Koshikawa, Y.; Kanaya, S.; Tsuchiya, M.; Tamura, M.; Somerman, M.J.; Shimauchi, H. Wnt Signaling Inhibits Cementoblast Differentiation and Promotes Proliferation. Bone 2009, 44, 805–812. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19442631 (accessed on 6 September 2023). [CrossRef]
- Sakisaka, Y.; Tsuchiya, M.; Nakamura, T.; Tamura, M.; Shimauchi, H.; Nemoto, E. Wnt5a attenuates Wnt3a-induced alkaline phosphatase expression in dental follicle cells. Exp. Cell Res. 2015, 336, 85–93. [Google Scholar] [CrossRef]
- Hozhabri, N.S.T.; Benson, M.D.; Vu, M.D.; Patel, R.H.; Martinez, R.M.; Nakhaie, F.N.; Kim, H.K.W.; Varanasi, V.G. Decreasing NF-kappa B Expression Enhances Odontoblastic Differentiation and Collagen Expression in Dental Pulp Stem Cells Exposed to Inflammatory Cytokines. PLoS ONE 2015, 10, e0127494. [Google Scholar] [CrossRef]
- Chang, J.; Liu, F.; Lee, M.; Wu, B.; Ting, K.; Zara, J.N.; Soo, C.; Al Hezaimi, K.; Zou, W.; Chen, X.; et al. NF-kappaB inhibits osteogenic differentiation of mesenchymal stem cells by promoting beta-catenin degradation. Proc. Natl. Acad. Sci. USA 2013, 110, 9469–9474. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, Z.; Wang, Y.; Chen, Y.; Ge, X.; Lu, J.; Bian, M.; Wu, J.; Yu, J. CTP-CM enhances osteogenic differentiation of hPDLSCs via NF-kappaB pathway. Oral. Dis. 2021, 27, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; He, W.; Weng, Y.; Wang, Y.; Liu, Y.; Xiang, Y.; Li, X.; Jiang, P.; Jin, Y.; Luo, J.; et al. Berberine inhibits osteogenic differentiation of aortic valve interstitial cells by interfering Smad1/5/8 and NF-kappaB pathways. Vascul Pharmacol. 2022, 144, 106986. [Google Scholar] [CrossRef] [PubMed]
- Holden, N.S.; Squires, P.E.; Kaur, M.; Bland, R.; Jones, C.E.; Newton, R. Phorbol ester-stimulated NF-kappaB-dependent transcription: Roles for isoforms of novel protein kinase C. Cell Signal 2008, 20, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Leonard, B.; McCann, J.L.; Starrett, G.J.; Kosyakovsky, L.; Luengas, E.M.; Molan, A.M.; Burns, M.B.; McDougle, R.M.; Parker, P.J.; Brown, W.L.; et al. The PKC/NF-kappa B Signaling Pathway Induces APOBEC3B Expression in Multiple Human Cancers. Cancer Res. 2015, 75, 4538–4547. [Google Scholar] [CrossRef]
- Niero, E.L.; Rocha-Sales, B.; Lauand, C.; Cortez, B.A.; de Souza, M.M.; Rezende-Teixeira, P.; Urabayashi, M.S.; Martens, A.A.; Neves, J.H.; Machado-Santelli, G.M. The multiple facets of drug resistance: One history, different approaches. J. Exp. Clin. Cancer Res. 2014, 33, 37. [Google Scholar] [CrossRef]
- Morsczeck, C.; Schmalz, G. Transcriptomes and Proteomes of DENTAL follicle Cells. J. Dent. Res. 2010, 89, 445–456. Available online: http://www.ncbi.nlm.nih.gov/pubmed/20348482 (accessed on 6 September 2023). [CrossRef]
- Kaul, N.; Forman, H.J. Activation of NF kappa B by the respiratory burst of macrophages. Free Radical Bio Med. 1996, 21, 401–405. [Google Scholar] [CrossRef]
- Busuttil, V.; Bottero, V.; Frelin, C.; Imbert, V.; Ricci, J.E.; Auberger, P.; Peyron, J.F. Blocking NF-kappaB activation in Jurkat leukemic T cells converts the survival agent and tumor promoter PMA into an apoptotic effector. Oncogene 2002, 21, 3213–3224. [Google Scholar] [CrossRef]
- Morsczeck, C.; Reck, A.; Reichert, T.E. WNT5A supports viability of senescent human dental follicle cells. Mol. Cell Biochem. 2018, 455, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Shirai, Y.; Saito, N. Activation mechanisms of protein kinase C: Maturation, catalytic activation, and targeting. J. Biochem. 2002, 132, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Nishizuka, Y. The Role of Protein Kinase-C in Cell-Surface Signal Transduction and Tumor Promotion. Nature 1984, 308, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Pieles, O.; Hoering, M.; Adel, S.; Reichert, T.E.; Liebisch, G.; Morsczeck, C. Energy Metabolism and Lipidome Are Highly Regulated during Osteogenic Differentiation of Dental Follicle Cells. Stem Cells Int. 2022, 2022, 3674931. [Google Scholar] [CrossRef]
- Chen, Y.; An, X.L.; Wang, Z.M.; Guan, S.H.; An, H.Y.; Huang, Q.Y.; Zhang, H.B.; Liang, L.; Huang, B.; Wang, H.Y.; et al. Transcriptome and lipidome profile of human mesenchymal stem cells with reduced senescence and increased trilineage differentiation ability upon drug treatment. Aging 2021, 13, 9991–10014. [Google Scholar] [CrossRef]
- Tigistu-Sahle, F.; Lampinen, M.; Kilpinen, L.; Holopainen, M.; Lehenkari, P.; Laitinen, S.; Kakela, R. Metabolism and phospholipid assembly of polyunsaturated fatty acids in human bone marrow mesenchymal stromal cells. J. Lipid Res. 2017, 58, 92–110. [Google Scholar] [CrossRef]
- Kilpinen, L.; Tigistu-Sahle, F.; Oja, S.; Greco, D.; Parmar, A.; Saavalainen, P.; Nikkila, J.; Korhonen, M.; Lehenkari, P.; Kakela, R.; et al. Aging bone marrow mesenchymal stromal cells have altered membrane glycerophospholipid composition and functionality. J. Lipid Res. 2013, 54, 622–635. [Google Scholar] [CrossRef]
- Kashirina, A.S.; Lopez-Duarte, I.; Kubankova, M.; Gulin, A.A.; Dudenkova, V.V.; Rodimova, S.A.; Torgomyan, H.G.; Zagaynova, E.V.; Meleshina, A.V.; Kuimova, M.K. Monitoring membrane viscosity in differentiating stem cells using BODIPY-based molecular rotors and FLIM. Sci. Rep. 2020, 10, 14063. [Google Scholar] [CrossRef]
- Kemp, E.T.; Zandberg, L.; Harvey, B.H.; Smuts, C.M.; Baumgartner, J. Iron and n-3 fatty acid depletion, alone and in combination, during early development provoke neurochemical changes, anhedonia, anxiety and social dysfunction in rats. Nutr. Neurosci. 2023. [Google Scholar] [CrossRef]
- Cinquina, V.; Keimpema, E.; Pollak, D.D.; Harkany, T. Adverse effects of gestational & omega;-3 and & omega;-6 polyunsaturated fatty acid imbalance on the programming of fetal brain development. J. Neuroendocrinol. 2023, 35, e13320. [Google Scholar] [CrossRef]
- Dubey, S.; Chen, C.W.; Patel, A.K.; Bhatia, S.K.; Singhania, R.R.; Dong, C.D. Development in health-promoting essential polyunsaturated fatty acids production by microalgae: A review. J. Food Sci. Technol. 2023. [Google Scholar] [CrossRef]
- Povero, D.; Chen, Y.B.; Johnson, S.M.; McMahon, C.E.; Pan, M.X.; Bao, H.M.; Petterson, X.M.T.; Blake, E.; Lauer, K.P.; O’Brien, D.R.; et al. HILPDA promotes NASH-driven HCC development by restraining intracellular fatty acid flux in hypoxia. J. Hepatol. 2023, 79, 378–393. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Chen, Y.; Wang, H.Y.; Bai, Y.F.; Zhao, J.X.; Zhang, X.H.; Liang, L.; Chen, Y.; Ye, C.F.; Li, Y.Q.; et al. Integrated Lipidomics and Transcriptomics Characterization upon Aging-Related Changes of Lipid Species and Pathways in Human Bone Marrow Mesenchymal Stem Cells. J. Proteome Res. 2019, 18, 2065–2077. [Google Scholar] [CrossRef]
- Levental, K.R.; Surma, M.A.; Skinkle, A.D.; Lorent, J.H.; Zhou, Y.; Klose, C.; Chang, J.T.; Hancock, J.F.; Levental, I. ω-3 polyunsaturated fatty acids direct differentiation of the membrane phenotype in mesenchymal stem cells to potentiate osteogenesis. Sci. Adv. 2017, 3, eaao1193. [Google Scholar] [CrossRef]
- Song, Z.; Wang, S.; Lu, L.; Xu, J.; Zhou, Q.; Lu, W.; Tong, H.; Zhang, Y.; Liu, W.; Wang, Z.; et al. Lipidomics and Transcriptomics Differ Liposarcoma Differentiation Characteristics That Can Be Altered by Pentose Phosphate Pathway Intervention. Metabolites 2022, 12, 1227. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhang, H.; Hu, Z.; Chin, Y.; Li, G.; Huang, J.; Zhang, X.; Jiang, B.; Hu, Y. Differentiation of three commercial tuna species through Q-Exactive Orbitrap mass spectrometry based lipidomics and chemometrics. Food Res. Int. 2022, 158, 111509. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhao, M.; Zhou, L.; Zhang, M.; Liu, J.; Marchioni, E. Identification and Differentiation of Wide Edible Mushrooms Based on Lipidomics Profiling Combined with Principal Component Analysis. J. Agric. Food Chem. 2021, 69, 9991–10001. [Google Scholar] [CrossRef]
- Silva, C.G.D.; Barretto, L.S.S.; Lo Turco, E.G.; Santos, A.L.; Lessio, C.; Martins Junior, H.A.; Almeida, F.G. Lipidomics of mesenchymal stem cell differentiation. Chem. Phys. Lipids 2020, 232, 104964. [Google Scholar] [CrossRef]
- Yu, X.; Chen, K.; Li, S.; Wang, Y.; Shen, Q. Lipidomics differentiation of soft-shelled turtle strains using hydrophilic interaction liquid chromatography and mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1112, 11–15. [Google Scholar] [CrossRef]
- Pieles, O.; Hartmann, M.; Morsczeck, C. AMP-activated protein kinase and the down-stream activated process of autophagy regulate the osteogenic differentiation of human dental follicle cells. Arch. Oral. Biol. 2021, 122, 104951. [Google Scholar] [CrossRef]
- Nebauer, R.; Rosenberger, S.; Daum, G. Phosphatidylethanolamine, a limiting factor of autophagy in yeast strains bearing a defect in the carboxypeptidase Y pathway of vacuolar targeting. J. Biol. Chem. 2007, 282, 16736–16743. [Google Scholar] [CrossRef]
- Newton, A.C.; Koshland, D.E., Jr. Phosphatidylserine affects specificity of protein kinase C substrate phosphorylation and autophosphorylation. Biochemistry 1990, 29, 6656–6661. [Google Scholar] [CrossRef]
- Xu, C.X.; Zhen, Z.B.; Fang, L.M.; Zha, N.R.; Lin, Z.H.; Liang, T.Z.; Zhai, Z.C.; Zhou, J.A. Phosphatidylserine enhances osteogenic differentiation in human mesenchymal stem cells via ERK signal pathways. Mat. Sci. Eng. C-Mater. 2013, 33, 1783–1788. [Google Scholar] [CrossRef] [PubMed]
- Muller, W.E.G.; Schroder, H.C.; Wang, X.H. The Understanding of the Metazoan Skeletal System, Based on the Initial Discoveries with Siliceous and Calcareous Sponges. Mar. Drugs 2017, 15, 172. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.A.E.; Ferreira, C.R.; Tovani, C.B.; de Oliveira, F.A.; Bolean, M.; Caseli, L.; Mebarek, S.; Millan, J.L.; Buchet, R.; Bottini, M.; et al. Phosphatidylserine controls calcium phosphate nucleation and growth on lipid monolayers: A physicochemical understanding of matrix vesicle-driven biomineralization. J. Struct. Biol. 2020, 212, 107607. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Li, L.; Weidong, T. The Role of Sphingolipid Metabolism in Bone Remodeling. Front. Cell Dev. Biol. 2021, 9, 752540. [Google Scholar] [CrossRef] [PubMed]
- Sato, C.; Iwasaki, T.; Kitano, S.; Tsunemi, S.; Sano, H. Sphingosine 1-phosphate receptor activation enhances BMP-2-induced osteoblast differentiation. Biochem. Biophys. Res. Commun. 2012, 423, 200–205. [Google Scholar] [CrossRef]
- Li, K.; Xiu, C.; Zhou, Q.; Ni, L.; Du, J.; Gong, T.; Li, M.; Saijilafu; Yang, H.; Chen, J. A dual role of cholesterol in osteogenic differentiation of bone marrow stromal cells. J. Cell Physiol. 2019, 234, 2058–2066. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.J.; Liu, Y.H. Simvastatin induces the osteogenic differentiation of human periodontal ligament stem cells. Fund. Clin. Pharmacol. 2014, 28, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Tai, I.C.; Wang, Y.H.; Chen, C.H.; Chuang, S.C.; Chang, J.K.; Ho, M.L. Simvastatin enhances Rho/actin/cell rigidity pathway contributing to mesenchymal stem cells’ osteogenic differentiation. Int. J. Nanomed. 2015, 10, 5881–5894. [Google Scholar] [CrossRef]
- Viale-Bouroncle, S.; Völlner, F.; Möhl, C.; Küpper, K.; Brockhoff, G.; Reichert, T.E.; Schmalz, G.; Morsczeck, C. Soft Matrix Supports Osteogenic Differentiation of Human Dental Follicle Cells. Biochem. Biophys. Res. Commun. 2011, 410, 587–592. Available online: http://www.ncbi.nlm.nih.gov/pubmed/21684253 (accessed on 6 September 2023). [CrossRef] [PubMed]
- Sanchez-Hidalgo, M.; Lu, Z.; Tan, D.A.; Maldonado, M.D.; Reiter, R.J.; Gregerman, R.I. Melatonin inhibits fatty acid-induced triglyceride accumulation in ROS17/2.8 cells: Implications for osteoblast differentiation and osteoporosis. Am. J. Physiol-Reg. I 2007, 292, R2208–R2215. [Google Scholar] [CrossRef]
- Sun, H.Y.; Zheng, M.M.; Liu, J.W.; Fan, W.G.; He, H.W.; Huang, F. Melatonin promoted osteogenesis of human periodontal ligament cells by regulating mitochondrial functions through the translocase of the outer mitochondrial membrane 20. J. Periodontal Res. 2023, 58, 53–69. [Google Scholar] [CrossRef]
- Shum, L.C.; White, N.S.; Mills, B.N.; Bentley, K.L.; Eliseev, R.A. Energy Metabolism in Mesenchymal Stem Cells During Osteogenic Differentiation. Stem Cells Dev. 2016, 25, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Gao, Z.; Chen, Y.; Guan, M.X. The role of mitochondria in osteogenic, adipogenic and chondrogenic differentiation of mesenchymal stem cells. Protein Cell 2017, 8, 439–445. [Google Scholar] [CrossRef]
- Smith, C.O.; Eliseev, R.A. Energy Metabolism During Osteogenic Differentiation: The Role of Akt. Stem Cells Dev. 2021, 30, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Maheshwari, D.; Nautiyal, N.; Shubham, S.; Rooge, S.; Anand, L.; Vyas, A.; Kumari, R.; Sharma, S.; Bihari, C.; et al. Defects in energy metabolism are associated with functional exhaustion of bone marrow mesenchymal stem cells in cirrhosis. Am. J. Stem Cells 2022, 11, 12–27. [Google Scholar]
- Mao, H.; Yang, A.; Zhao, Y.; Lei, L.; Li, H. Succinate Supplement Elicited “Pseudohypoxia” Condition to Promote Proliferation, Migration, and Osteogenesis of Periodontal Ligament Cells. Stem Cells Int. 2020, 2020, 2016809. [Google Scholar] [CrossRef]
- Deng, L.D.; Yi, S.Q.; Yin, X.H.; Li, Y.; Luan, Q.X. MFN2 knockdown promotes osteogenic differentiation of iPSC-MSCs through aerobic glycolysis mediated by the Wnt/beta-catenin signaling pathway. Stem Cell Res. Ther. 2022, 13, 162. [Google Scholar] [CrossRef]
- Guo, Y.; Chi, X.; Wang, Y.; Heng, B.C.; Wei, Y.; Zhang, X.; Zhao, H.; Yin, Y.; Deng, X. Mitochondria transfer enhances proliferation, migration, and osteogenic differentiation of bone marrow mesenchymal stem cell and promotes bone defect healing. Stem Cell Res. Ther. 2020, 11, 245. [Google Scholar] [CrossRef]
- Zhang, Y.L.; An, Y.; Sun, L.J.; Qu, H.L.; Li, X.; He, X.T.; Wu, R.X.; Chen, F.M.; Tian, B.M.; Yin, Y. NADPH-dependent ROS accumulation contributes to the impaired osteogenic differentiation of periodontal ligament stem cells under high glucose conditions. Front. Endocrinol 2023, 14, 1152845. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, C.; Khoury, M. Musculoskeletal Progenitor/Stromal Cell-Derived Mitochondria Modulate Cell Differentiation and Therapeutical Function. Front. Immunol. 2021, 12, 606781. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Picone, G.; Cappadone, C.; Sorrentino, A.; Columbaro, M.; Farruggia, G.; Catelli, E.; Sciutto, G.; Prati, S.; Oliete, R.; et al. Shedding Light on Osteosarcoma Cell Differentiation: Impact on Biomineralization and Mitochondria Morphology. Int. J. Mol. Sci. 2023, 24, 8559. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Mechanistic Insight into Oxidative Stress-Triggered Signaling Pathways and Type 2 Diabetes. Molecules 2022, 27, 950. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.R. Reactive Oxygen Species as the Molecular Modulators of Calcium Oxalate Kidney Stone Formation: Evidence from Clinical and Experimental Investigations. J. Urol. 2013, 189, 803–811. [Google Scholar] [CrossRef]
- Sarviya, N.; Basu, S.M.; Mani, R.; Chauhan, M.; Kingshott, P.; Giri, J. Biomimicking nanofibrous gelatin microspheres recreating the stem cell niche for their ex-vivo expansion and in-vivo like differentiation for injectable stem cell transplantation. Biomater. Adv. 2022, 139, 212981. [Google Scholar] [CrossRef]
- Yi, G.; Zhang, S.; Ma, Y.; Yang, X.; Huo, F.; Chen, Y.; Yang, B.; Tian, W. Matrix vesicles from dental follicle cells improve alveolar bone regeneration via activation of the PLC/PKC/MAPK pathway. Stem Cell Res. Ther. 2022, 13, 41. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morsczeck, C.; De Pellegrin, M.; Reck, A.; Reichert, T.E. Evaluation of Current Studies to Elucidate Processes in Dental Follicle Cells Driving Osteogenic Differentiation. Biomedicines 2023, 11, 2787. https://doi.org/10.3390/biomedicines11102787
Morsczeck C, De Pellegrin M, Reck A, Reichert TE. Evaluation of Current Studies to Elucidate Processes in Dental Follicle Cells Driving Osteogenic Differentiation. Biomedicines. 2023; 11(10):2787. https://doi.org/10.3390/biomedicines11102787
Chicago/Turabian StyleMorsczeck, Christian, Michela De Pellegrin, Anja Reck, and Torsten E. Reichert. 2023. "Evaluation of Current Studies to Elucidate Processes in Dental Follicle Cells Driving Osteogenic Differentiation" Biomedicines 11, no. 10: 2787. https://doi.org/10.3390/biomedicines11102787