
Network-Based Assessment of Minimal Change Disease Identifies Glomerular Response to IL-7 and IL-12 Pathways Activation as Innovative Treatment Target
 ,
,  , and
, and 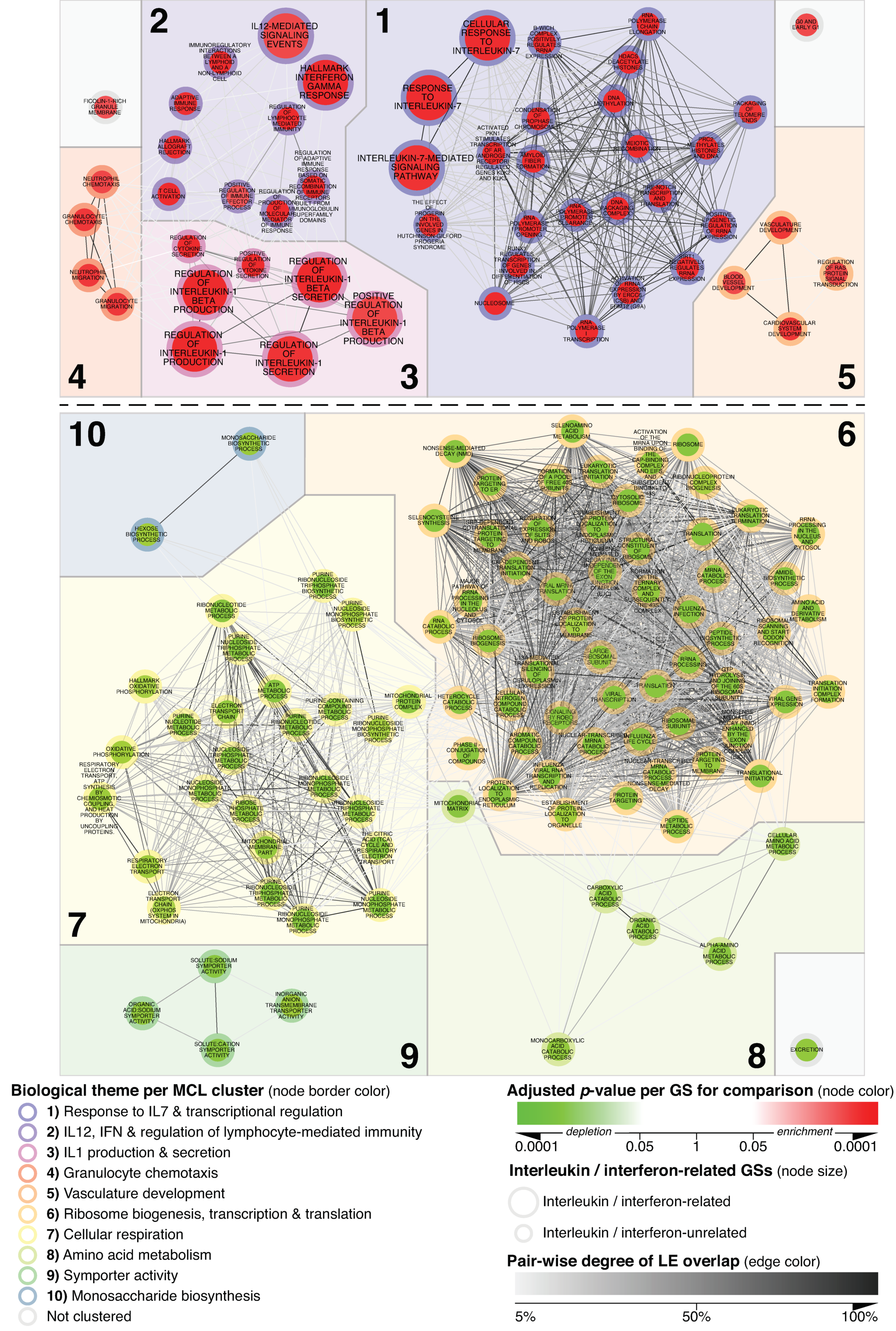{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Patients, Materials and Methods
2.1. Patients
2.2. RNA Purification and Sequencing
2.3. Processing Gene Expression Data Prior to Mapping Transcriptional Landscapes
2.4. Mapping Transcriptional Landscapes
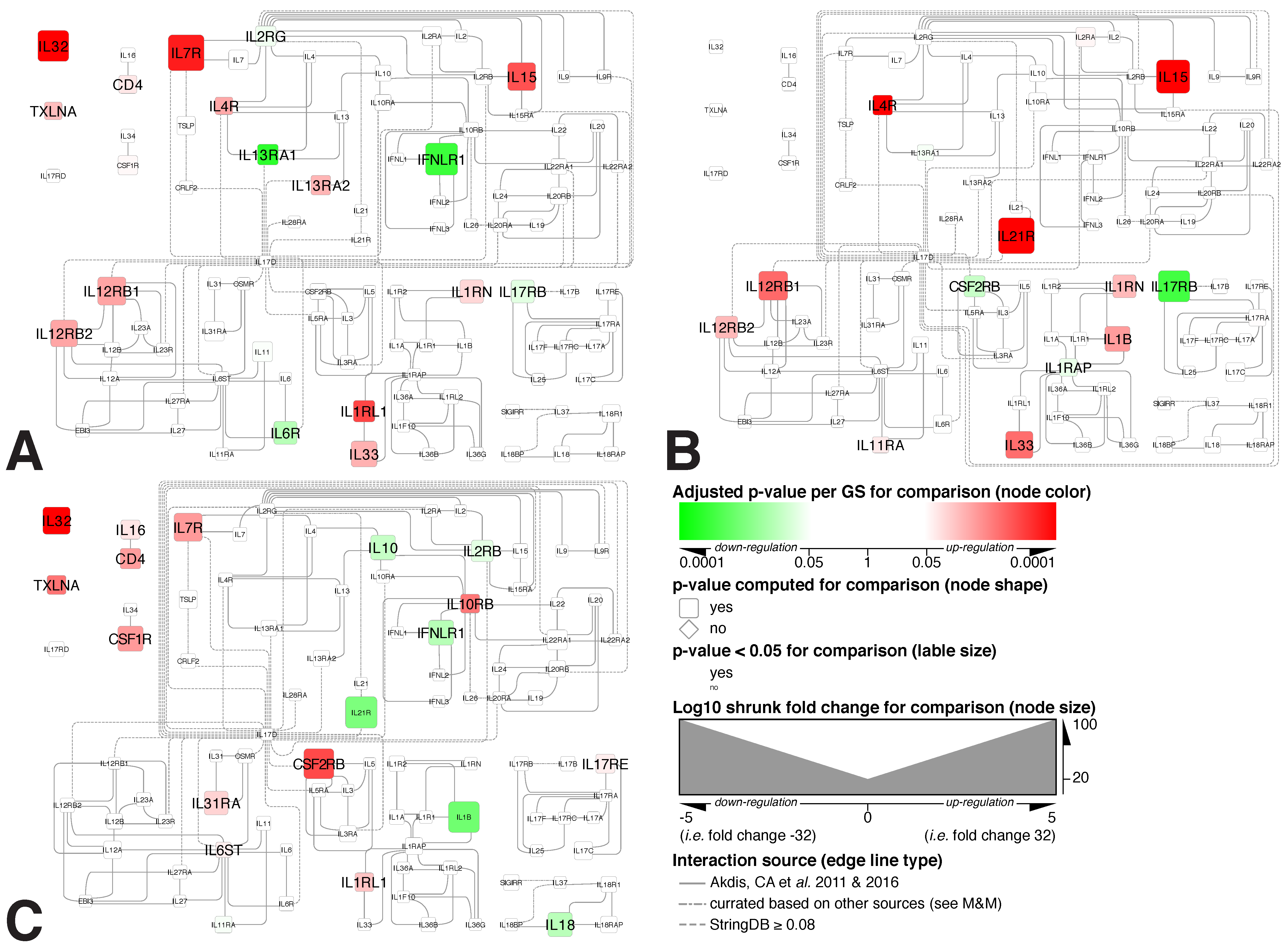
2.5. Interleukin Interactome
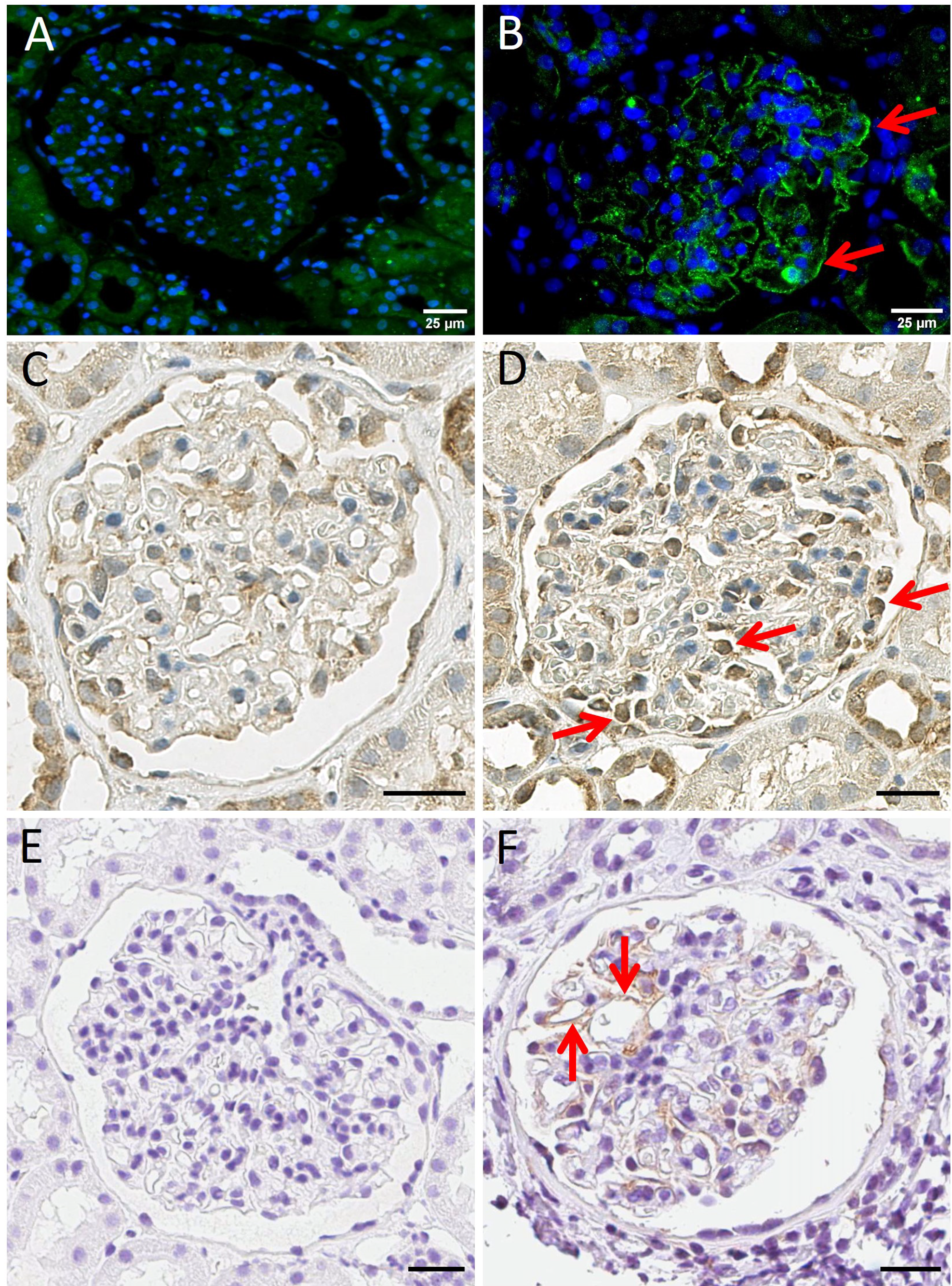
2.6. Immunohistochemistry
3. Results
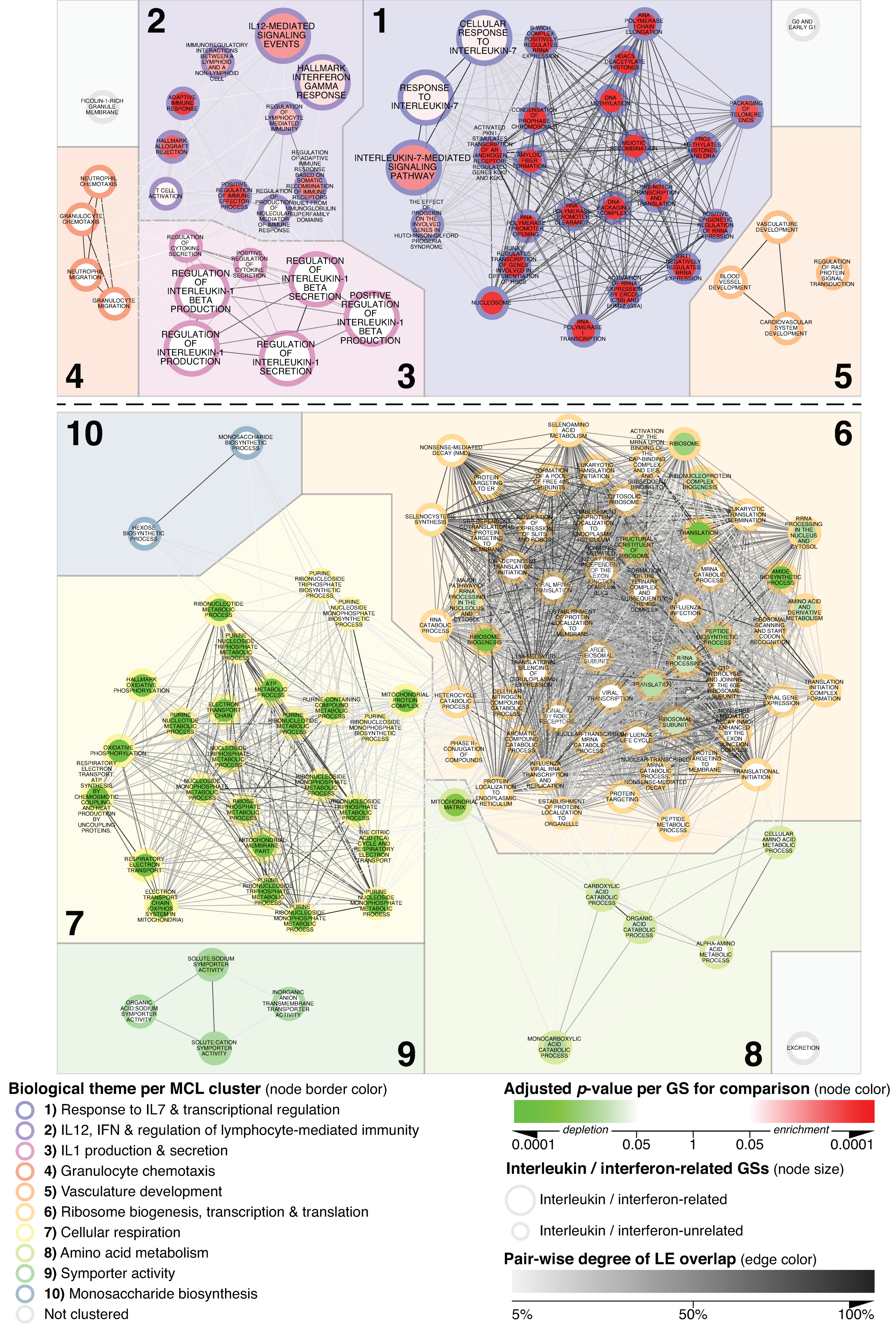
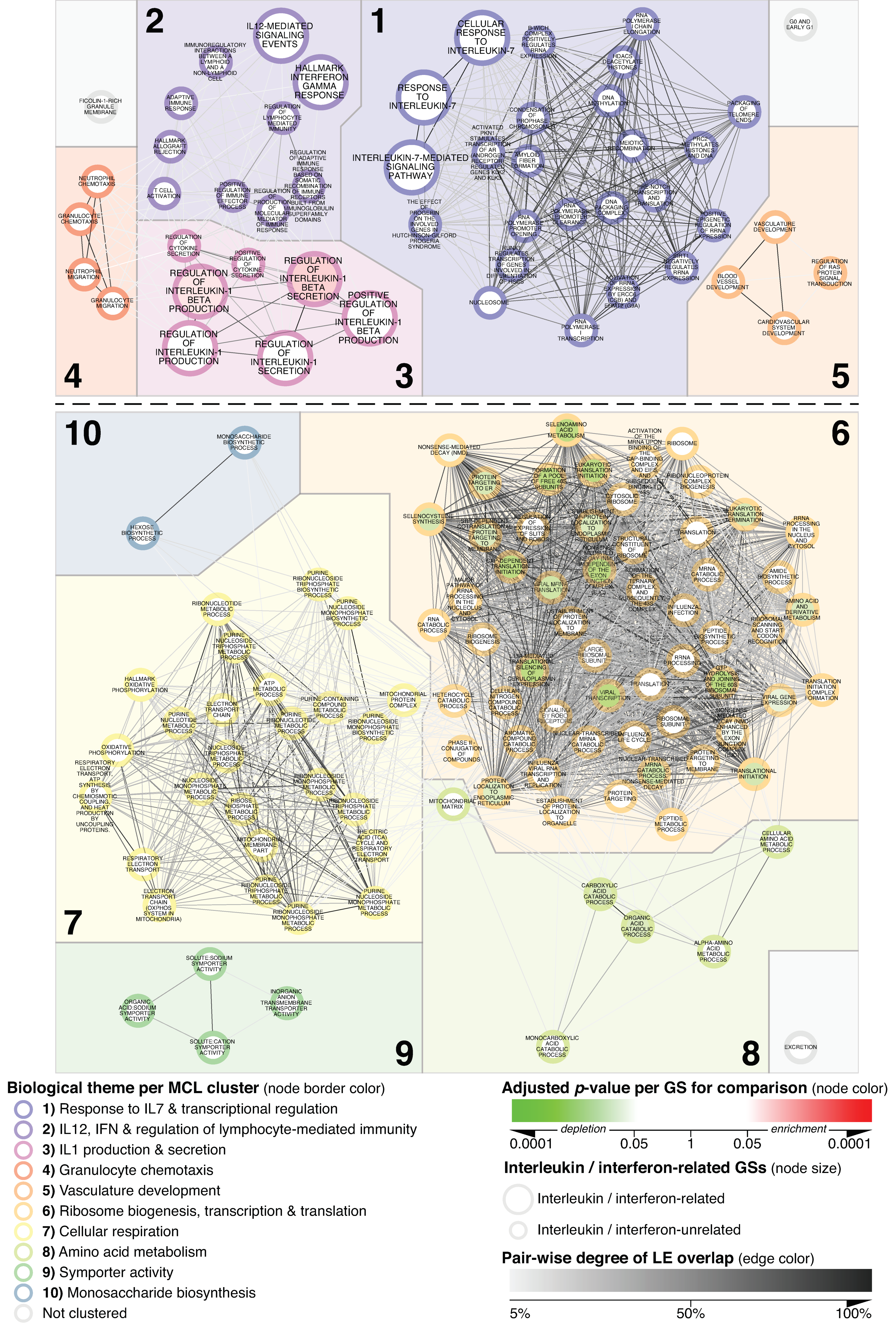
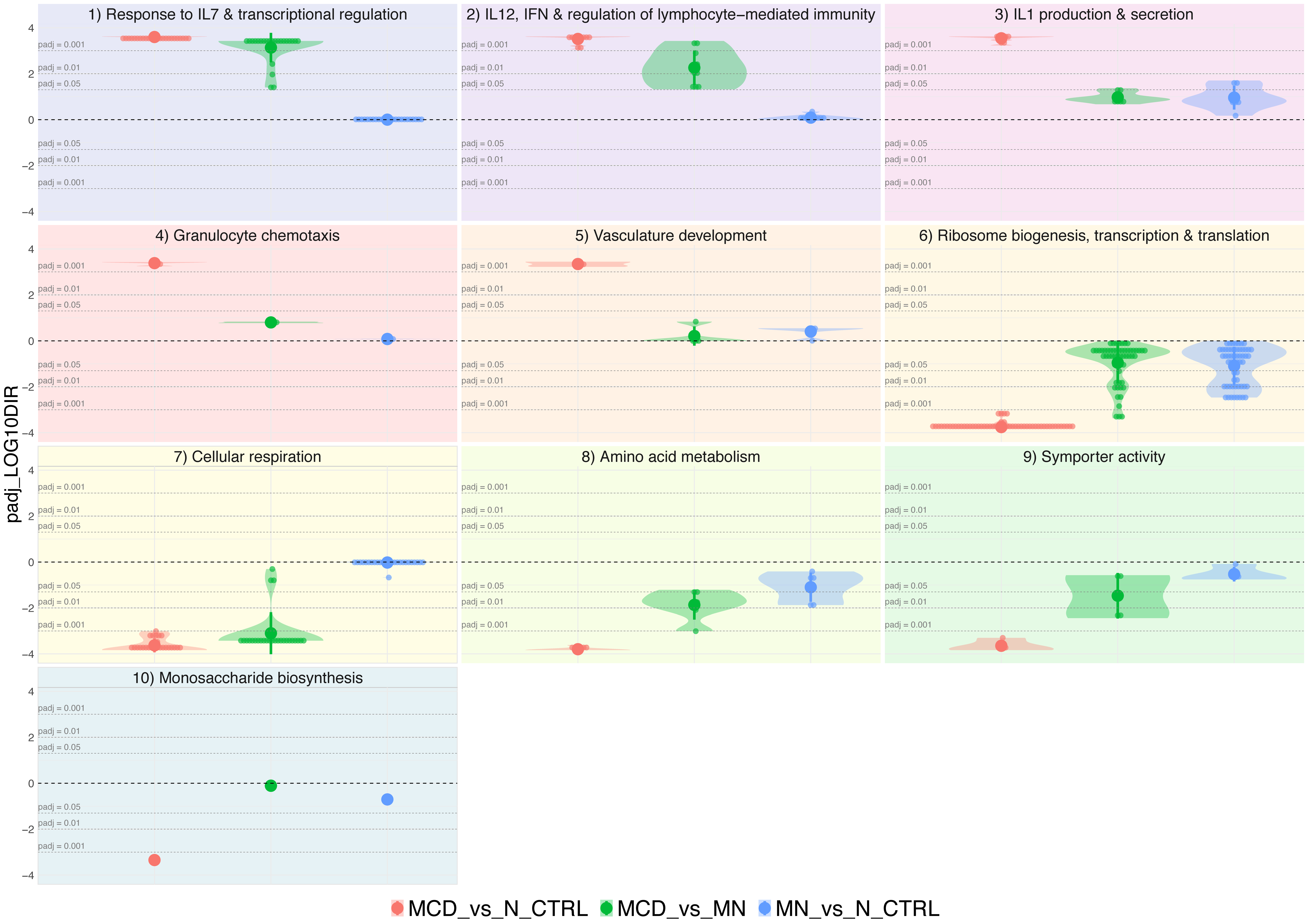
3.1. Network-Based Delineation of MCD’s Glomerular Transcriptional Landscape
3.2. Specificity of Alterations Mapped within the MCD vs. N_CTRL-Focused Network
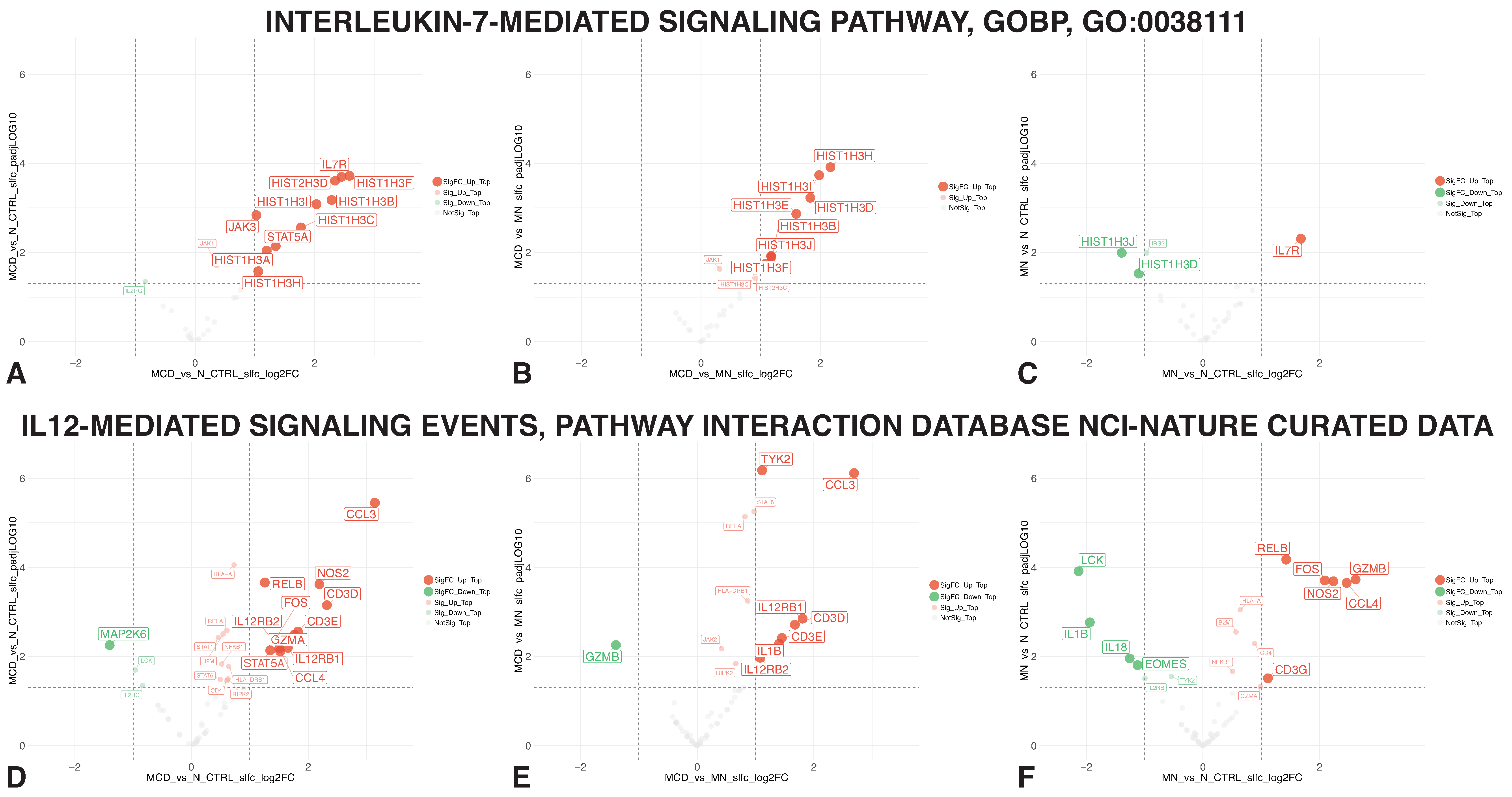
3.3. IL-7 and IL-12-Mediated Signaling Member Genes
3.4. Mapping Alterations within the Interleukin Interactome
3.5. Immunohistochemistry and Immunofluorescence Staining for IL7R, IL12RB2 and Human AXL Tyrosine Kinase Receptor
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vivarelli, M.; Massella, L.; Ruggiero, B.; Emma, F. Minimal Change Disease. Clin. J. Am. Soc. Nephrol. CJASN 2017, 12, 332–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldman, M.; Crew, R.J.; Valeri, A.; Busch, J.; Stokes, B.; Markowitz, G.; D’Agati, V.; Appel, G. Adult minimal-change disease: Clinical characteristics, treatment, and outcomes. Clin. J. Am. Soc. Nephrol. CJASN 2007, 2, 445–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivarelli, M.; Moscaritolo, E.; Tsalkidis, A.; Massella, L.; Emma, F. Time for initial response to steroids is a major prognostic factor in idiopathic nephrotic syndrome. J. Pediatr. 2010, 156, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Shalhoub, R.J. Pathogenesis of lipoid nephrosis: A disorder of T-cell function. Lancet 1974, 2, 556–560. [Google Scholar] [CrossRef]
- Le Berre, L.; Bruneau, S.; Naulet, J.; Renaudin, K.; Buzelin, F.; Usal, C.; Smit, H.; Condamine, T.; Soulillou, J.P.; Dantal, J. Induction of T regulatory cells attenuates idiopathic nephrotic syndrome. J. Am. Soc. Nephrol. JASN 2009, 20, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Hoxha, E.; Stahl, R.A.; Harendza, S. Rituximab in adult patients with immunosuppressive-dependent minimal change disease. Clin. Nephrol. 2011, 76, 151–158. [Google Scholar] [CrossRef]
- Yang, T.; Nast, C.C.; Vo, A.; Jordan, S.C. Rapid remission of steroid and mycophenolate mofetil (mmf)-resistant minimal change nephrotic syndrome after rituximab therapy. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc.-Eur. Ren. Assoc. 2008, 23, 377–380. [Google Scholar] [CrossRef] [Green Version]
- Munyentwali, H.; Bouachi, K.; Audard, V.; Remy, P.; Lang, P.; Mojaat, R.; Deschenes, G.; Ronco, P.M.; Plaisier, E.M.; Dahan, K.Y. Rituximab is an efficient and safe treatment in adults with steroid-dependent minimal change disease. Kidney Int. 2013, 83, 511–516. [Google Scholar] [CrossRef] [Green Version]
- Dabbs, D.J.; Striker, L.M.; Mignon, F.; Striker, G. Glomerular lesions in lymphomas and leukemias. Am. J. Med. 1986, 80, 63–70. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Audard, V.; Fan, Q.; Pawlak, A.; Lang, P.; Sahali, D. Immunopathogenesis of idiopathic nephrotic syndrome. Contrib. Nephrol. 2011, 169, 94–106. [Google Scholar] [CrossRef]
- Pereira Wde, F.; Brito-Melo, G.E.; Guimaraes, F.T.; Carvalho, T.G.; Mateo, E.C.; Simoes e Silva, A.C. The role of the immune system in idiopathic nephrotic syndrome: A review of clinical and experimental studies. Inflamm. Res. 2014, 63, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Van Den Berg, J.G.; Aten, J.; Annink, C.; Ravesloot, J.H.; Weber, E.; Weening, J.J. Interleukin-4 and -13 promote basolateral secretion of H(+) and cathepsin L by glomerular epithelial cells. Am. J. Physiol. Ren. Physiol. 2002, 282, F26–F33. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.W.; Wei, C.L.; Tan, L.K.; Tan, P.H.; Chiang, G.S.; Lee, C.G.; Jordan, S.C.; Yap, H.K. Overexpression of interleukin-13 induces minimal-change-like nephropathy in rats. J. Am. Soc. Nephrol. JASN 2007, 18, 1476–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, S.; Zhao, L.; Zhang, Y.; Ma, Q. Interleukin-7 stimulation inhibits nephrin activation and induces podocyte injury. Biochem. Biophys. Res. Commun. 2018, 507, 100–105. [Google Scholar] [CrossRef]
- Rinschen, M.M.; Godel, M.; Grahammer, F.; Zschiedrich, S.; Helmstadter, M.; Kretz, O.; Zarei, M.; Braun, D.A.; Dittrich, S.; Pahmeyer, C.; et al. A Multi-layered Quantitative In Vivo Expression Atlas of the Podocyte Unravels Kidney Disease Candidate Genes. Cell Rep. 2018, 23, 2495–2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paunas, T.I.F.; Finne, K.; Leh, S.; Marti, H.P.; Mollnes, T.E.; Berven, F.; Vikse, B.E. Glomerular abundance of complement proteins characterized by proteomic analysis of laser-captured microdissected glomeruli associates with progressive disease in IgA nephropathy. Clin. Proteom. 2017, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Wang, Z.; Cao, Y.; Zhang, D.; Mi, X. Bioinformatic Analysis Reveals Novel Immune-Associated Hub Genes in Human Membranous Nephropathy. Genet. Test. Mol. Biomark. 2019, 23, 23–31. [Google Scholar] [CrossRef]
- Williams, A.M.; Jensen, D.M.; Pan, X.; Liu, P.; Liu, J.; Huls, S.; Regner, K.R.; Iczkowski, K.A.; Wang, F.; Li, J.; et al. Histologically resolved small RNA maps in primary focal segmental glomerulosclerosis indicate progressive changes within glomerular and tubulointerstitial regions. Kidney Int. 2022, 101, 766–778. [Google Scholar] [CrossRef]
- Eikrem, O.S.; Strauss, P.; Beisland, C.; Scherer, A.; Landolt, L.; Flatberg, A.; Leh, S.; Beisvag, V.; Skogstrand, T.; Hjelle, K.; et al. Development and confirmation of potential gene classifiers of human clear cell renal cell carcinoma using next-generation RNA sequencing. Scand. J. Urol. 2016, 50, 452–462. [Google Scholar] [CrossRef]
- Landolt, L.; Marti, H.P.; Beisland, C.; Flatberg, A.; Eikrem, O.S. RNA extraction for RNA sequencing of archival renal tissues. Scand. J. Clin. Lab. Investig. 2016, 76, 426–434. [Google Scholar] [CrossRef]
- Nakken, S.; Eikrem, O.; Marti, H.P.; Beisland, C.; Bostad, L.; Scherer, A.; Flatberg, A.; Beisvag, V.; Skandalou, E.; Furriol, J.; et al. AGAP2-AS1 as a prognostic biomarker in low-risk clear cell renal cell carcinoma patients with progressing disease. Cancer Cell Int. 2021, 21, 690. [Google Scholar] [CrossRef]
- Strauss, P.; Rivedal, M.; Scherer, A.; Eikrem, O.; Nakken, S.; Beisland, C.; Bostad, L.; Flatberg, A.; Skandalou, E.; Beisvag, V.; et al. A multiomics disease progression signature of low-risk ccRCC. Sci. Rep. 2022, 12, 13503. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Merico, D.; Isserlin, R.; Stueker, O.; Emili, A.; Bader, G.D. Enrichment map: A network-based method for gene-set enrichment visualization and interpretation. PLoS ONE 2010, 5, e13984. [Google Scholar] [CrossRef] [Green Version]
- Sergushichev, A.A. An algorithm for fast preranked gene set enrichment analysis using cumulative statistic calculation. Biorxiv Prepr. 2016, 60012, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Delaleu, N.; Nguyen, C.Q.; Tekle, K.M.; Jonsson, R.; Peck, A.B. Transcriptional landscapes of emerging autoimmunity: Transient aberrations in the targeted tissue’s extracellular milieu precede immune responses in Sjogren’s syndrome. Arthritis Res. Ther. 2013, 15, R174. [Google Scholar] [CrossRef] [Green Version]
- Emming, S.; Bianchi, N.; Polletti, S.; Balestrieri, C.; Leoni, C.; Montagner, S.; Chirichella, M.; Delaleu, N.; Natoli, G.; Monticelli, S. A molecular network regulating the proinflammatory phenotype of human memory T lymphocytes. Nat. Immunol. 2020, 21, 388–399. [Google Scholar] [CrossRef]
- Di Mitri, D.; Mirenda, M.; Vasilevska, J.; Calcinotto, A.; Delaleu, N.; Revandkar, A.; Gil, V.; Boysen, G.; Losa, M.; Mosole, S.; et al. Re-education of Tumor-Associated Macrophages by CXCR2 Blockade Drives Senescence and Tumor Inhibition in Advanced Prostate Cancer. Cell Rep. 2019, 28, 2156–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Guccini, I.; Di Mitri, D.; Brina, D.; Revandkar, A.; Sarti, M.; Pasquini, E.; Alajati, A.; Pinton, S.; Losa, M.; et al. Compartmentalized activities of the pyruvate dehydrogenase complex sustain lipogenesis in prostate cancer. Nat. Genet. 2018, 50, 219–228. [Google Scholar] [CrossRef]
- Morris, J.H.; Apeltsin, L.; Newman, A.M.; Baumbach, J.; Wittkop, T.; Su, G.; Bader, G.D.; Ferrin, T.E. clusterMaker: A multi-algorithm clustering plugin for Cytoscape. BMC Bioinform. 2011, 12, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, G.; Morris, J.H.; Demchak, B.; Bader, G.D. Biological network exploration with Cytoscape 3. Curr. Protoc. Bioinform. 2014, 47, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akdis, M.; Burgler, S.; Crameri, R.; Eiwegger, T.; Fujita, H.; Gomez, E.; Klunker, S.; Meyer, N.; O’Mahony, L.; Palomares, O.; et al. Interleukins, from 1 to 37, and interferon-gamma: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2011, 127, 701–721.e701–770. [Google Scholar] [CrossRef] [PubMed]
- Akdis, M.; Aab, A.; Altunbulakli, C.; Azkur, K.; Costa, R.A.; Crameri, R.; Duan, S.; Eiwegger, T.; Eljaszewicz, A.; Ferstl, R.; et al. Interleukins (from IL-1 to IL-38), interferons, transforming growth factor beta, and TNF-alpha: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2016, 138, 984–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; He, X.; Li, X.; Qian, Y. The roles and functional mechanisms of interleukin-17 family cytokines in mucosal immunity. Cell. Mol. Immunol. 2016, 13, 418–431. [Google Scholar] [CrossRef] [Green Version]
- Di Carlo, E. Interleukin-30: A novel microenvironmental hallmark of prostate cancer progression. Oncoimmunology 2014, 3, e27618. [Google Scholar] [CrossRef] [Green Version]
- Marrakchi, S.; Guigue, P.; Renshaw, B.R.; Puel, A.; Pei, X.Y.; Fraitag, S.; Zribi, J.; Bal, E.; Cluzeau, C.; Chrabieh, M.; et al. Interleukin-36-receptor antagonist deficiency and generalized pustular psoriasis. N. Engl. J. Med. 2011, 365, 620–628. [Google Scholar] [CrossRef]
- Molgora, M.; Barajon, I.; Mantovani, A.; Garlanda, C. Regulatory Role of IL-1R8 in Immunity and Disease. Front. Immunol. 2016, 7, 149. [Google Scholar] [CrossRef] [Green Version]
- Tonacci, A.; Quattrocchi, P.; Gangemi, S. IL33/ST2 Axis in Diabetic Kidney Disease: A Literature Review. Medicina 2019, 55, 50. [Google Scholar] [CrossRef] [Green Version]
- Landolt, L.; Furriol, J.; Babickova, J.; Ahmed, L.; Eikrem, O.; Skogstrand, T.; Scherer, A.; Suliman, S.; Leh, S.; Lorens, J.B.; et al. AXL targeting reduces fibrosis development in experimental unilateral ureteral obstruction. Physiol. Rep. 2019, 7, e14091. [Google Scholar] [CrossRef]
- Niemir, Z.I.; Stein, H.; Dworacki, G.; Mundel, P.; Koehl, N.; Koch, B.; Autschbach, F.; Andrassy, K.; Ritz, E.; Waldherr, R.; et al. Podocytes are the major source of IL-1 alpha and IL-1 beta in human glomerulonephritides. Kidney Int. 1997, 52, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mentzel, S.; Van Son, J.P.; De Jong, A.S.; Dijkman, H.B.; Koene, R.A.; Wetzels, J.F.; Assmann, K.J. Mouse glomerular epithelial cells in culture with features of podocytes in vivo express aminopeptidase A and angiotensinogen but not other components of the renin-angiotensin system. J. Am. Soc. Nephrol. JASN 1997, 8, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Badot, V.; Luijten, R.K.; van Roon, J.A.; Depresseux, G.; Aydin, S.; Van den Eynde, B.J.; Houssiau, F.A.; Lauwerys, B.R. Serum soluble interleukin 7 receptor is strongly associated with lupus nephritis in patients with systemic lupus erythematosus. Ann. Rheum. Dis. 2013, 72, 453–456. [Google Scholar] [CrossRef] [Green Version]
- Watts, A.J.B.; Keller, K.H.; Lerner, G.; Rosales, I.; Collins, A.B.; Sekulic, M.; Waikar, S.S.; Chandraker, A.; Riella, L.V.; Alexander, M.P.; et al. Discovery of Autoantibodies Targeting Nephrin in Minimal Change Disease Supports a Novel Autoimmune Etiology. J. Am. Soc. Nephrol. JASN 2022, 33, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Schwarting, A.; Tesch, G.; Kinoshita, K.; Maron, R.; Weiner, H.L.; Kelley, V.R. IL-12 drives IFN-gamma-dependent autoimmune kidney disease in MRL-Fas(lpr) mice. J. Immunol. 1999, 163, 6884–6891. [Google Scholar] [CrossRef]
- Bussolati, B.; Mariano, F.; Biancone, L.; Foa, R.; David, S.; Cambi, V.; Camussi, G. Interleukin-12 is synthesized by mesangial cells and stimulates platelet-activating factor synthesis, cytoskeletal reorganization, and cell shape change. Am. J. Pathol. 1999, 154, 623–632. [Google Scholar] [CrossRef] [Green Version]
- Pearce, E.L.; Pearce, E.J. Metabolic pathways in immune cell activation and quiescence. Immunity 2013, 38, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Yanagita, M. Gas6, warfarin, and kidney diseases. Clin. Exp. Nephrol. 2004, 8, 304–309. [Google Scholar] [CrossRef]
- Rea, I.M.; Gibson, D.S.; McGilligan, V.; McNerlan, S.E.; Alexander, H.D.; Ross, O.A. Age and Age-Related Diseases: Role of Inflammation Triggers and Cytokines. Front. Immunol. 2018, 9, 586. [Google Scholar] [CrossRef]
- Ellis, J.; van Maurik, A.; Fortunato, L.; Gisbert, S.; Chen, K.; Schwartz, A.; McHugh, S.; Want, A.; Santos Franco, S.; Oliveira, J.J.; et al. Anti-IL-7 receptor alpha monoclonal antibody (GSK2618960) in healthy subjects—A randomized, double-blind, placebo-controlled study. Br. J. Clin. Pharm. 2019, 85, 304–315. [Google Scholar] [CrossRef]
- Kashani, A.; Schwartz, D.A. The Expanding Role of Anti-IL-12 and/or Anti-IL-23 Antibodies in the Treatment of Inflammatory Bowel Disease. Gastroenterol. Hepatol. 2019, 15, 255–265. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eikrem, Ø.; Lillefosse, B.; Delaleu, N.; Strauss, P.; Osman, T.; Vikse, B.E.; Debiec, H.; Ronco, P.; Sekulic, M.; Koch, E.; et al. Network-Based Assessment of Minimal Change Disease Identifies Glomerular Response to IL-7 and IL-12 Pathways Activation as Innovative Treatment Target. Biomedicines 2023, 11, 226. https://doi.org/10.3390/biomedicines11010226
Eikrem Ø, Lillefosse B, Delaleu N, Strauss P, Osman T, Vikse BE, Debiec H, Ronco P, Sekulic M, Koch E, et al. Network-Based Assessment of Minimal Change Disease Identifies Glomerular Response to IL-7 and IL-12 Pathways Activation as Innovative Treatment Target. Biomedicines. 2023; 11(1):226. https://doi.org/10.3390/biomedicines11010226
Chicago/Turabian StyleEikrem, Øystein, Bjørnar Lillefosse, Nicolas Delaleu, Philipp Strauss, Tarig Osman, Bjørn Egil Vikse, Hanna Debiec, Pierre Ronco, Miroslav Sekulic, Even Koch, and et al. 2023. "Network-Based Assessment of Minimal Change Disease Identifies Glomerular Response to IL-7 and IL-12 Pathways Activation as Innovative Treatment Target" Biomedicines 11, no. 1: 226. https://doi.org/10.3390/biomedicines11010226