

Osteopontin and Cancer: Insights into Its Role in Drug Resistance


Abstract
:1. Introduction
2. OPN Expression for the Evaluation of Response to Cancer Treatment
2.1. OPN Expression and Response to Radiation Therapies
2.2. OPN Expression in Tumours and Response to Chemotherapies
2.3. Serum Levels of OPN and Patients’ Responses to Treatment

{kind=link}
{kind=link}
| Cancer Types | Study | Inclusion Year | Sample Size | Cancer Stage | Therapeutic Regimens | Test Method | Treatment Response | n, (%) | OPN Expression | p Value | HR/OR (95% CI) | p Value | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low n, (%) | High n, (%) | ||||||||||||
| Cervical SCC | Feng et al. 2018 [13] | January 2015–December 2015 | 116 | IIB-IIIB | IMRT + platinum-based chemotherapy | IHC | Sensitive (CR) | 97 (84%) | 41 (95%) | 56 (77%) | 0.009 | ||
| Resistant (PR + MR + PD) | 19 (16%) | 2 (5%) | 17 (23%) | ||||||||||
| NSCLC | Huang et al. 2018 [14] | March 2010–August 2016 | 73 | I-IV | First-line platinum-based chemotherapy | IHC | PR | 27 (37%) | 12 (38%) | 15 (37%) | 0.038 | 2.326 (1.721–2.616) | 0.005 |
| SD | 38 (52%) | 18 (56%) | 20 (49%) | ||||||||||
| PD | 8 (11%) | 2 (6%) | 6 (15%) | ||||||||||
| OSCC | Luo et al. 2015 [16] | 2006–2012 | 121 | IV | IC + CRRT | IHC | CR/PR | 52 (43%) | 19 (70%) | 33 (35%) | 0.002 | 0.320 (0.120–0.854) | 0.023 |
| SD/PD | 69 (57%) | 8 (30%) | 61 (65%) | ||||||||||
| PC | Aksoy et al. 2017 [17] | 2009–2015 | 30 | IV | TBR | IHC | Responders | 12 (40%) | 1 (20%) | 11 (44%) | 0.622 | ||
| Non-responders | 18 (60%) | 4 (80%) | 14 (56%) | ||||||||||
| LACSCC | Huang et al. 2015 [16] | January 2005–March 2012 | 111 | Ib-Iva (FIGO) | Primary radical radiotherapy | IHC | Radiation-sensitive | 85 (77%) | 59 (95%) | 26 (53%) | <0.001 | ||
| Radiation-resistant | 26 (23%) | 3 (5%) | 23 (47%) | ||||||||||
| Cancer Type | Study | Inclusion Year | Sample Size | Cancer Stage | Therapeutic Regimens | Test Method | Treatment Response | n | OPN Expression | p Value |
|---|---|---|---|---|---|---|---|---|---|---|
| Cervical cancer | Feng et al. 2018 [13] | January 2015–December 2015 | 116 | IIB-IIIB | IMRT + platinum-based chemotherapy | ELISA | Sensitive (CR) | 97 (84%) | 74.01 ± 27.95 (ng/mL) | <0.001 |
| Resistant (PR + MR + PD) | 19 (16%) | 116.98 ± 10.36 (ng/mL) | ||||||||
| SCLC | Xu et al. 2020 [20] | _ | 96 | _ | First-line chemotherapy (EP/EC) | ELISA | CR + PR | 78 (81%) | 71.15 ± 18.33 (ng/mL) | 0.485 |
| SD + PD | 18 (19%) | 78.01 ± 13.74 (ng/mL) | ||||||||
| Metastatic breast cancer | Elbaiomy et al. 2020 [18] | January 2017–March 2019 | 115 | IV | First-line chemotherapy (DC) | ELISA | CR, PR, and SD | 41 (36%) | 35.5 ± 34.3 (ng/dL) | 0.05 |
| PD | 73 (64%) | 49.1 ± 33.8 (ng/dL) | ||||||||
| GC | Yazici et al. 2020 [9] | 2009–2015 | 42 | IV | Modified DCF | ELISA | Responders | _ | 110.7 ± 29.3 (ng/mL) | 0.002 |
| Non-responders | _ | 211.9 ± 24.4 (ng/mL) | ||||||||
| OSCC | Nakamura et al. 2014 [19] | 1999–2004 | 49 | I-IV | 5-FU-based chemoradiotherapy | qRT-PCR | Sensitive | 30 (61%) | OPN overexpression in OSCC tissues with resistance to 5-FU-based chemoradiotherapy | <0.05 |
| Resistant | 19 (39%) |
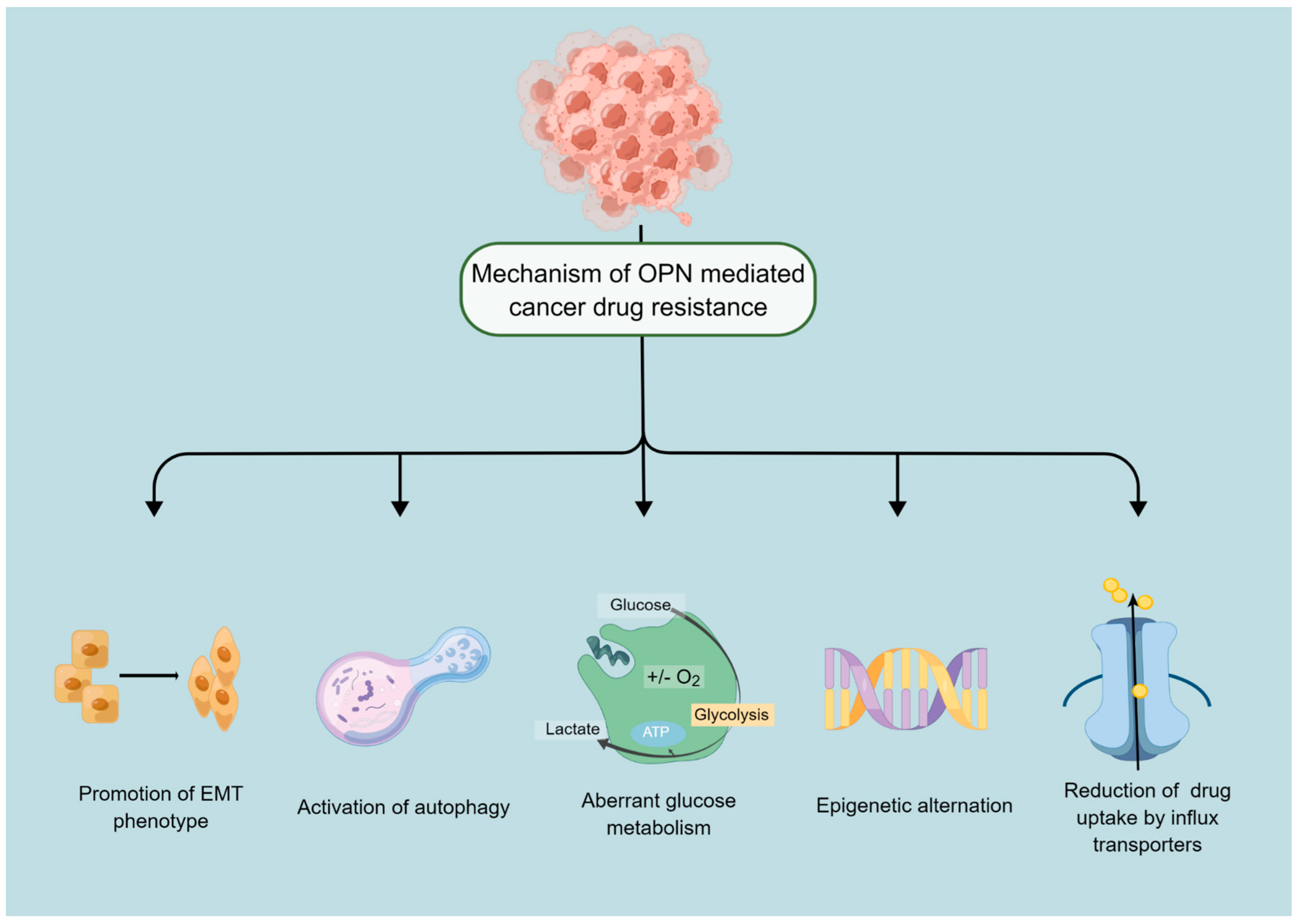
3. Mechanism of OPN Mediated Anticancer Drug Resistance
3.1. OPN Is a Potential Regulator of Autophagy
3.2. OPN and Drug Efflux of Cancer Cells
3.3. Epigenetic Changes
3.4. OPN Is a Prime Regulator of EMT Process
3.5. OPN Critically Regulates the Glycolytic Process
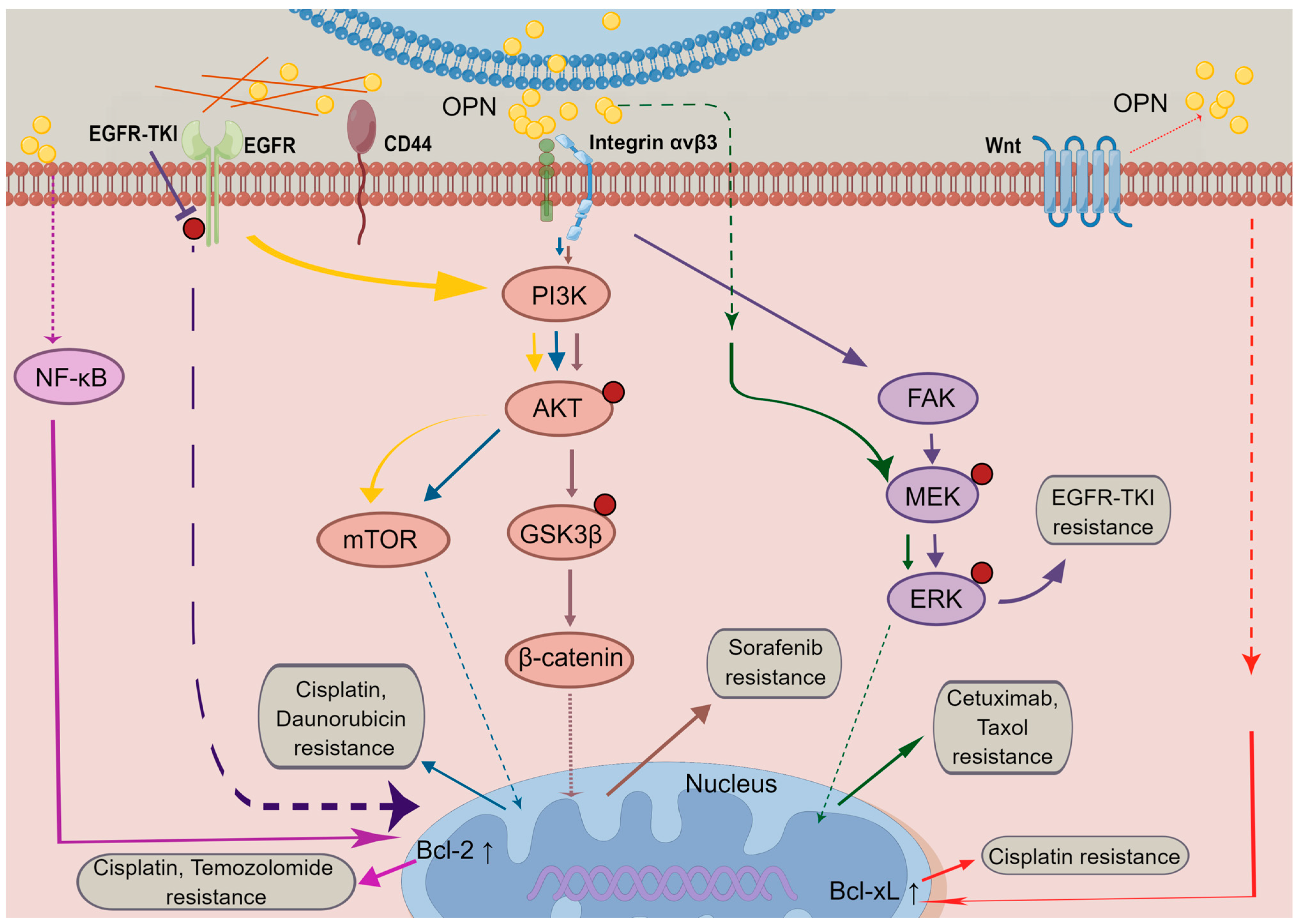
4. OPN Mediates Signalling Pathways in Cancer Resistance
4.1. PI3K/Akt Signalling
4.2. MAPK Signalling
4.3. EGFR Signalling
5. OPN Alternative Splicing in Cancer Drug Resistance
6. Future Prospective
6.1. Questions to Be Addressed in Future Studies
6.2. OPN and Combination Therapies
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xie, K.; Zhou, H.; Wu, Y.; Li, C.; Liu, Y.; Liu, Z.; Xu, Q.; Liu, S.; Xiao, D.; et al. Role of non-coding RNAs and RNA modifiers in cancer therapy resistance. Mol. Cancer 2020, 19, 47. [Google Scholar] [CrossRef] [Green Version]
- Dagogo-Jack, I.; Shaw, A.T. Tumour heterogeneity and resistance to cancer therapies. Nat. Rev. Clin. Oncol. 2018, 15, 81–94. [Google Scholar] [CrossRef]
- Juchum, M.; Gunther, M.; Laufer, S.A. Fighting cancer drug resistance: Opportunities and challenges for mutation-specific EGFR inhibitors. Drug Resist. Updat. 2015, 20, 12–28. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Shevde, L.A.; Samant, R.S. Role of osteopontin in the pathophysiology of cancer. Matrix Biol. 2014, 37, 131–141. [Google Scholar] [CrossRef]
- Anborgh, P.H.; Caria, L.B.; Chambers, A.F.; Tuck, A.B.; Stitt, L.W.; Brackstone, M. Role of plasma osteopontin as a biomarker in locally advanced breast cancer. Am. J. Transl. Res. 2015, 7, 723–732. [Google Scholar]
- Hao, C.; Cui, Y.; Chang, S.; Huang, J.; Birkin, E.; Hu, M.; Zhi, X.; Li, W.; Zhang, L.; Cheng, S.; et al. OPN promotes the aggressiveness of non-small-cell lung cancer cells through the activation of the RON tyrosine kinase. Sci. Rep. 2019, 9, 18101. [Google Scholar] [CrossRef] [Green Version]
- Yazici, O.; Dogan, M.; Ozal, G.; Aktas, S.H.; Demirkazik, A.; Utkan, G.; Senler, F.C.; Icli, F.; Akbulut, H. Osteopontin is a prognostic factor in patients with advanced gastric cancer. Comb. Chem. High Throughput Screen. 2020, 24, 1229–1235. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, Q.; Alam, A.; Cui, J.; Suen, K.C.; Soo, A.P.; Eguchi, S.; Gu, J.; Ma, D. The role of osteopontin in the progression of solid organ tumour. Cell Death Dis. 2018, 9, 356. [Google Scholar] [CrossRef]
- Yan, C.H.; Lv, M.; Li, H.; Song, X.; Yan, F.; Cao, S.; Ren, X. Osteopontin is a novel prognostic biomarker in early-stage non-small cell lung cancer after surgical resection. J. Cancer Res. Clin. Oncol. 2015, 141, 1371–1378. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, J.W.; Woo, S.H.; Go, D.M.; Kwon, H.J.; Jang, J.J.; Kim, D.Y. Suppression of osteopontin inhibits chemically induced hepatic carcinogenesis by induction of apoptosis in mice. Oncotarget 2016, 7, 87219–87231. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.-W.; Du, X.-L.; Wang, C.; Shan, T.-J. Association between osteopontin and matrix metalloproteinase-9 levels and sensitivity of chemoradiotherapy in advanced cervical cancer. Chin. J. Cancer Prev. Treat. 2018, 25, 717–724. [Google Scholar] [CrossRef]
- Huang, X.; Qian, Y.; Wu, H.; Xie, X.; Zhou, Q.; Wang, Y.; Kuang, W.; Shen, L.; Li, K.; Su, J.; et al. Aberrant expression of osteopontin and E-cadherin indicates radiation resistance and poor prognosis for patients with cervical carcinoma. J. Histochem. Cytochem. 2015, 63, 88–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, X.; Huang, Y.; Jin, X.; Zhao, W.; Hu, T.; Wu, F.; Huang, J. Osteopontin promotes cancer cell drug resistance, invasion, and lactate production and is associated with poor outcome of patients with advanced non-small-cell lung cancer. OncoTargets Ther. 2018, 11, 5933–5941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.D.; Chen, Y.J.; Liu, C.T.; Rau, K.M.; Chen, Y.C.; Tsai, H.T.; Chen, C.H.; Chiu, T.J. Osteopontin Involves Cisplatin Resistance and Poor Prognosis in Oral Squamous Cell Carcinoma. BioMed Res. Int. 2015, 2015, 508587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksoy, A.; Artas, G.; Sevindik, O.G. Predictive value of stathmin-1 and osteopontin expression for taxan resistance in metastatic castrate-resistant prostate cancer. Pak. J. Med. Sci. 2017, 33, 560–565. [Google Scholar] [CrossRef]
- Elbaiomy, M.A.; Akl, T.; Elhelaly, R.; El-Beshbishi, W.; El Ghonemy, M.S.; Elzehery, R. Osteopontin level and promoter polymorphism in patients with metastatic breast cancer. Curr. Oncol. 2020, 27, 444–450. [Google Scholar] [CrossRef]
- Nakamura, T.; Shinriki, S.; Jono, H.; Ueda, M.; Nagata, M.; Guo, J.; Hayashi, M.; Yoshida, R.; Ota, T.; Ota, K.; et al. Osteopontin-integrin α(v)β(3) axis is crucial for 5-fluorouracil resistance in oral squamous cell carcinoma. FEBS Lett. 2015, 589, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Yuan, Q.; Wang, W.; Chi, C.; Zhang, Q.; Li, L.; Yang, R.; Wang, Y. Prognostic significance of serum osteopontin levels in small cell lung cancer. BMC Pulm. Med. 2020, 20, 235. [Google Scholar] [CrossRef]
- Boya, P.; Reggiori, F.; Codogno, P. Emerging regulation and functions of autophagy. Nat. Cell Biol. 2013, 15, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Pietrocola, F.; Bravo-San Pedro, J.M.; Amaravadi, R.K.; Baehrecke, E.H.; Cecconi, F.; Codogno, P.; Debnath, J.; Gewirtz, D.A.; Karantza, V.; et al. Autophagy in malignant transformation and cancer progression. EMBO J. 2015, 34, 856–880. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, S.; Ma, B. Autophagy and autophagy-related proteins in cancer. Mol. Cancer 2020, 19, 12. [Google Scholar] [CrossRef] [PubMed]
- Firat, E.; Weyerbrock, A.; Gaedicke, S.; Grosu, A.L.; Niedermann, G. Chloroquine or chloroquine-PI3K/Akt pathway inhibitor combinations strongly promote gamma-irradiation-induced cell death in primary stem-like glioma cells. PLoS ONE 2012, 7, e47357. [Google Scholar] [CrossRef]
- Qin, W.; Li, C.; Zheng, W.; Guo, Q.; Zhang, Y.; Kang, M.; Zhang, B.; Yang, B.; Li, B.; Yang, H.; et al. Inhibition of autophagy promotes metastasis and glycolysis by inducing ROS in gastric cancer cells. Oncotarget 2015, 6, 39839–39854. [Google Scholar] [CrossRef] [Green Version]
- Battista, R.A.; Resnati, M.; Facchi, C.; Ruggieri, E.; Cremasco, F.; Paradiso, F.; Orfanelli, U.; Giordano, L.; Bussi, M.; Cenci, S.; et al. Autophagy mediates epithelial cancer chemoresistance by reducing p62/SQSTM1 accumulation. PLoS ONE 2018, 13, e0201621. [Google Scholar] [CrossRef]
- Soni, M.; Patel, Y.; Markoutsa, E.; Jie, C.; Liu, S.; Xu, P.; Chen, H. Autophagy, Cell Viability, and Chemoresistance Are Regulated By miR-489 in Breast Cancer. Mol. Cancer Res. 2018, 16, 1348–1360. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.W.; Kang, K.A.; Piao, M.J.; Ryu, Y.S.; Fernando, P.; Oh, M.C.; Park, J.E.; Shilnikova, K.; Na, S.Y.; Jeong, S.U.; et al. Reduced Autophagy in 5-Fluorouracil Resistant Colon Cancer Cells. Biomol. Ther. 2017, 25, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Zhang, C.; Yu, B.; Chen, B.; Liu, Z.; Hou, C.; Wang, F.; Shen, H.; Chen, Z. Autophagic degradation of FOXO3a represses the expression of PUMA to block cell apoptosis in cisplatin-resistant osteosarcoma cells. Am. J. Cancer Res. 2017, 7, 1407–1422. [Google Scholar]
- Datta, S.; Choudhury, D.; Das, A.; Mukherjee, D.D.; Dasgupta, M.; Bandopadhyay, S.; Chakrabarti, G. Autophagy inhibition with chloroquine reverts paclitaxel resistance and attenuates metastatic potential in human nonsmall lung adenocarcinoma A549 cells via ROS mediated modulation of beta-catenin pathway. Apoptosis 2019, 24, 414–433. [Google Scholar] [CrossRef]
- Zheng, Y.H.; Tian, C.; Meng, Y.; Qin, Y.W.; Du, Y.H.; Du, J.; Li, H.H. Osteopontin stimulates autophagy via integrin/CD44 and p38 MAPK signaling pathways in vascular smooth muscle cells. J. Cell. Physiol. 2012, 227, 127–135. [Google Scholar] [CrossRef]
- Li, Y.; Guo, T.; Zhang, Z.; Yao, Y.; Chang, S.; Nor, J.E.; Clarkson, B.H.; Ni, L.; Liu, J. Autophagy Modulates Cell Mineralization on Fluorapatite-Modified Scaffolds. J. Dent. Res. 2016, 95, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Fan, X.; Tang, M.; Chen, R.; Wang, H.; Jia, R.; Zhou, X.; Jing, W.; Wang, H.; Yang, Y.; et al. Osteopontin induces autophagy to promote chemo-resistance in human hepatocellular carcinoma cells. Cancer Lett. 2016, 383, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.C.; Wang, H.C.; Hou, Y.C.; Tung, H.L.; Chiu, T.J.; Shan, Y.S. Blockade of autophagy reduces pancreatic cancer stem cell activity and potentiates the tumoricidal effect of gemcitabine. Mol. Cancer 2015, 14, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.H.; Minai-Tehrani, A.; Shin, J.Y.; Park, S.; Kim, J.E.; Yu, K.N.; Hong, S.H.; Hong, C.M.; Lee, K.H.; Beck, G.R., Jr.; et al. Beclin1-induced autophagy abrogates radioresistance of lung cancer cells by suppressing osteopontin. J. Radiat. Res. 2012, 53, 422–432. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.H.; Vastermark, A.; Shlykov, M.A.; Reddy, V.; Sun, E.I.; Saier, M.H., Jr. Evolutionary relationships of ATP-Binding Cassette (ABC) uptake porters. BMC Microbiol. 2013, 13, 98. [Google Scholar] [CrossRef] [Green Version]
- Locher, K.P. Mechanistic diversity in ATP-binding cassette (ABC) transporters. Nat. Struct. Mol. Biol. 2016, 23, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef]
- Domenichini, A.; Adamska, A.; Falasca, M. ABC transporters as cancer drivers: Potential functions in cancer development. Biochim. Biophys. Acta. Gen. Subj. 2019, 1863, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Zheng, Y.; Ma, L.; Tian, L.; Sun, Q. Clinically-Relevant ABC Transporter for Anti-Cancer Drug Resistance. Front. Pharmacol. 2021, 12, 648407. [Google Scholar] [CrossRef]
- Ambjorner, S.E.B.; Wiese, M.; Kohler, S.C.; Svindt, J.; Lund, X.L.; Gajhede, M.; Saaby, L.; Brodin, B.; Rump, S.; Weigt, H.; et al. The Pyrazolo [3,4-d]pyrimidine Derivative, SCO-201, Reverses Multidrug Resistance Mediated by ABCG2/BCRP. Cells 2020, 9, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, I.S.; Huang, W.H.; Liou, H.C.; Chuang, W.J.; Yang, R.S.; Fu, W.M. Upregulation of drug transporter expression by osteopontin in prostate cancer cells. Mol. Pharmacol. 2013, 83, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sharma, P.; Kumar, D.; Chakraborty, G.; Gorain, M.; Kundu, G.C. Functional characterization of stromal osteopontin in melanoma progression and metastasis. PLoS ONE 2013, 8, e69116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Shilatifard, A. Epigenetic modifications of histones in cancer. Genome Biol. 2019, 20, 245. [Google Scholar] [CrossRef] [Green Version]
- Kanwal, R.; Gupta, S. Epigenetic modifications in cancer. Clin. Genet. 2012, 81, 303–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilting, R.H.; Dannenberg, J.H. Epigenetic mechanisms in tumorigenesis, tumor cell heterogeneity and drug resistance. Drug Resist. Updat. 2012, 15, 21–38. [Google Scholar] [CrossRef] [Green Version]
- Zeller, C.; Brown, R. Therapeutic modulation of epigenetic drivers of drug resistance in ovarian cancer. Ther. Adv. Med. Oncol. 2010, 2, 319–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammad, H.P.; Barbash, O.; Creasy, C.L. Targeting epigenetic modifications in cancer therapy: Erasing the roadmap to cancer. Nat. Med. 2019, 25, 403–418. [Google Scholar] [CrossRef]
- Gao, X.; Sheng, Y.; Yang, J.; Wang, C.; Zhang, R.; Zhu, Y.; Zhang, Z.; Zhang, K.; Yan, S.; Sun, H.; et al. Osteopontin alters DNA methylation through up-regulating DNMT1 and sensitizes CD133+/CD44+ cancer stem cells to 5 azacytidine in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 179. [Google Scholar] [CrossRef]
- Si, W.; Shen, J.; Zheng, H.; Fan, W. The role and mechanisms of action of microRNAs in cancer drug resistance. Clin. Epigenetics 2019, 11, 25. [Google Scholar] [CrossRef]
- Iorio, M.V.; Croce, C.M. MicroRNAs in cancer: Small molecules with a huge impact. J. Clin. Oncol. 2009, 27, 5848–5856. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.E.; Weiss, G.J. Resistance may not be futile: microRNA biomarkers for chemoresistance and potential therapeutics. Mol. Cancer Ther. 2010, 9, 3126–3136. [Google Scholar] [CrossRef] [PubMed]
- Shevde, L.A.; Metge, B.J.; Mitra, A.; Xi, Y.; Ju, J.; King, J.A.; Samant, R.S. Spheroid-forming subpopulation of breast cancer cells demonstrates vasculogenic mimicry via hsa-miR-299-5p regulated de novo expression of osteopontin. J. Cell. Mol. Med. 2010, 14, 1693–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.; Huang, J.; Han, Y.; Hao, J.; Wu, X.; Song, H.; Chen, X.; Shen, Q.; Dong, X.; Pang, H.; et al. The microRNA miR-181c enhances chemosensitivity and reduces chemoresistance in breast cancer cells via down-regulating osteopontin. Int. J. Biol. Macromol. 2019, 125, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Jiang, X.; Chen, L.; Zhao, Y.; Huang, Z.; Zhou, H.; Shi, M. MiR-181a Promotes Apoptosis and Reduces Cisplatin Resistance by Inhibiting Osteopontin in Cervical Cancer Cells. Cancer Biother. Radiopharm. 2019, 34, 559–565. [Google Scholar] [CrossRef]
- Sleeman, J.P.; Thiery, J.P. SnapShot: The epithelial-mesenchymal transition. Cell 2011, 145, 162–162.e1. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Weinberg, R.A. Epithelial—Mesenchymal Plasticity: A Central Regulator of Cancer Progression. Trends Cell Biol. 2015, 25, 675–686. [Google Scholar] [CrossRef] [Green Version]
- Moustakas, A.; de Herreros, A.G. Epithelial-mesenchymal transition in cancer. Mol. Oncol. 2017, 11, 715–717. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, D.M.; Medici, D. Signaling mechanisms of the epithelial-mesenchymal transition. Sci. Signal. 2014, 7, re8. [Google Scholar] [CrossRef] [Green Version]
- Sui, H.; Zhu, L.; Deng, W.; Li, Q. Epithelial-mesenchymal transition and drug resistance: Role, molecular mechanisms, and therapeutic strategies. Oncol. Res. Treat. 2014, 37, 584–589. [Google Scholar] [CrossRef]
- Xuan, X.; Tian, C.; Zhao, M.; Sun, Y.; Huang, C. Mesenchymal stem cells in cancer progression and anticancer therapeutic resistance. Cancer Cell Int. 2021, 21, 595. [Google Scholar] [CrossRef] [PubMed]
- Kothari, A.N.; Arffa, M.L.; Chang, V.; Blackwell, R.H.; Syn, W.K.; Zhang, J.; Mi, Z.; Kuo, P.C. Osteopontin—A Master Regulator of Epithelial-Mesenchymal Transition. J. Clin. Med. 2016, 5, 39. [Google Scholar] [CrossRef]
- Das, S.; Samant, R.S.; Shevde, L.A. Nonclassical activation of Hedgehog signaling enhances multidrug resistance and makes cancer cells refractory to Smoothened-targeting Hedgehog inhibition. J. Biol. Chem. 2013, 288, 11824–11833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wang, R.; Wang, M.; Luo, J.; Liu, C. Inhibition of osteopontin overcomes acquired resistance to afatinib in EGFR-mutant non-small-cell lung cancer. Transl. Cancer Res. 2020, 9, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Icard, P.; Shulman, S.; Farhat, D.; Steyaert, J.M.; Alifano, M.; Lincet, H. How the Warburg effect supports aggressiveness and drug resistance of cancer cells? Drug Resist. Updates 2018, 38, 1–11. [Google Scholar] [CrossRef]
- Bonuccelli, G.; Whitaker-Menezes, D.; Castello-Cros, R.; Pavlides, S.; Pestell, R.G.; Fatatis, A.; Witkiewicz, A.K.; Vander Heiden, M.G.; Migneco, G.; Chiavarina, B.; et al. The reverse Warburg effect: Glycolysis inhibitors prevent the tumor promoting effects of caveolin-1 deficient cancer associated fibroblasts. Cell Cycle 2010, 9, 1960–1971. [Google Scholar] [CrossRef] [Green Version]
- Amin, S.; Yang, P.; Li, Z. Pyruvate kinase M2: A multifarious enzyme in non-canonical localization to promote cancer progression. Biochim. Biophys. Acta. Rev. Cancer 2019, 1871, 331–341. [Google Scholar] [CrossRef]
- Ganapathy-Kanniappan, S.; Vali, M.; Kunjithapatham, R.; Buijs, M.; Syed, L.H.; Rao, P.P.; Ota, S.; Kwak, B.K.; Loffroy, R.; Geschwind, J.F. 3-bromopyruvate: A new targeted antiglycolytic agent and a promise for cancer therapy. Curr. Pharm. Biotechnol. 2010, 11, 510–517. [Google Scholar] [CrossRef]
- Nakano, A.; Tsuji, D.; Miki, H.; Cui, Q.; El Sayed, S.M.; Ikegame, A.; Oda, A.; Amou, H.; Nakamura, S.; Harada, T.; et al. Glycolysis inhibition inactivates ABC transporters to restore drug sensitivity in malignant cells. PLoS ONE 2011, 6, e27222. [Google Scholar] [CrossRef]
- Lu, C.; Fang, S.; Weng, Q.; Lv, X.; Meng, M.; Zhu, J.; Zheng, L.; Hu, Y.; Gao, Y.; Wu, X.; et al. Integrated analysis reveals critical glycolytic regulators in hepatocellular carcinoma. Cell Commun. Signal. 2020, 18, 97. [Google Scholar] [CrossRef]
- Bellahcene, A.; Castronovo, V.; Ogbureke, K.U.; Fisher, L.W.; Fedarko, N.S. Small integrin-binding ligand N-linked glycoproteins (SIBLINGs): Multifunctional proteins in cancer. Nat. Rev. Cancer 2008, 8, 212–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Luo, M.; Hu, J.; Chen, C.; Mei, H. Osteopontin enhances cisplatin resistance of human A549 lung cancer cells via stimulating the PI3K signaling pathway and upregulating ERCC1 expression. Transl. Cancer Res. 2020, 9, 3258–3265. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.; Fan, L.; Chen, S.; Wang, Y.; Yu, H.; Sun, Y.; Yu, J.; Wang, L.; Liu, X.; Liu, Y. Overexpression of osteopontin promotes resistance to cisplatin treatment in HCC. Oncol. Rep. 2015, 34, 3297–3303. [Google Scholar] [CrossRef] [Green Version]
- Zahed Panah, M.; Nikbakht, M.; Sajjadi, S.M.; Rostami, S.; Norooznezhad, A.H.; Kamranzadeh Fumani, H.; Ghavamzadeh, A.; Mohammadi, S. Anti-Apoptotic Effects of Osteopontin via the Up-Regulation of AKT/mTOR/beta-Catenin Loop in Acute Myeloid Leukemia Cells. Int. J. Hematol. Oncol. Stem Cell Res. 2017, 11, 148–157. [Google Scholar] [PubMed]
- Yi, H.; Zeng, D.; Shen, Z.; Liao, J.; Wang, X.; Liu, Y.; Zhang, X.; Kong, P. Integrin alphavbeta3 enhances beta-catenin signaling in acute myeloid leukemia harboring Fms-like tyrosine kinase-3 internal tandem duplication mutations: Implications for microenvironment influence on sorafenib sensitivity. Oncotarget 2016, 7, 40387–40397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Wang, J.; Lin, C.; Liu, J.; Zuo, W. Osteopontin Mediates Cetuximab Resistance via the MAPK Pathway in NSCLC Cells. OncoTargets Ther. 2019, 12, 10177–10185. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Wang, J.; Ma, S.; Huang, Z.; Zhang, G. Requirement of Osteopontin in the migration and protection against Taxol-induced apoptosis via the ATX-LPA axis in SGC7901 cells. BMC Cell Biol. 2011, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Sigismund, S.; Avanzato, D.; Lanzetti, L. Emerging functions of the EGFR in cancer. Mol. Oncol. 2018, 12, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Li, L.; Puliyappadamba, V.T.; Guo, G.; Hatanpaa, K.J.; Mickey, B.; Souza, R.F.; Vo, P.; Herz, J.; Chen, M.R.; et al. Constitutive and ligand-induced EGFR signalling triggers distinct and mutually exclusive downstream signalling networks. Nat. Commun. 2014, 5, 5811. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.D.; Ma, L.; Wong, M.P.; Lee, V.H.; Yan, H. Contribution of EGFR and ErbB-3 Heterodimerization to the EGFR Mutation-Induced Gefitinib- and Erlotinib-Resistance in Non-Small-Cell Lung Carcinoma Treatments. PLoS ONE 2015, 10, e0128360. [Google Scholar] [CrossRef] [Green Version]
- Baldacci, S.; Mazieres, J.; Tomasini, P.; Girard, N.; Guisier, F.; Audigier-Valette, C.; Monnet, I.; Wislez, M.; Perol, M.; Do, P.; et al. Outcome of EGFR-mutated NSCLC patients with MET-driven resistance to EGFR tyrosine kinase inhibitors. Oncotarget 2017, 8, 105103–105114. [Google Scholar] [CrossRef] [PubMed]
- Lamour, V.; Henry, A.; Kroonen, J.; Nokin, M.J.; von Marschall, Z.; Fisher, L.W.; Chau, T.L.; Chariot, A.; Sanson, M.; Delattre, J.Y.; et al. Targeting osteopontin suppresses glioblastoma stem-like cell character and tumorigenicity in vivo. Int. J. Cancer 2015, 137, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhang, Y.; Lei, Z.; Liu, T.; Cai, T.; Wang, A.; Du, W.; Zeng, Y.; Zhu, J.; Liu, Z.; et al. Abnormally activated OPN/integrin alphaVbeta3/FAK signalling is responsible for EGFR-TKI resistance in EGFR mutant non-small-cell lung cancer. J. Hematol. Oncol. 2020, 13, 169. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.; Li, P.; Yan, W.; Shi, L.; Zhang, J.; Wang, Y.; Liu, H.; You, Y. Downregulation of osteopontin enhances the sensitivity of glioma U251 cells to temozolomide and cisplatin by targeting the NF-κB/Bcl2 pathway. Mol. Med. Rep. 2015, 11, 1951–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.W.; Kim, M.S.; Voon, D.C.; Kim, S.J.; Bae, J.; Mun, D.G.; Ko, S.I.; Kim, H.K.; Lee, S.W.; Kim, D.Y. Multi-omics analysis identifies pathways and genes involved in diffuse-type gastric carcinogenesis induced by E-cadherin, p53, and Smad4 loss in mice. Mol. Carcinog. 2018, 57, 947–954. [Google Scholar] [CrossRef]
- Boidot, R.; Vegran, F.; Lizard-Nacol, S. Predictive value of survivin alternative transcript expression in locally advanced breast cancer patients treated with neoadjuvant chemotherapy. Int. J. Mol. Med. 2009, 23, 285–291. [Google Scholar]
- Thadani-Mulero, M.; Portella, L.; Sun, S.; Sung, M.; Matov, A.; Vessella, R.L.; Corey, E.; Nanus, D.M.; Plymate, S.R.; Giannakakou, P. Androgen receptor splice variants determine taxane sensitivity in prostate cancer. Cancer Res. 2014, 74, 2270–2282. [Google Scholar] [CrossRef] [Green Version]
- Marima, R.; Francies, F.Z.; Hull, R.; Molefi, T.; Oyomno, M.; Khanyile, R.; Mbatha, S.; Mabongo, M.; Owen Bates, D.; Dlamini, Z. MicroRNA and Alternative mRNA Splicing Events in Cancer Drug Response/Resistance: Potent Therapeutic Targets. Biomedicines 2021, 9, 1818. [Google Scholar] [CrossRef]
- Gimba, E.R.; Tilli, T.M. Human osteopontin splicing isoforms: Known roles, potential clinical applications and activated signaling pathways. Cancer Lett. 2013, 331, 11–17. [Google Scholar] [CrossRef]
- Mirzaei, A.; Mohammadi, S.; Ghaffari, S.H.; Nikbakht, M.; Bashash, D.; Alimoghaddam, K.; Ghavamzadeh, A. Osteopontin b and c isoforms: Molecular Candidates Associated with Leukemic Stem Cell Chemoresistance in Acute Myeloid Leukemia. Asian Pac. J. Cancer Prev. 2017, 18, 1707–1715. [Google Scholar] [CrossRef]
- Mirzaei, A.; Ghaffari, S.H.; Nikbakht, M.; Kamranzadeh Foumani, H.; Vaezi, M.; Mohammadi, S.; Alimoghaddam, K.; Ghavamzadeh, A. OPN b and c Isoforms Doubtless Veto Anti-angiogenesis Effects of Curcumin in Combination with Conventional AML Regiment. Asian Pac. J. Cancer Prev. 2017, 18, 2591–2599. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.D.; Tilli, T.M.; Wanderley, J.L.; Palumbo, A., Jr.; Mattos, R.M.; Ferreira, A.C.; Klumb, C.E.; Nasciutti, L.E.; Gimba, E.R. Osteopontin splice variants expression is involved on docetaxel resistance in PC3 prostate cancer cells. Tumor Biol. 2016, 37, 2655–2663. [Google Scholar] [CrossRef] [PubMed]
- Brum, M.C.M.; Dos Santos Guimaraes, I.; Ferreira, L.B.; Rangel, L.B.A.; Maia, R.C.; Nestal De Moraes, G.; Gimba, E.R.P. Osteopontinc isoform inhibition modulates ovarian cancer cell cisplatin resistance, viability and plasticity. Oncol. Rep. 2021, 45, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Huang, J.; Niu, H.; Wang, J.; Si, Y.; Bai, Z.; Cheng, S.; Ding, W. Epigenetic regulation of osteopontin splicing isoform c defines its role as a microenvironmental factor to promote the survival of colon cancer cells from 5-FU treatment. Cancer Cell Int. 2020, 20, 452. [Google Scholar] [CrossRef] [PubMed]
- Yosifov, D.Y.; Reufsteck, C.; Konstantinov, S.M.; Berger, M.R. Interleukin-6, osteopontin and Raf/MEK/ERK signaling modulate the sensitivity of human myeloma cells to alkylphosphocholines. Leuk. Res. 2012, 36, 764–772. [Google Scholar] [CrossRef]
- Christensen, B.; Kazanecki, C.C.; Petersen, T.E.; Rittling, S.R.; Denhardt, D.T.; Sorensen, E.S. Cell type-specific post-translational modifications of mouse osteopontin are associated with different adhesive properties. J. Biol. Chem. 2007, 282, 19463–19472. [Google Scholar] [CrossRef] [Green Version]
- Gothlin Eremo, A.; Lagergren, K.; Othman, L.; Montgomery, S.; Andersson, G.; Tina, E. Evaluation of SPP1/osteopontin expression as predictor of recurrence in tamoxifen treated breast cancer. Sci. Rep. 2020, 10, 1451. [Google Scholar] [CrossRef] [Green Version]
- Faria, M.; Halquist, M.S.; Yuan, M.; Mylott, W., Jr.; Jenkins, R.G.; Karnes, H.T. Comparison of a stable isotope labeled (SIL) peptide and an extended SIL peptide as internal standards to track digestion variability of an unstable signature peptide during quantification of a cancer biomarker, human osteopontin, from plasma using capillary microflow LC-MS/MS. J. Chromatogr. B 2015, 1001, 156–168. [Google Scholar] [CrossRef]
- Macur, K.; Hagen, L.; Ciesielski, T.M.; Konieczna, L.; Skokowski, J.; Jenssen, B.M.; Slupphaug, G.; Baczek, T. A targeted mass spectrometry immunoassay to quantify osteopontin in fresh-frozen breast tumors and adjacent normal breast tissues. J. Proteom. 2019, 208, 103469. [Google Scholar] [CrossRef]
- Meirinho, S.G.; Dias, L.G.; Peres, A.M.; Rodrigues, L.R. Development of an electrochemical RNA-aptasensor to detect human osteopontin. Biosens. Bioelectron. 2015, 71, 332–341. [Google Scholar] [CrossRef] [Green Version]
- Zagani, R.; Hamzaoui, N.; Cacheux, W.; de Reynies, A.; Terris, B.; Chaussade, S.; Romagnolo, B.; Perret, C.; Lamarque, D. Cyclooxygenase-2 inhibitors down-regulate osteopontin and Nr4A2-new therapeutic targets for colorectal cancers. Gastroenterology 2009, 137, 1358–1366.e3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yamada, O.; Kida, S.; Matsushita, Y.; Murase, S.; Hattori, T.; Kubohara, Y.; Kikuchi, H.; Oshima, Y. Identification of brefelamide as a novel inhibitor of osteopontin that suppresses invasion of A549 lung cancer cells. Oncol. Rep. 2016, 36, 2357–2364. [Google Scholar] [CrossRef] [PubMed]


| Cancer Type | Preclinical Models | Biological Function | Reference |
|---|---|---|---|
| Lung cancer | In vitro | Exogenous beclin1-induced autophagy abrogated radioresistance of lung cancer cells by suppressing OPN. | [35] |
| In vitro | OPN over-expression induced the expressions of EMT biomarkers, and promoted acquired resistance to afatinib in H1650-AR and H1975-AR cells. | [64] | |
| In vitro | OPN promoted cancer cell lactate and LDHA production and able to predict the response of A549 and SK-MES-1 to first-line platinum-based chemotherapy. | [14] | |
| In vitro | OPN enhanced cisplatin resistance of A549 lung cancer cells via stimulating the PI3K signalling pathway and upregulating ERCC1 expression | [72] | |
| In vitro | Overexpression of OPN reduced the sensitivity of NSCLC cells to cetuximab by upregulating MAPK pathway-related proteins. | [76] | |
| In vitro | OPN was overexpressed in acquired EGFR-TKI-resistant NSCLCs. OPN contributed to acquired gefitinib resistance by activating the integrin αVβ3/FAK pathway. | [83] | |
| Breast cancer | In vivo | The OPN-targeting miRNA, hsa-mir-299–5p was commonly downregulated in the SFC subpopulations from three breast cancer cell lines, and the elevated OPN highlighted their role in tumour angiogenesis. | [53] |
| In vitro | MiR-181c may regulate chemosensitivity by downregulating OPN, resulting in enhanced p53-dependent transactivation and apoptosis in resistant MCF-7 cells. | [54] | |
| In vitro | OPN enhanced EMT phenotype and the expression of ABCB1 and ABCG2 proteins in a GLI-dependent manner. Silencing OPN improved the susceptibility of breast cancer cells to DOX, paclitaxel and cisplatin. | [63] | |
| Hepatocellular carcinoma (HCC) | In vitro | OPN expression was elevated during starvation-induced autophagy in HCCs. OPN engaged with integrin αvβ3 and sustained the stability of FoxO3a to induce autophagy, which further promoted stem-like phenotype of HCCs and resistance to epirubicin and cisplatin. | [33] |
| In vitro and in vivo | OPN was closely related to the sensitivity of CD133+/CD44+ subgroup of HCC cells to 5 Aza. | [49] | |
| In vitro | OPN enhanced chemoresistance of cisplatin in HCC cells by activating PI3K/AKT signalling pathway. | [73] | |
| In vivo | OPN enhanced HCC glycolysis by activating the αvβ3-NF-κB signalling and hepatocarcinogenesis induced by DEN. | [70] | |
| Gastric cancer | In vitro | Expression of OPN was mediated by the activation of Akt and MAPK/ERK pathways through the LPA2 receptor and protected SGC7901 cells from apoptosis induced by Taxol treatment. | [77] |
| In vivo | OPN contributes to stemness of diffuse-type gastric cancer cells and chemoresistance by pro-moting tumour sphere formation and induction of Bcl-xL. | [85] | |
| Colon cancer | In vitro | OPN-c could transmit the stress signal of cells upon 5-FU treatment in tumor microenvironment and promoted the survival of adjacent colon cancer cells. | [94] |
| Pancreatic cancer | In vitro | OPN/NF-κB signalling upregulated pancreatic CSC activity by activating autophagy. | [34] |
| In vivo | Autophagy blockade sensitizes pancreatic CSCs to gemcitabine. | ||
| Prostate cancer | In vitro | OPN upregulated P-gp expression through integrin αvβ3 in PC-3 cancer cells and inhibited DUN-Induced cell death. Knockdown of endogenous OPN enhanced the cytotoxicity of paclitaxel, doxorubicin, actinomycin-D, and rapamycin. | [42] |
| In vivo | Knockdown of OPN enhanced the cytotoxicity of DUN in xenograft animal model. | ||
| In vitro | OPN-c or OPN-b overexpression in PC3 cells could mediate resistance and cell survival features in response to DXT-induced cell death. | [92] | |
| Glioblastoma | In vivo | Endogenous OPN-mediated EGFR activation promoted GICs stemness phenotype and tumorigenicity. | [82] |
| In vitro | OPN downregulation in U251 cells enhanced the effects of TMZ and DDP chemotherapy by targeting the NF-κB/Bcl-2 pathway. | [84] | |
| Cervical cancer | In vitro | Overexpression of miR-181a can inhibit the expression of OPN and reduce DDP resistance in CaSki cells. | [55] |
| Ovarian cancer | In vitro | OPN-c was upregulated in cisplatin -resistant ACRP cells. Cells with OPN-c positive expression were more sensitive to cisplatin cytotoxicity compared with the negative control cells. | [93] |
| Melanoma | In vivo | OPN enhanced ABCG2 expression and enriched SP phenotype through ERK signalling in murine melanoma cells, and SP cells had high efflux capacity for mitoxantrone. | [43] |
| Acute myeloid leukemia (AML) | In vitro | Resistance of CD34+ AML cells to DNR might be relevant to increasing of OPN mRNA expression and activity of other mediators including AKT, mTOR, PTEN, and β-catenin. | [74] |
| In vitro | Integrin αvβ3/ PI3K/ Akt/ GSK3β/ β-catenin/OPN axis was crucial for microenvironment mediated sorafenib insensitivity in FLT3ITD cells. | [75] | |
| In vitro | OPN isoforms, particularly b and c, alongside with VEGF isoforms and other gene pathways might promote chemoresistance in leukaemia. | [91] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, C.; Lane, J.; Jiang, W.G. Osteopontin and Cancer: Insights into Its Role in Drug Resistance. Biomedicines 2023, 11, 197. https://doi.org/10.3390/biomedicines11010197
Hao C, Lane J, Jiang WG. Osteopontin and Cancer: Insights into Its Role in Drug Resistance. Biomedicines. 2023; 11(1):197. https://doi.org/10.3390/biomedicines11010197
Chicago/Turabian StyleHao, Chengcheng, Jane Lane, and Wen G. Jiang. 2023. "Osteopontin and Cancer: Insights into Its Role in Drug Resistance" Biomedicines 11, no. 1: 197. https://doi.org/10.3390/biomedicines11010197