
Cytokine-Induced iNOS in A549 Alveolar Epithelial Cells: A Potential Role in COVID-19 Lung Pathology
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Models
2.2. Experimental Treatments
2.3. RT-qPCR Analysis
2.4. Western Blot Analysis
2.5. Cytokine Analysis
2.6. Cell Viability
2.7. Determination of Nitric Oxide Production
2.8. Statistical Analysis
2.9. Materials
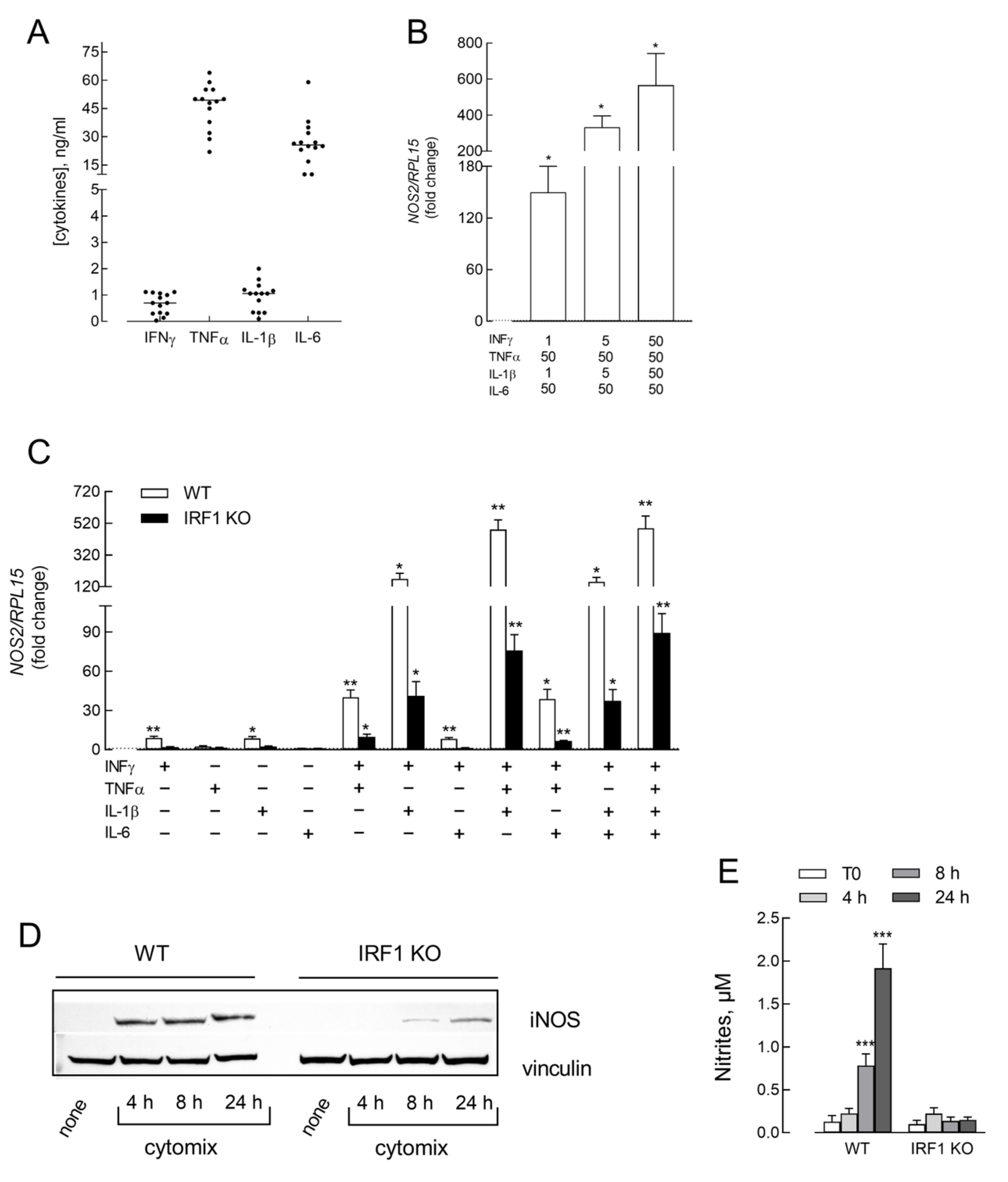
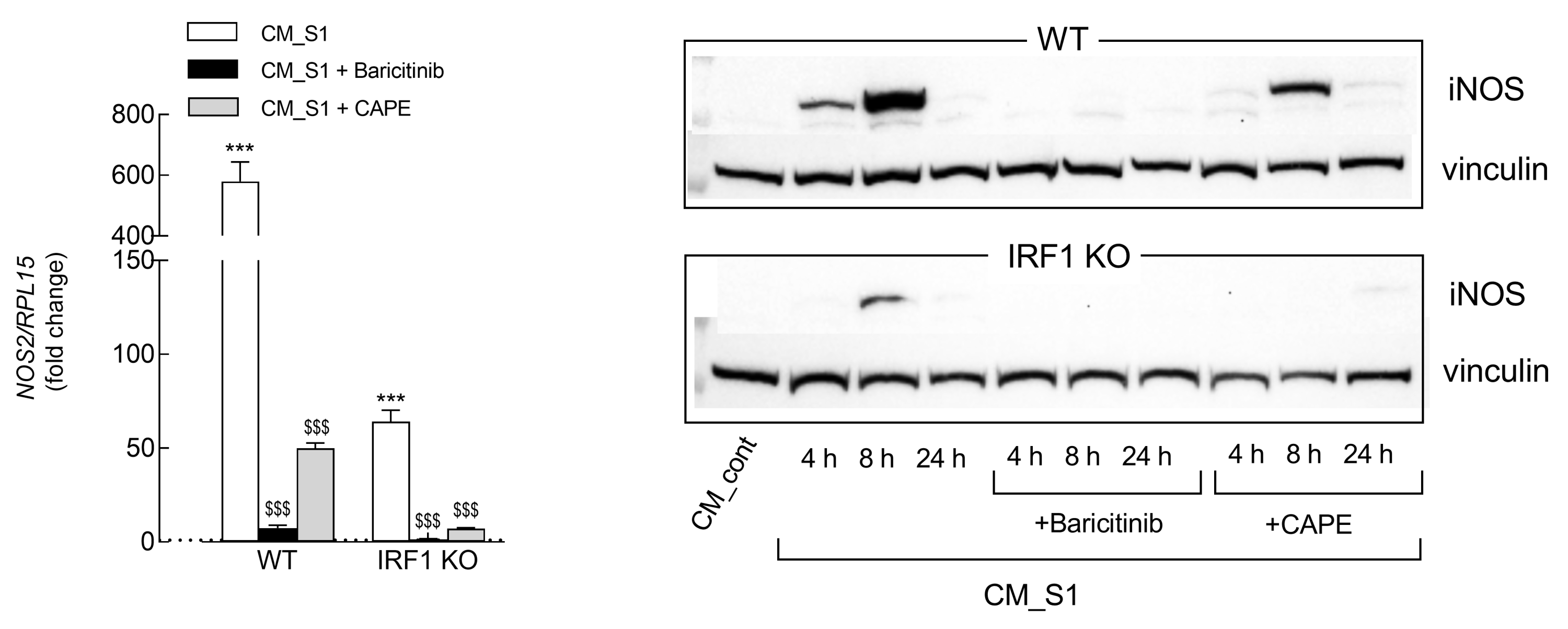
3. Results
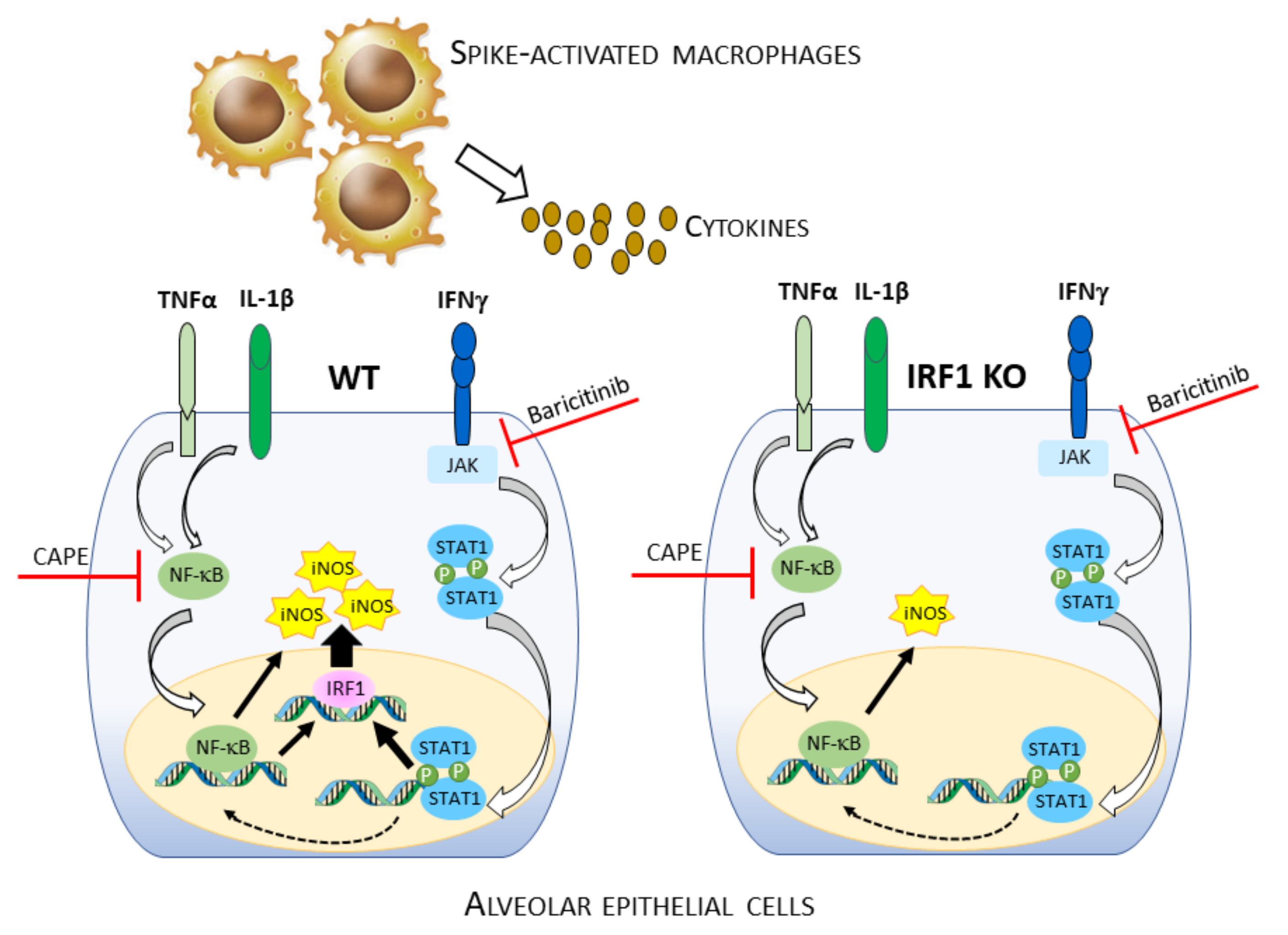
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Cron, R.Q. COVID-19 cytokine storm: Targeting the appropriate cytokine. Lancet Rheumatol. 2021, 3, e236–e237. [Google Scholar] [CrossRef] [PubMed]
- Aslan, A.; Aslan, C.; Zolbanin, N.M.; Jafari, R. Acute respiratory distress syndrome in COVID-19: Possible mechanisms and therapeutic management. Pneumonia 2021, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Alomair, B.M.; Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Al-Buhadily, A.K.; Alexiou, A.; Papadakis, M.; Alshammari, M.A.; Saad, H.M.; El-Saber Batiha, G. Mixed storm in SARS-CoV-2 infection: A narrative review and new term in the COVID-19 era. Immun. Inflamm. Dis. 2023, 11, e838. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.J.A.; Ribeiro, L.R.; Gouveia, M.I.M.; Marcelino, B.D.R.; Santos, C.S.D.; Lima, K.V.B.; Lima, L.N.G.C. Hyperinflammatory response in COVID-19: A systematic review. Viruses 2023, 15, 553. [Google Scholar] [CrossRef]
- Matthay, M.A.; Zemans, R.L.; Zimmerman, G.A.; Arabi, Y.M.; Beitler, J.R.; Mercat, A.; Herridge, M.; Randolph, A.G.; Calfee, C.S. Acute respiratory distress syndrome. Nat. Rev. Dis. Primers 2019, 5, 18. [Google Scholar] [CrossRef]
- Chakraborty, R.K.; Burns, B. Systemic inflammatory response syndrome. In Statpearls; Statpearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Robertson, C.M.; Coopersmith, C.M. The systemic inflammatory response syndrome. Microbes Infect. 2006, 8, 1382–1389. [Google Scholar] [CrossRef]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef]
- Chi, Y.; Ge, Y.; Wu, B.; Zhang, W.; Wu, T.; Wen, T.; Liu, J.; Guo, X.; Huang, C.; Jiao, Y.; et al. Serum cytokine and chemokine profile in relation to the severity of coronavirus disease 2019 in china. J. Infect. Dis. 2020, 222, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Chen, Z.; Lui, G.; Wong, C.K.; Wong, W.T.; Ng, R.W.Y.; Tso, E.Y.K.; Fung, K.S.C.; Chan, V.; Yeung, A.C.M.; et al. Longitudinal cytokine profile in patients with mild to critical COVID-19. Front. Immunol. 2021, 12, 763292. [Google Scholar] [CrossRef]
- Dharra, R.; Kumar Sharma, A.; Datta, S. Emerging aspects of cytokine storm in COVID-19: The role of proinflammatory cytokines and therapeutic prospects. Cytokine 2023, 169, 156287. [Google Scholar] [CrossRef]
- Faraj, S.S.; Jalal, P.J. Il1beta, il-6, and tnf-alpha cytokines cooperate to modulate a complicated medical condition among covid-19 patients: Case-control study. Ann. Med. Surg. 2023, 85, 2291–2297. [Google Scholar] [CrossRef]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zhang, M.; Sundaram, B.; Banoth, B.; Malireddi, R.K.S.; et al. Synergism of tnf-alpha and ifn-gamma triggers inflammatory cell death, tissue damage, and mortality in SARS-CoV-2 infection and cytokine shock syndromes. Cell 2021, 184, 149–168.e17. [Google Scholar] [CrossRef]
- Beckman, J.S.; Koppenol, W.H. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol. 1996, 271, C1424–C1437. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide synthase in innate and adaptive immunity: An update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Predonzani, A.; Cali, B.; Agnellini, A.H.; Molon, B. Spotlights on immunological effects of reactive nitrogen species: When inflammation says nitric oxide. World J. Exp. Med. 2015, 5, 64–76. [Google Scholar] [CrossRef]
- Xue, Q.; Yan, Y.; Zhang, R.; Xiong, H. Regulation of inos on immune cells and its role in diseases. Int. J. Mol. Sci. 2018, 19, 3805. [Google Scholar] [CrossRef] [PubMed]
- Domenico, R. Pharmacology of nitric oxide: Molecular mechanisms and therapeutic strategies. Curr. Pharm. Des. 2004, 10, 1667–1676. [Google Scholar] [CrossRef]
- Zhao, K.; Huang, Z.; Lu, H.; Zhou, J.; Wei, T. Induction of inducible nitric oxide synthase increases the production of reactive oxygen species in raw264.7 macrophages. Biosci. Rep. 2010, 30, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Oswald, I.P.; Wynn, T.A.; Sher, A.; James, S.L. No as an effector molecule of parasite killing: Modulation of its synthesis by cytokines. Comp. Biochem. Physiol. Pharmacol. Toxicol. Endocrinol. 1994, 108, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Jorens, P.G.; Matthys, K.E.; Bult, H. Modulation of nitric oxide synthase activity in macrophages. Mediat. Inflamm. 1995, 4, 75–89. [Google Scholar] [CrossRef]
- Lee, J.Y.; Lowell, C.A.; Lemay, D.G.; Youn, H.S.; Rhee, S.H.; Sohn, K.H.; Jang, B.; Ye, J.; Chung, J.H.; Hwang, D.H. The regulation of the expression of inducible nitric oxide synthase by src-family tyrosine kinases mediated through myd88-independent signaling pathways of toll-like receptor 4. Biochem. Pharmacol. 2005, 70, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Salim, T.; Sershen, C.L.; May, E.E. Investigating the role of tnf-alpha and ifn-gamma activation on the dynamics of inos gene expression in lps stimulated macrophages. PLoS ONE 2016, 11, e0153289. [Google Scholar] [CrossRef] [PubMed]
- Barilli, A.; Visigalli, R.; Ferrari, F.; Recchia Luciani, G.; Soli, M.; Dall’Asta, V.; Rotoli, B.M. Growth arrest of alveolar cells in response to cytokines from spike s1-activated macrophages: Role of ifn-gamma. Biomedicines 2022, 10, 3085. [Google Scholar] [CrossRef] [PubMed]
- Barilli, A.; Visigalli, R.; Ferrari, F.; Bianchi, M.G.; Dall’Asta, V.; Rotoli, B.M. Immune-mediated inflammatory responses of alveolar epithelial cells: Implications for COVID-19 lung pathology. Biomedicines 2022, 10, 618. [Google Scholar] [CrossRef] [PubMed]
- Rotoli, B.M.; Barilli, A.; Visigalli, R.; Ferrari, F.; Dall’Asta, V. Endothelial cell activation by SARS-CoV-2 spike s1 protein: A crosstalk between endothelium and innate immune cells. Biomedicines 2021, 9, 1220. [Google Scholar] [CrossRef]
- Ingoglia, F.; Visigalli, R.; Rotoli, B.M.; Barilli, A.; Riccardi, B.; Puccini, P.; Dall’Asta, V. Functional activity of l-carnitine transporters in human airway epithelial cells. Biochim. Biophys. Acta 2016, 1858, 210–219. [Google Scholar] [CrossRef]
- Rotoli, B.M.; Barilli, A.; Visigalli, R.; Ingoglia, F.; Milioli, M.; Di Lascia, M.; Riccardi, B.; Puccini, P.; Dall’Asta, V. Downregulation of slc7a7 triggers an inflammatory phenotype in human macrophages and airway epithelial cells. Front. Immunol. 2018, 9, 508. [Google Scholar] [CrossRef]
- Barilli, A.; Visigalli, R.; Ferrari, F.; Borsani, G.; Dall’Asta, V.; Rotoli, B.M. Flagellin from pseudomonas aeruginosa stimulates atb(0,+) transporter for arginine and neutral amino acids in human airway epithelial cells. Front. Immunol. 2021, 12, 641563. [Google Scholar] [CrossRef]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the alamar blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef]
- Rotoli, B.M.; Visigalli, R.; Ferrari, F.; Ranieri, M.; Tamma, G.; Dall’Asta, V.; Barilli, A. Desmopressin stimulates nitric oxide production in human lung microvascular endothelial cells. Biomolecules 2022, 12, 389. [Google Scholar] [CrossRef]
- Barilli, A.; Visigalli, R.; Ferrari, F.; Recchia Luciani, G.; Soli, M.; Dall’Asta, V.; Rotoli, B.M. The jak1/2 inhibitor baricitinib mitigates the spike-induced inflammatory response of immune and endothelial cells in vitro. Biomedicines 2022, 10, 2324. [Google Scholar] [CrossRef]
- Feng, H.; Zhang, Y.B.; Gui, J.F.; Lemon, S.M.; Yamane, D. Interferon regulatory factor 1 (irf1) and anti-pathogen innate immune responses. PLoS Pathog. 2021, 17, e1009220. [Google Scholar] [CrossRef]
- Jorgensen, S.C.J.; Tse, C.L.Y.; Burry, L.; Dresser, L.D. Baricitinib: A review of pharmacology, safety, and emerging clinical experience in COVID-19. Pharmacotherapy 2020, 40, 843–856. [Google Scholar] [CrossRef]
- Natarajan, K.; Singh, S.; Burke, T.R.; Jr Grunberger, D.; Aggarwal, B.B. Caffeic acid phenethyl ester is a potent and specific inhibitor of activation of nuclear transcription factor nf-kappa b. Proc. Natl. Acad. Sci. USA 1996, 93, 9090–9095. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, L.M.F.; Rossini, C.V.T.; Lameu, C. Implications of SARS-CoV-2 infection on enos and inos activity: Consequences for the respiratory and vascular systems. Nitric Oxide 2021, 111–112, 64–71. [Google Scholar] [CrossRef]
- Gelzo, M.; Scialo, F.; Cacciapuoti, S.; Pinchera, B.; De Rosa, A.; Cernera, G.; Comegna, M.; Tripodi, L.; Moriello, N.S.; Mormile, M.; et al. Inducible nitric oxide synthase (inos): Why a different production in COVID-19 patients of the two waves? Viruses 2022, 14, 534. [Google Scholar] [CrossRef] [PubMed]
- Soufli, I.; Toumi, R.; Rafa, H.; Touil-Boukoffa, C. Overview of cytokines and nitric oxide involvement in immuno-pathogenesis of inflammatory bowel diseases. World J. Gastrointest. Pharmacol. Ther. 2016, 7, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Giannakodimos, I.; Gkountana, G.V.; Lykouras, D.; Karkoulias, K.; Tsakas, S. The role of interleukin-6 in the pathogenesis, prognosis and treatment of severe COVID-19. Curr. Med. Chem. 2021, 28, 5328–5338. [Google Scholar] [CrossRef]
- Asano, K.; Chee, C.B.; Gaston, B.; Lilly, C.M.; Gerard, C.; Drazen, J.M.; Stamler, J.S. Constitutive and inducible nitric oxide synthase gene expression, regulation, and activity in human lung epithelial cells. Proc. Natl. Acad. Sci. USA 1994, 91, 10089–10093. [Google Scholar] [CrossRef]
- Berkman, N.; Robichaud, A.; Robbins, R.A.; Roesems, G.; Haddad, E.B.; Barnes, P.J.; Chung, K.F. Inhibition of inducible nitric oxide synthase expression by interleukin-4 and interleukin-13 in human lung epithelial cells. Immunology 1996, 89, 363–367. [Google Scholar] [CrossRef]
- Robbins, R.A.; Barnes, P.J.; Springall, D.R.; Warren, J.B.; Kwon, O.J.; Buttery, L.D.; Wilson, A.; Geller, D.; Polak, J. Expression of inducible nitric oxide in human lung epithelial cells. Biochem. Biophys. Res. Commun. 1994, 203, 209–218. [Google Scholar] [CrossRef]
- Gross, T.J.; Kremens, K.; Powers, L.S.; Brink, B.; Knutson, T.; Domann, F.E.; Philibert, R.A.; Milhem, M.M.; Monick, M.M. Epigenetic silencing of the human NOS2 gene: Rethinking the role of nitric oxide in human macrophage inflammatory responses. J. Immunol. 2014, 192, 2326–2338. [Google Scholar] [CrossRef]
- Young, R.; Bush, S.J.; Lefevre, L.; McCulloch, M.E.B.; Lisowski, Z.M.; Muriuki, C.; Waddell, L.A.; Sauter, K.A.; Pridans, C.; Clark, E.L.; et al. Species-specific transcriptional regulation of genes involved in nitric oxide production and arginine metabolism in macrophages. Immunohorizons 2018, 2, 27–37. [Google Scholar] [CrossRef]
- Knowles, R.G.; Moncada, S. Nitric oxide synthases in mammals. Biochem. J. 1994, 298, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Albina, J.E. On the expression of nitric oxide synthase by human macrophages. Why no no? J. Leukoc. Biol. 1995, 58, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Schneemann, M.; Schoedon, G.; Hofer, S.; Blau, N.; Guerrero, L.; Schaffner, A. Nitric oxide synthase is not a constituent of the antimicrobial armature of human mononuclear phagocytes. J. Infect. Dis. 1993, 167, 1358–1363. [Google Scholar] [CrossRef] [PubMed]
- De Stefano, D.; Maiuri, M.C.; Iovine, B.; Ialenti, A.; Bevilacqua, M.A.; Carnuccio, R. The role of nf-kappab, irf-1, and stat-1alpha transcription factors in the inos gene induction by gliadin and ifn-gamma in raw 264.7 macrophages. J. Mol. Med. 2006, 84, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Yang, P.; Zhao, Y.; Zhuang, Z.; Wang, Z.; Song, R.; Zhang, J.; Liu, C.; Gao, Q.; Xu, Q.; et al. Single-cell sequencing of peripheral mononuclear cells reveals distinct immune response landscapes of COVID-19 and influenza patients. Immunity 2020, 53, 685–696.e3. [Google Scholar] [CrossRef]
- Miki, H.; Ohmori, Y. Transcriptional synergism between nf-κb and stat1. J. Oral Biosci. 2005, 47, 230–242. [Google Scholar]
- Ohmori, Y.; Schreiber, R.D.; Hamilton, T.A. Synergy between interferon-gamma and tumor necrosis factor-alpha in transcriptional activation is mediated by cooperation between signal transducer and activator of transcription 1 and nuclear factor kappab. J. Biol. Chem. 1997, 272, 14899–14907. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Ugel, S.; Tinazzi, E.; Vella, A.; De Sanctis, F.; Cane, S.; Batani, V.; Trovato, R.; Fiore, A.; Petrova, V.; et al. Baricitinib restrains the immune dysregulation in patients with severe COVID-19. J. Clin. Investig. 2020, 130, 6409–6416. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, V.; Finn, A.; Lal, A.; Khan, M.S.; Dapaah-Afriyie, K.; Carino, G.P. Baricitinib in hospitalised patients with COVID-19: A meta-analysis of randomised controlled trials. EClinicalMedicine 2022, 49, 101489. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Protein | Forward Primer | Reverse Primer |
|---|---|---|
| RPL15/RPL15 | GCAGCCATCAGGTAAGCCAAG | AGCGGACCCTCAGAAGAAAGC |
| IRF1/IRF1 | CTGTGCGAGTGTACCGGATG | ATCCCCACATGACTTCCTCTT |
| NOS2/iNOS | CACGCTCGCCTTCAAGTTC | AGGCACTAATGTAGGACCCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barilli, A.; Recchia Luciani, G.; Visigalli, R.; Sala, R.; Soli, M.; Dall’Asta, V.; Rotoli, B.M. Cytokine-Induced iNOS in A549 Alveolar Epithelial Cells: A Potential Role in COVID-19 Lung Pathology. Biomedicines 2023, 11, 2699. https://doi.org/10.3390/biomedicines11102699
Barilli A, Recchia Luciani G, Visigalli R, Sala R, Soli M, Dall’Asta V, Rotoli BM. Cytokine-Induced iNOS in A549 Alveolar Epithelial Cells: A Potential Role in COVID-19 Lung Pathology. Biomedicines. 2023; 11(10):2699. https://doi.org/10.3390/biomedicines11102699
Chicago/Turabian StyleBarilli, Amelia, Giulia Recchia Luciani, Rossana Visigalli, Roberto Sala, Maurizio Soli, Valeria Dall’Asta, and Bianca Maria Rotoli. 2023. "Cytokine-Induced iNOS in A549 Alveolar Epithelial Cells: A Potential Role in COVID-19 Lung Pathology" Biomedicines 11, no. 10: 2699. https://doi.org/10.3390/biomedicines11102699