
FDG PET Imaging of the Pain Matrix in Neuropathic Pain Model Rats
 ,
,
Abstract
:1. Introduction
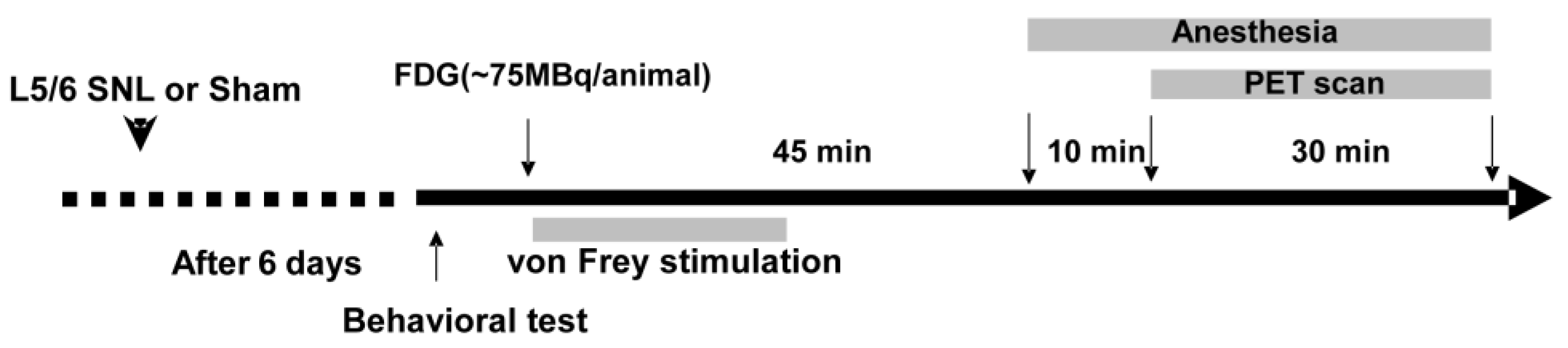
2. Materials and Methods
2.1. Neuropathic Pain Animal Model Preparation
2.2. Behavior Test
2.3. PET Scanning
2.4. Image Analysis
2.5. Data Analyses
3. Results
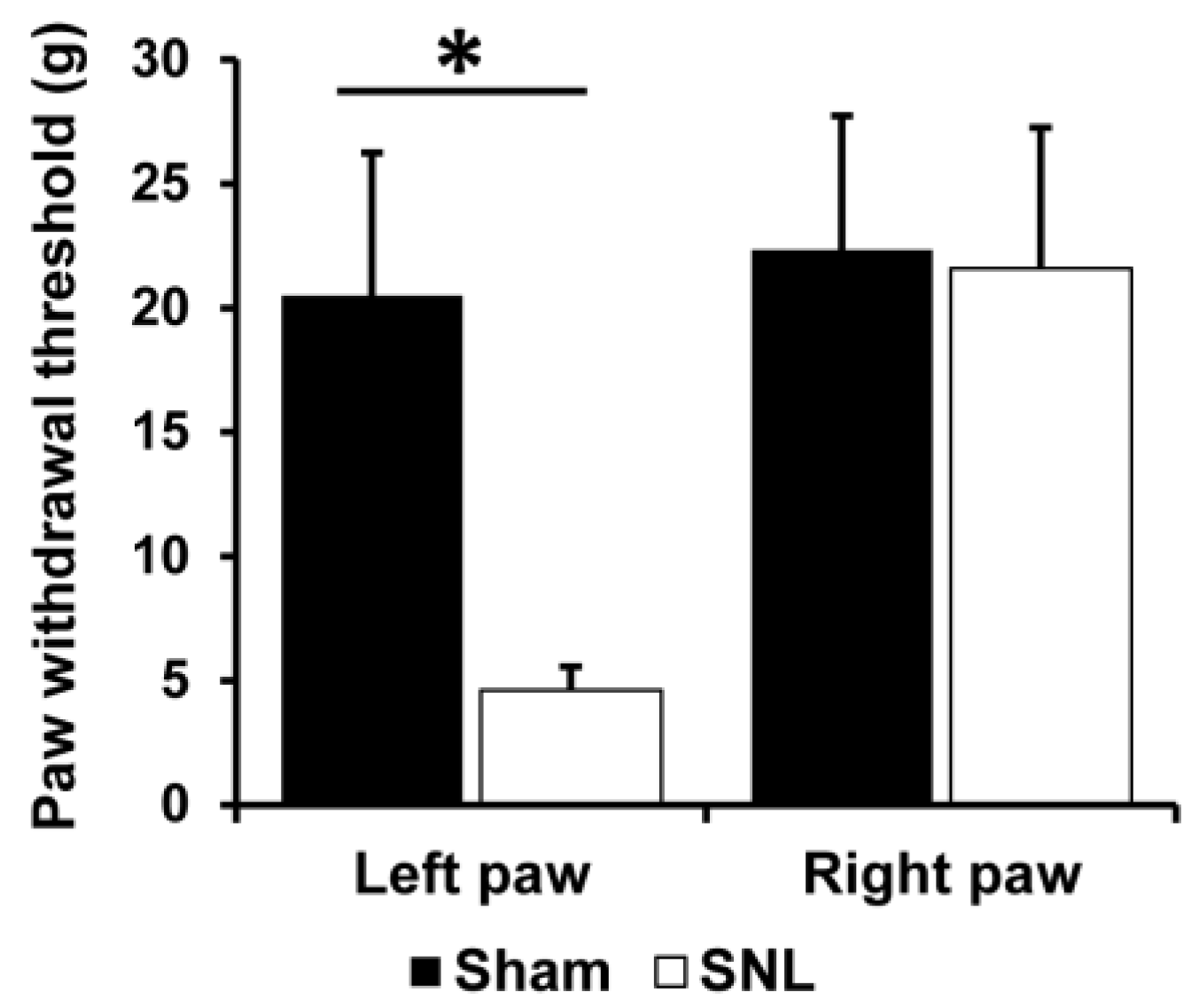
3.1. Mechanical Allodynia in SNL Rats
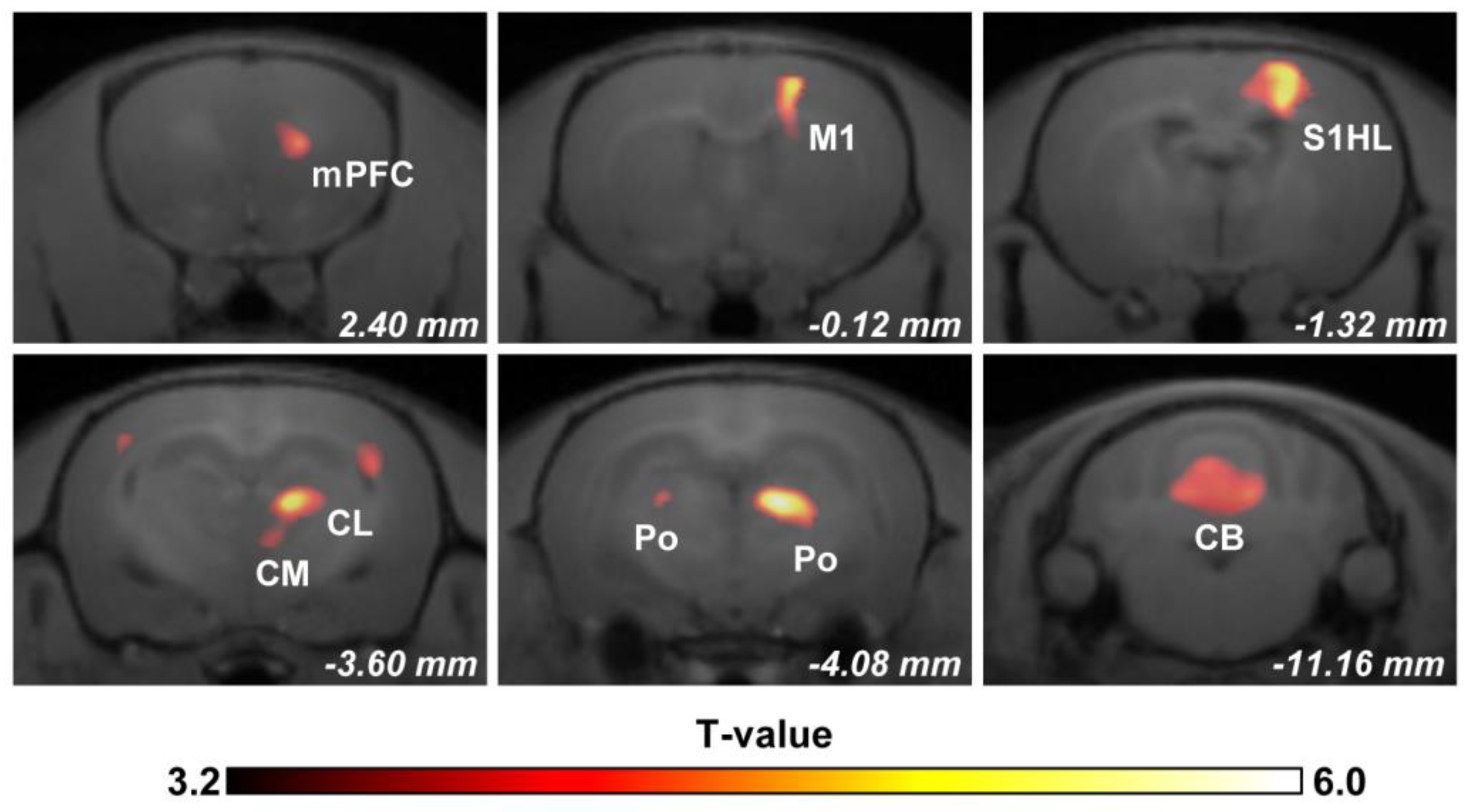
3.2. Regional Brain Activity in Response to Mechanical Stimulation
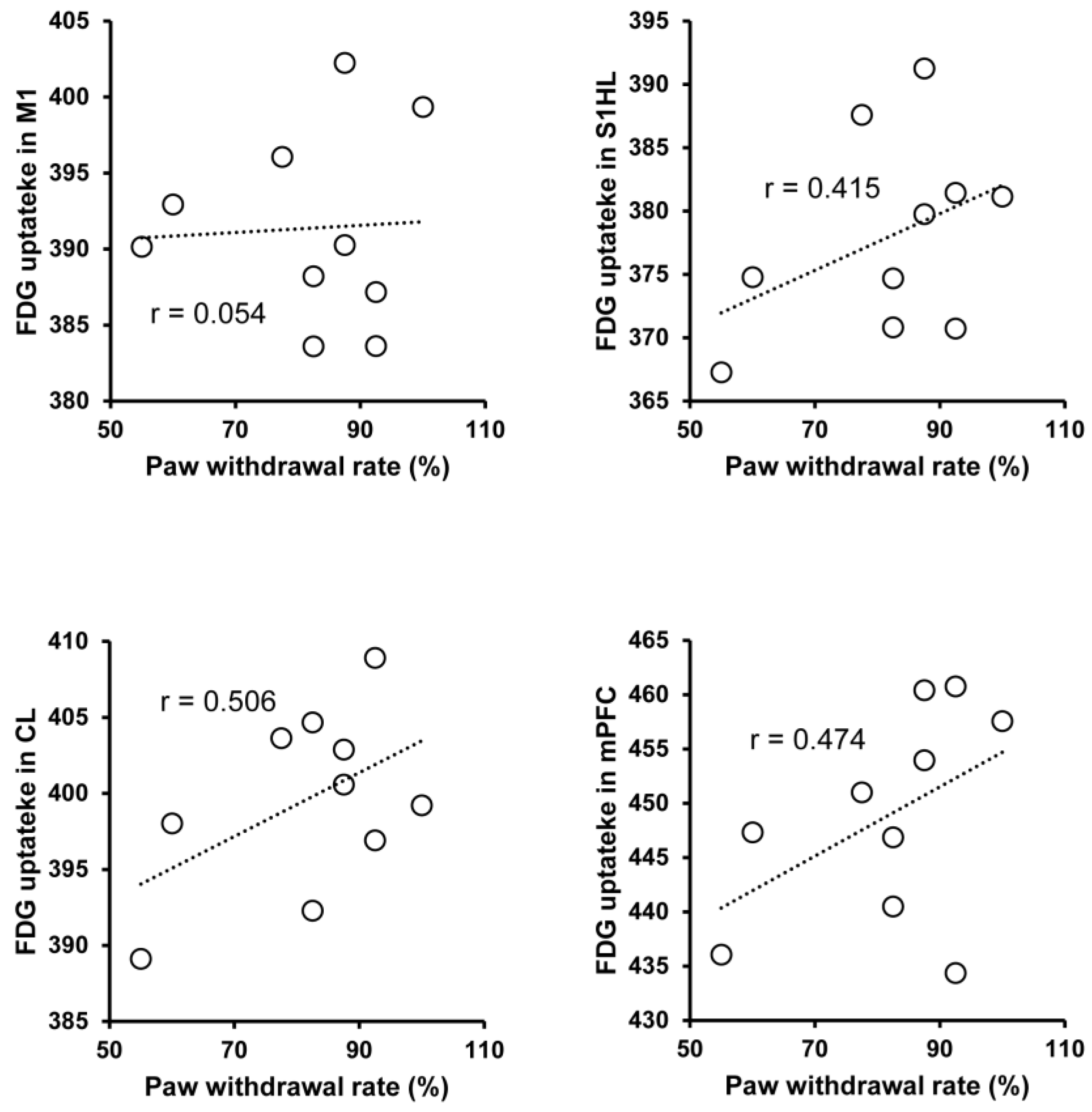
3.3. Regional Brain Activity and Behavior Correlation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Melzack, R.; Casey, K.L. Sensory, Motivational, and Central Control Determinants of Pain: A New Conceptual Model. In The Skin Senses; Kenshalo, D.R., Ed.; Charles, C. Thomas: Springfield, IL, USA, 1968; pp. 423–443. [Google Scholar]
- Tracey, I. Neuroimaging enters the pain biomarker arena. Sci. Transl. Med. 2021, 13, eabj7358. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.D.; Aghaeepour, N.; Ahn, A.H.; Angst, M.S.; Borsook, D.; Brenton, A.; Burczynski, M.E.; Crean, C.; Edwards, R.; Gaudilliere, B.; et al. Discovery and validation of biomarkers to aid the development of safe and effective pain therapeutics: Challenges and opportunities. Nat. Rev. Neurol. 2020, 16, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.D.; Flor, H.; Greely, H.T.; Iannetti, G.D.; Mackey, S.; Ploner, M.; Pustilnik, A.; Tracey, I.; Treede, R.D.; Wager, T.D. Brain imaging tests for chronic pain: Medical, legal and ethical issues and recommendations. Nat. Rev. Neurol. 2017, 13, 624–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martucci, K.T.; Mackey, S.C. Neuroimaging of Pain: Human Evidence and Clinical Relevance of Central Nervous System Processes and Modulation. Anesthesiology 2018, 128, 1241–1254. [Google Scholar] [CrossRef]
- Mouraux, A.; Iannetti, G.D. The search for pain biomarkers in the human brain. Brain J. Neurol. 2018, 141, 3290–3307. [Google Scholar] [CrossRef] [Green Version]
- Moisset, X.; Bouhassira, D. Brain imaging of neuropathic pain. NeuroImage 2007, 37 (Suppl. 1), S80–S88. [Google Scholar] [CrossRef]
- Apkarian, A.V.; Bushnell, M.C.; Treede, R.D.; Zubieta, J.K. Human brain mechanisms of pain perception and regulation in health and disease. Eur. J. Pain 2005, 9, 463–484. [Google Scholar] [CrossRef]
- Baliki, M.N.; Petre, B.; Torbey, S.; Herrmann, K.M.; Huang, L.; Schnitzer, T.J.; Fields, H.L.; Apkarian, A.V. Corticostriatal functional connectivity predicts transition to chronic back pain. Nat. Neurosci. 2012, 15, 1117–1119. [Google Scholar] [CrossRef] [Green Version]
- Tracey, I.; Woolf, C.J.; Andrews, N.A. Composite Pain Biomarker Signatures for Objective Assessment and Effective Treatment. Neuron 2019, 101, 783–800. [Google Scholar] [CrossRef] [Green Version]
- Apkarian, A.V.; Sosa, Y.; Sonty, S.; Levy, R.M.; Harden, R.N.; Parrish, T.B.; Gitelman, D.R. Chronic back pain is associated with decreased prefrontal and thalamic gray matter density. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 10410–10415. [Google Scholar] [CrossRef] [Green Version]
- Peyron, R.; Garcia-Larrea, L.; Gregoire, M.C.; Convers, P.; Lavenne, F.; Veyre, L.; Froment, J.C.; Mauguiere, F.; Michel, D.; Laurent, B. Allodynia after lateral-medullary (Wallenberg) infarct. A PET study. Brain J. Neurol. 1998, 121 Pt 2, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albe-Fessard, D.; Berkley, K.J.; Kruger, L.; Ralston, H.J., 3rd; Willis, W.D., Jr. Diencephalic mechanisms of pain sensation. Brain Res. 1985, 356, 217–296. [Google Scholar] [CrossRef] [PubMed]
- Rainville, P.; Duncan, G.H.; Price, D.D.; Carrier, B.; Bushnell, M.C. Pain affect encoded in human anterior cingulate but not somatosensory cortex. Science 1997, 277, 968–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casey, K.L. Forebrain mechanisms of nociception and pain: Analysis through imaging. Proc. Natl. Acad. Sci. USA 1999, 96, 7668–7674. [Google Scholar] [CrossRef] [Green Version]
- Peyron, R.; Garcia-Larrea, L.; Gregoire, M.C.; Costes, N.; Convers, P.; Lavenne, F.; Mauguiere, F.; Michel, D.; Laurent, B. Haemodynamic brain responses to acute pain in humans: Sensory and attentional networks. Brain J. Neurol. 1999, 122 Pt 9, 1765–1780. [Google Scholar] [CrossRef] [Green Version]
- Bennett, G.J.; Xie, Y.K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef]
- Kim, S.H.; Chung, J.M. An Experimental-Model for Peripheral Neuropathy Produced by Segmental Spinal Nerve Ligation in the Rat. Pain 1992, 50, 355–363. [Google Scholar] [CrossRef]
- Seltzer, Z.; Dubner, R.; Shir, Y. A novel behavioral model of neuropathic pain disorders produced in rats by partial sciatic nerve injury. Pain 1990, 43, 205–218. [Google Scholar] [CrossRef]
- Stephenson, D.T.; Arneric, S.P. Neuroimaging of pain: Advances and future prospects. J. Pain 2008, 9, 567–579. [Google Scholar] [CrossRef]
- Vierck, C.J.; Hansson, P.T.; Yezierski, R.P. Clinical and pre-clinical pain assessment: Are we measuring the same thing? Pain 2008, 135, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Birnie, K.A.; Hundert, A.S.; Lalloo, C.; Nguyen, C.; Stinson, J.N. Recommendations for selection of self-report pain intensity measures in children and adolescents: A systematic review and quality assessment of measurement properties. Pain 2019, 160, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Main, C.J. Pain assessment in context: A state of the science review of the McGill pain questionnaire 40 years on. Pain 2016, 157, 1387–1399. [Google Scholar] [CrossRef] [PubMed]
- Thompson, S.J.; Bushnell, M.C. Rodent functional and anatomical imaging of pain. Neurosci. Lett. 2012, 520, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Borsook, D.; Becerra, L. CNS animal fMRI in pain and analgesia. Neurosci. Biobehav. Rev. 2011, 35, 1125–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.C.; Centeno, M.V.; Procissi, D.; Baria, A.; Apkarian, A.V. Brain activity for tactile allodynia: A longitudinal awake rat functional magnetic resonance imaging study tracking emergence of neuropathic pain. Pain 2017, 158, 488–497. [Google Scholar] [CrossRef]
- Mizuma, H.; Shukuri, M.; Hayashi, T.; Watanabe, Y.; Onoe, H. Establishment of in vivo brain imaging method in conscious mice. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2010, 51, 1068–1075. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.E.; Kim, Y.K.; Chung, G.; Im, H.J.; Lee, D.S.; Kim, J.; Kim, S.J. Identifying neuropathic pain using (18)F-FDG micro-PET: A multivariate pattern analysis. NeuroImage 2014, 86, 311–316. [Google Scholar] [CrossRef]
- Kobayashi, M.; Cui, Y.; Sako, T.; Sasabe, T.; Mizoguchi, N.; Yamamoto, K.; Wada, Y.; Kataoka, Y.; Koshikawa, N. Functional neuroimaging of aversive taste-related areas in the alert rat revealed by positron emission tomography. J. Neurosci. Res. 2013, 91, 1363–1370. [Google Scholar] [CrossRef]
- Sung, K.K.; Jang, D.P.; Lee, S.; Kim, M.; Lee, S.Y.; Kim, Y.B.; Park, C.W.; Cho, Z.H. Neural responses in rat brain during acute immobilization stress: A [F-18]FDG micro PET imaging study. NeuroImage 2009, 44, 1074–1080. [Google Scholar] [CrossRef]
- Zeng, Y.; Hu, D.; Yang, W.; Hayashinaka, E.; Wada, Y.; Watanabe, Y.; Zeng, Q.; Cui, Y. A voxel-based analysis of neurobiological mechanisms in placebo analgesia in rats. NeuroImage 2018, 178, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, W.K.; Mirrione, M.M.; Dewey, S.L. Optimizing experimental protocols for quantitative behavioral imaging with 18F-FDG in rodents. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2007, 48, 277–287. [Google Scholar]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Toyoda, H.; Sako, T.; Onoe, K.; Hayashinaka, E.; Wada, Y.; Yokoyama, C.; Onoe, H.; Kataoka, Y.; Watanabe, Y. A voxel-based analysis of brain activity in high-order trigeminal pathway in the rat induced by cortical spreading depression. NeuroImage 2015, 108, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Coghill, R.C.; Sang, C.N.; Maisog, J.M.; Iadarola, M.J. Pain intensity processing within the human brain: A bilateral, distributed mechanism. J. Neurophysiol. 1999, 82, 1934–1943. [Google Scholar] [CrossRef]
- Saade, N.E.; Al Amin, H.; Abdel Baki, S.; Chalouhi, S.; Jabbur, S.J.; Atweh, S.F. Reversible attenuation of neuropathic-like manifestations in rats by lesions or local blocks of the intralaminar or the medial thalamic nuclei. Exp. Neurol. 2007, 204, 205–219. [Google Scholar] [CrossRef]
- Becerra, L.; Morris, S.; Bazes, S.; Gostic, R.; Sherman, S.; Gostic, J.; Pendse, G.; Moulton, E.; Scrivani, S.; Keith, D.; et al. Trigeminal neuropathic pain alters responses in CNS circuits to mechanical (brush) and thermal (cold and heat) stimuli. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 10646–10657. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Gadotti, V.M.; Chen, L.; Souza, I.A.; Huang, S.; Wang, D.; Ramakrishnan, C.; Deisseroth, K.; Zhang, Z.; Zamponi, G.W. A neuronal circuit for activating descending modulation of neuropathic pain. Nat. Neurosci. 2019, 22, 1659–1668. [Google Scholar] [CrossRef]
- Lorenz, J.; Cross, D.J.; Minoshima, S.; Morrow, T.J.; Paulson, P.E.; Casey, K.L. A unique representation of heat allodynia in the human brain. Neuron 2002, 35, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Gadotti, V.M.; Chen, L.; Souza, I.A.; Stemkowski, P.L.; Zamponi, G.W. Role of Prelimbic GABAergic Circuits in Sensory and Emotional Aspects of Neuropathic Pain. Cell Rep. 2015, 12, 752–759. [Google Scholar] [CrossRef] [Green Version]
- Huo, B.B.; Zheng, M.X.; Hua, X.Y.; Shen, J.; Wu, J.J.; Xu, J.G. Metabolic Brain Network Analysis with (18)F-FDG PET in a Rat Model of Neuropathic Pain. Front. Neurol. 2021, 12, 566119. [Google Scholar] [CrossRef] [PubMed]
- Burns, E.; Chipchase, L.S.; Schabrun, S.M. Primary sensory and motor cortex function in response to acute muscle pain: A systematic review and meta-analysis. Eur. J. Pain 2016, 20, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Schabrun, S.M.; Christensen, S.W.; Mrachacz-Kersting, N.; Graven-Nielsen, T. Motor Cortex Reorganization and Impaired Function in the Transition to Sustained Muscle Pain. Cereb Cortex 2016, 26, 1878–1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.J.; O’Connell, N.E.; Beckenkamp, P.R.; Alhassani, G.; Liston, M.B.; Schabrun, S.M. Altered Primary Motor Cortex Structure, Organization, and Function in Chronic Pain: A Systematic Review and Meta-Analysis. J. Pain 2018, 19, 341–359. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Hao, Y.; Du, M.; Wang, X.; Zhang, J.; Manor, B.; Jiang, X.; Fang, W.; Wang, D. Quantitative cerebral blood flow mapping and functional connectivity of postherpetic neuralgia pain: A perfusion fMRI study. Pain 2013, 154, 110–118. [Google Scholar] [CrossRef]
- Desouza, D.D.; Moayedi, M.; Chen, D.Q.; Davis, K.D.; Hodaie, M. Sensorimotor and Pain Modulation Brain Abnormalities in Trigeminal Neuralgia: A Paroxysmal, Sensory-Triggered Neuropathic Pain. PLoS ONE 2013, 8, e66340. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Kurata, J.; Sekiguchi, M.; Kokubun, M.; Akaishizawa, T.; Chiba, Y.; Konno, S.; Kikuchi, S. Augmented cerebral activation by lumbar mechanical stimulus in chronic low back pain patients: An FMRI study. Spine 2009, 34, 2431–2436. [Google Scholar] [CrossRef]
- Pijnenburg, M.; Brumagne, S.; Caeyenberghs, K.; Janssens, L.; Goossens, N.; Marinazzo, D.; Swinnen, S.P.; Claeys, K.; Siugzdaite, R. Resting-State Functional Connectivity of the Sensorimotor Network in Individuals with Nonspecific Low Back Pain and the Association with the Sit-to-Stand-to-Sit Task. Brain Connect. 2015, 5, 303–311. [Google Scholar] [CrossRef]
- Dolgin, E. Fluctuating baseline pain implicated in failure of clinical trials. Nat. Med. 2010, 16, 1053. [Google Scholar] [CrossRef]
- Thompson, S.J.; Millecamps, M.; Aliaga, A.; Seminowicz, D.A.; Low, L.A.; Bedell, B.J.; Stone, L.S.; Schweinhardt, P.; Bushnell, M.C. Metabolic brain activity suggestive of persistent pain in a rat model of neuropathic pain. NeuroImage 2014, 91, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Okauchi, T.; Hu, D.; Shigeta, M.; Wu, Y.; Wada, Y.; Hayashinaka, E.; Wang, S.; Kogure, Y.; Noguchi, K.; et al. Pain matrix shift in the rat brain following persistent colonic inflammation revealed by voxel-based statistical analysis. Mol. Pain 2019, 15, 1744806919891327. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Huo, B.B.; Hua, X.Y.; Zheng, M.X.; Lu, Y.C.; Wu, J.J.; Shan, C.L.; Xu, J.G. Cerebral (18)F-FDG metabolism alteration in a neuropathic pain model following brachial plexus avulsion: A PET/CT study in rats. Brain Res. 2019, 1712, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Neyama, H.; Nishiyori, M.; Cui, Y.; Watanabe, Y.; Ueda, H. Lysophosphatidic acid receptor type-1 mediates brain activation in micro-positron emission tomography analysis in a fibromyalgia-like mouse model. Eur. J. Neurosci. 2022, 56, 4224–4233. [Google Scholar] [CrossRef] [PubMed]
- Franks, N.P. General anaesthesia: From molecular targets to neuronal pathways of sleep and arousal. Nat. Rev. Neurosci. 2008, 9, 370–386. [Google Scholar] [CrossRef]
- Onoe, H.; Inoue, O.; Suzuki, K.; Tsukada, H.; Itoh, T.; Mataga, N.; Watanabe, Y. Ketamine increases the striatal N-[11C]methylspiperone binding in vivo: Positron emission tomography study using conscious rhesus monkey. Brain Res. 1994, 663, 191–198. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Regions | Laterality | T-Value (Peak) | Volume (mm3) |
|---|---|---|---|
| Anterior olfactory nucleus lateral part (AOL) | R | 3.97 | 0.32 |
| Medial prefrontal cortex (mPFC) | R | 4.45 | 1.49 |
| Cluster 1 | 15.44 | ||
| Primary motor cortex (M1) | R | 5.3 | |
| Primary somatosensory cortex hindlimb region (S1HL) | R | 5.39 | |
| Primary somatosensory cortex barrel field (S1BF)/ Primary somatosensory cortex dysgranular zone (S1DZ) | L | 3.89 | 0.85 |
| Cluster 2 | 7.54 | ||
| Centrolateral thalamic nucleus (CL) | R | 5.35 | |
| Central medial thalamic nucleus (CM) | R | 3.81 | |
| Posterior thalamic nucleus (Po) | R | 6.55 | |
| Posterior thalamic nucleus (Po) | L | 3.98 | 0.34 |
| Cerebellum (CB) | R/L | 5.92 | 30.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Y.; Neyama, H.; Hu, D.; Huang, T.; Hayashinaka, E.; Wada, Y.; Watanabe, Y. FDG PET Imaging of the Pain Matrix in Neuropathic Pain Model Rats. Biomedicines 2023, 11, 63. https://doi.org/10.3390/biomedicines11010063
Cui Y, Neyama H, Hu D, Huang T, Hayashinaka E, Wada Y, Watanabe Y. FDG PET Imaging of the Pain Matrix in Neuropathic Pain Model Rats. Biomedicines. 2023; 11(1):63. https://doi.org/10.3390/biomedicines11010063
Chicago/Turabian StyleCui, Yilong, Hiroyuki Neyama, Di Hu, Tianliang Huang, Emi Hayashinaka, Yasuhiro Wada, and Yasuyoshi Watanabe. 2023. "FDG PET Imaging of the Pain Matrix in Neuropathic Pain Model Rats" Biomedicines 11, no. 1: 63. https://doi.org/10.3390/biomedicines11010063