
The Inflammatory Gene PYCARD of the Entorhinal Cortex as an Early Diagnostic Target for Alzheimer’s Disease


Abstract
:1. Introduction
2. Methods
2.1. Data Download and Preprocessing
2.2. Identification of Differentially Expressed Genes (DEGs) for AD
2.3. Functional Enrichment Analysis of DEGs
2.4. Verification of the DEGs
3. Results
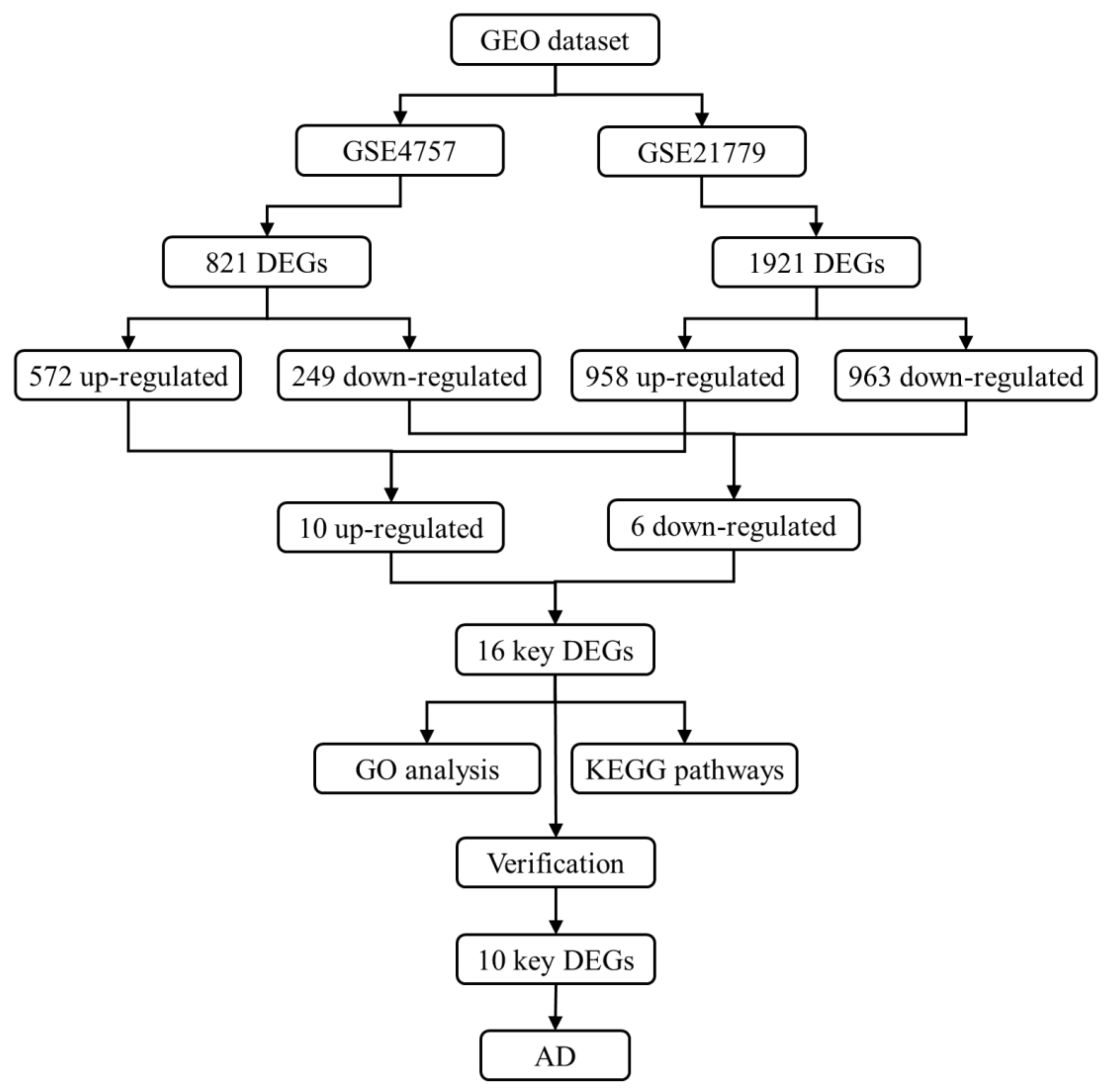
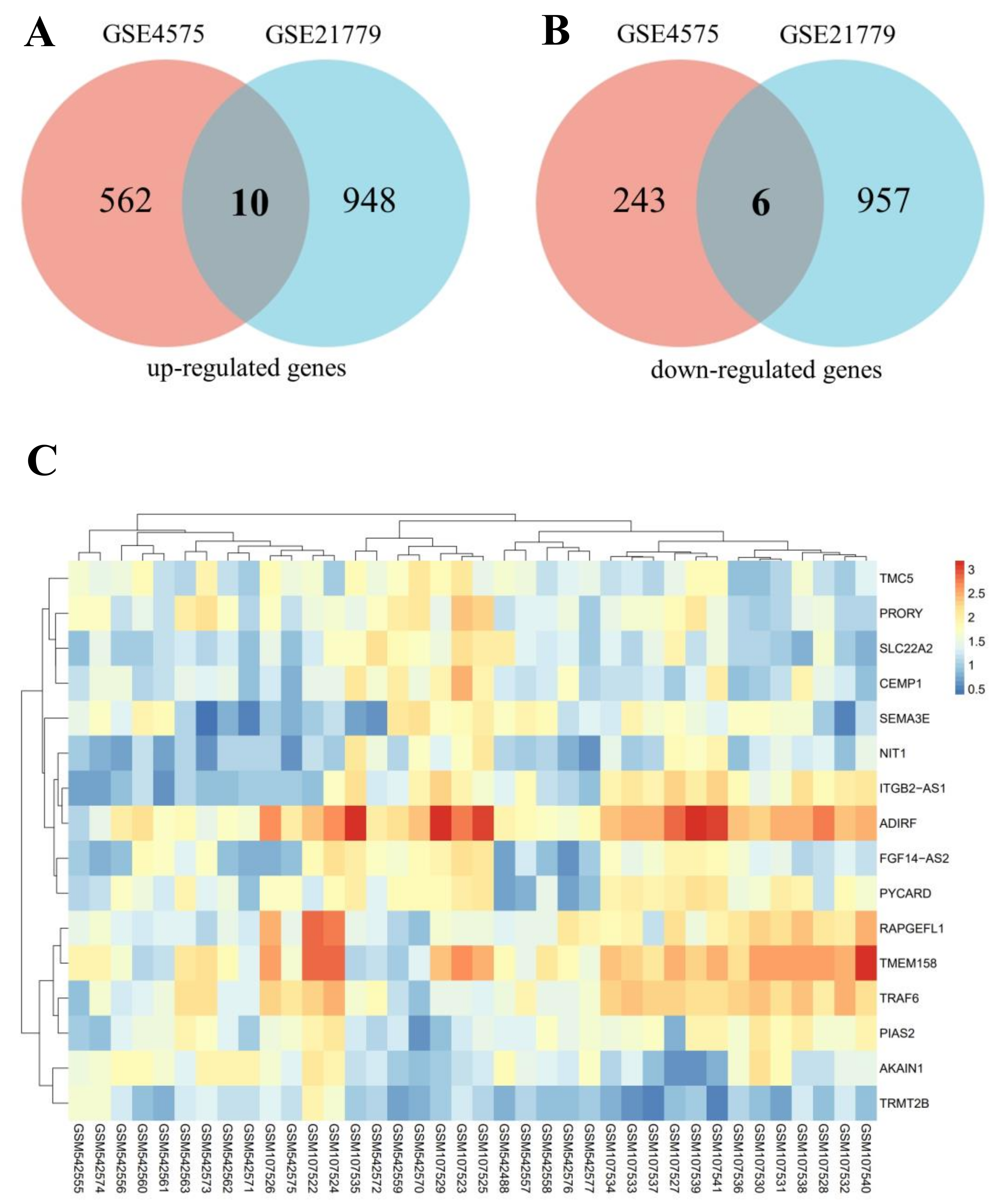
3.1. Screening of DEGs
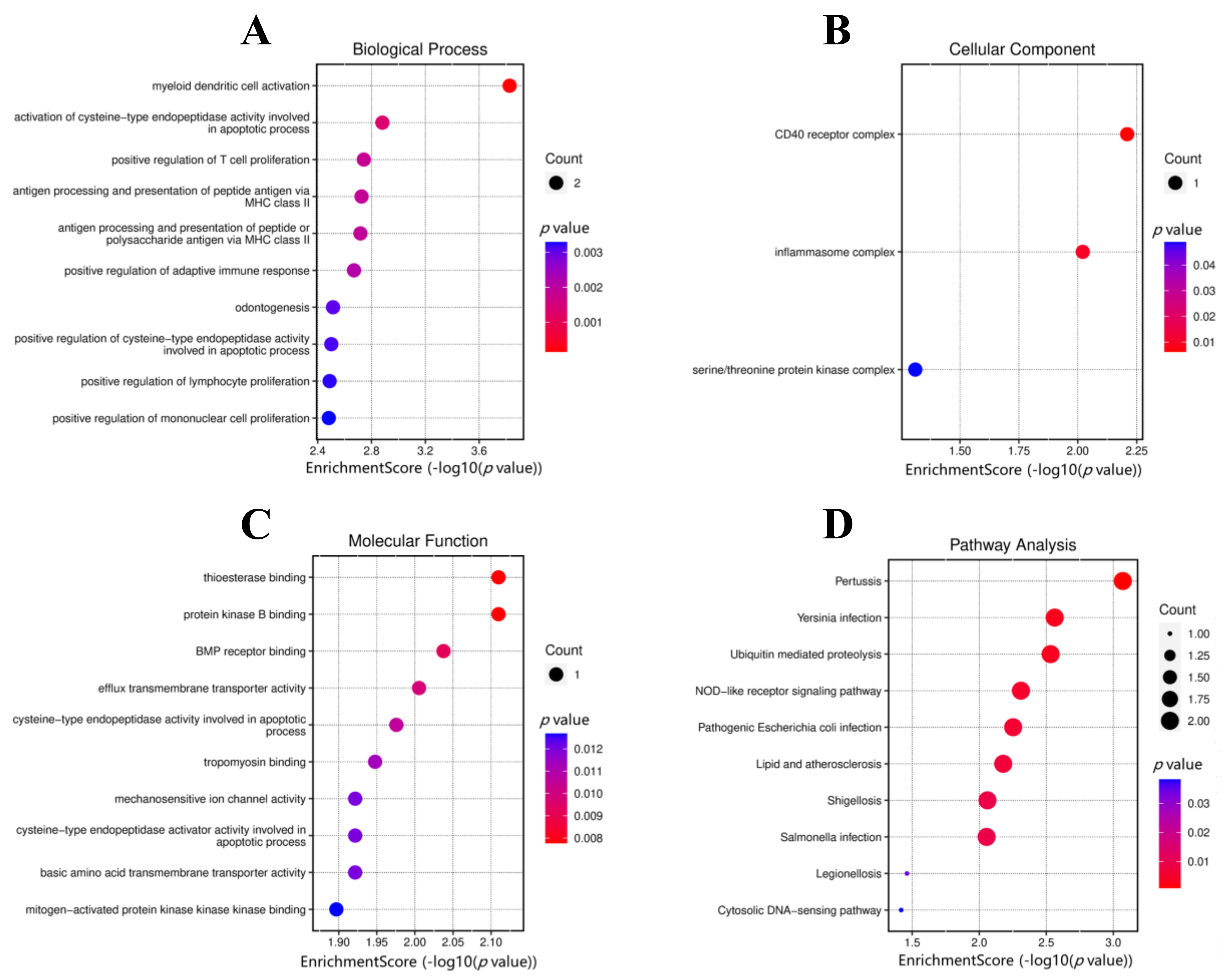
3.2. DEGs Functional Enrichment Analysis
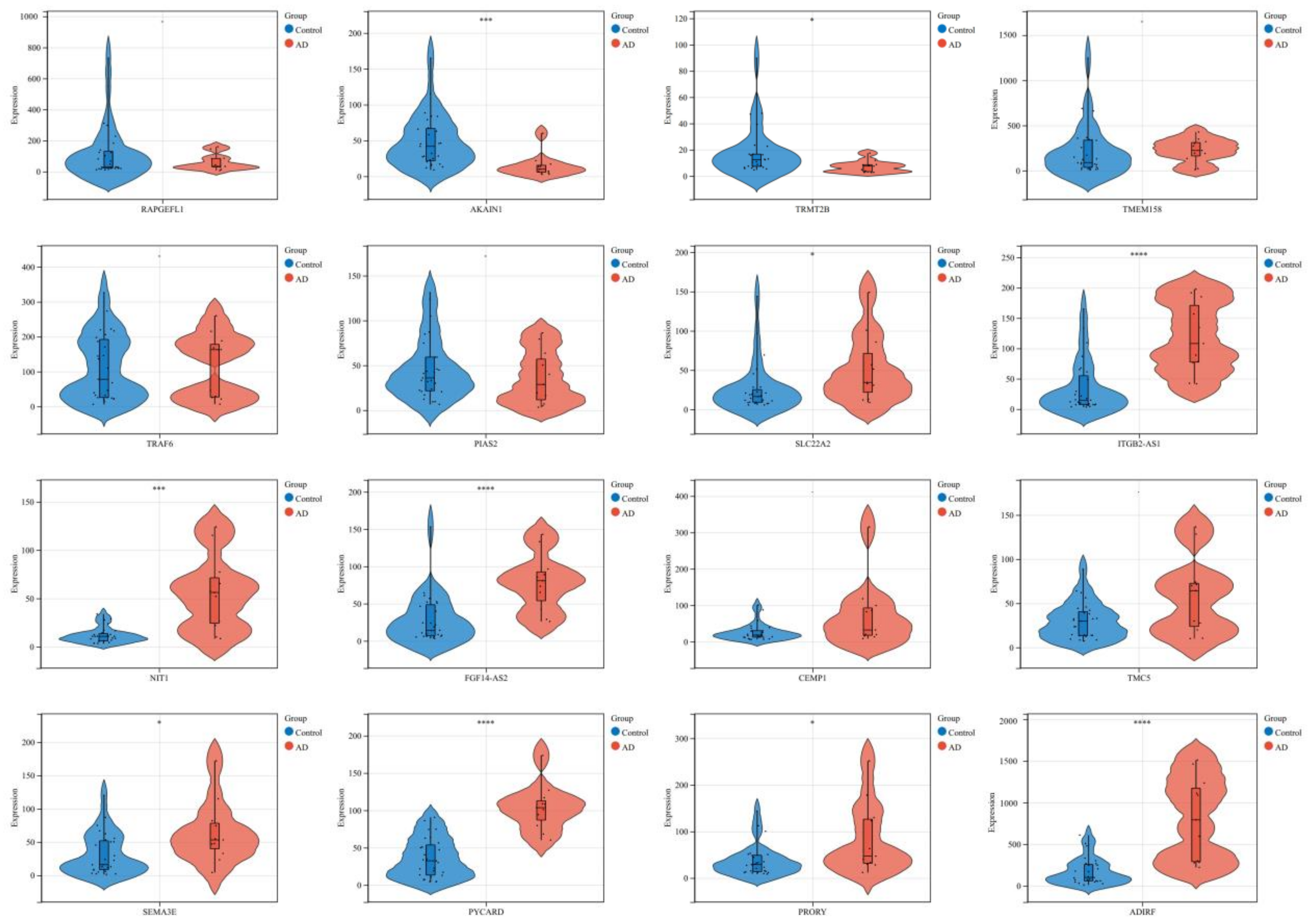
3.3. Significant and Highly Valued DEGs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Armstrong, R.A. What causes Alzheimer’s disease? Folia Neuropathol. 2013, 51, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Crous-Bou, M.; Minguillón, C.; Gramunt, N.; Molinuevo, J.L. Alzheimer’s disease prevention: From risk factors to early intervention. Alzheimer’s Res. 2017, 9, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Soria Lopez, J.A.; González, H.H.; Léger, G.C. Alzheimer’s disease. Handb. Clin. Neurol. 2019, 167, 231–255. [Google Scholar] [CrossRef] [PubMed]
- Suh, Y.Y.; Checler, F. Amyloid precursor protein, presenilins, and alpha-synuclein: Molecular pathogenesis and pharmacological applications in Alzheimer’s disease. Pharm. Rev. 2002, 54, 469–525. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Gu, B.B.; Masters, C.C.; Wang, Y.J. A systemic view of Alzheimer disease-insights from amyloid-β metabolism beyond the brain. Nat. Rev. Neurol. 2017, 13, 703. [Google Scholar] [CrossRef] [Green Version]
- Khan, U.U.; Liu, L.; Provenzano, F.F.; Berman, D.D.; Profaci, C.C.; Sloan, R.; Mayeux, R.; Duff, K.E.; Small, S.A. Molecular drivers and cortical spread of lateral entorhinal cortex dysfunction in preclinical Alzheimer’s disease. Nat. Neurosci. 2014, 17, 304–311. [Google Scholar] [CrossRef] [Green Version]
- Whitwell, J.J.; Przybelski, S.S.; Weigand, S.S.; Knopman, D.D.; Boeve, B.B.; Petersen, R.C.; Jack, C.R. 3D maps from multiple MRI illustrate changing atrophy patterns as subjects progress from mild cognitive impairment to Alzheimer’s disease. Brain 2007, 130, 1777–1786. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, F.; Zhao, L.; Qian, J.; Dong, C. Entorhinal cortex: A good biomarker of mild cognitive impairment and mild Alzheimer’s disease. Rev. Neurosci. 2016, 27, 185–195. [Google Scholar] [CrossRef]
- Holbrook, A.A.; Tustison, N.N.; Marquez, F.; Roberts, J.; Yassa, M.M.; Gillen, D.L.; Alzheimer’s Disease Neuroimaging Initiative. Anterolateral entorhinal cortex thickness as a new biomarker for early detection of Alzheimer’s disease. Alzheimer’s Dement. 2020, 12, e12068. [Google Scholar] [CrossRef]
- Leandrou, S.; Lamnisos, D.; Mamais, I.; Kyriacou, P.P.; Pattichis, C.S.; Alzheimer’s Disease Neuroimaging Initiative. Assessment of Alzheimer’s Disease Based on Texture Analysis of the Entorhinal Cortex. Front. Aging Neurosci. 2020, 12, 176. [Google Scholar] [CrossRef]
- Olajide, O.O.; Suvanto, M.M.; Chapman, C.A. Molecular mechanisms of neurodegeneration in the entorhinal cortex that underlie its selective vulnerability during the pathogenesis of Alzheimer’s disease. Biol. Open 2021, 10, bio056796. [Google Scholar] [CrossRef]
- Cameron, B.; Landreth, G.E. Inflammation, microglia, and Alzheimer’s disease. Neurobiol. Dis. 2010, 37, 503–509. [Google Scholar] [CrossRef] [Green Version]
- Markiewicz, I.; Lukomska, B. The role of astrocytes in the physiology and pathology of the central nervous system. Acta Neurobiol. Exp. 2006, 66, 343–358. [Google Scholar]
- Matias, I.; Morgado, J.; Gomes, F.C.A. Astrocyte Heterogeneity: Impact to Brain Aging and Disease. Front. Aging Neurosci. 2019, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Ransom, B.B.; Ransom, C.B. Astrocytes: Multitalented stars of the central nervous system. In Astrocytes; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; Volume 814, pp. 3–7. [Google Scholar] [CrossRef]
- Zhang, D.; Hu, X.; Qian, L.; O’Callaghan, J.J.; Hong, J.S. Astrogliosis in CNS pathologies: Is there a role for microglia? Mol. Neurobiol. 2010, 41, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Simon, E.; Obst, J.; Gomez-Nicola, D. The Evolving Dialogue of Microglia and Neurons in Alzheimer’s Disease: Microglia as Necessary Transducers of Pathology. Neuroscience 2019, 405, 24–34. [Google Scholar] [CrossRef]
- Asai, H.; Ikezu, S.; Tsunoda, S.; Medalla, M.; Luebke, J.; Haydar, T.; Wolozin, B.; Butovsky, O.; Kügler, S.; Ikezu, T. Depletion of microglia and inhibition of exosome synthesis halt tau propagation. Nat. Neurosci. 2015, 18, 1584–1593. [Google Scholar] [CrossRef] [Green Version]
- Criscuolo, C.; Fontebasso, V.; Middei, S.; Stazi, M.; Ammassari-Teule, M.; Yan, S.S.; Origlia, N. Entorhinal Cortex dysfunction can be rescued by inhibition of microglial RAGE in an Alzheimer’s disease mouse model. Sci. Rep. 2017, 7, 42370. [Google Scholar] [CrossRef] [Green Version]
- Janelsins, M.M.; Mastrangelo, M.M.; Oddo, S.; LaFerla, F.F.; Federoff, H.H.; Bowers, W.J. Early correlation of microglial activation with enhanced tumor necrosis factor-alpha and monocyte chemoattractant protein-1 expression specifically within the entorhinal cortex of triple transgenic Alzheimer’s disease mice. J. Neuroinflamm. 2005, 2, 23. [Google Scholar] [CrossRef]
- Thangavel, R.; Kempuraj, D.; Stolmeier, D.; Anantharam, P.; Khan, M.; Zaheer, A. Glia maturation factor expression in entorhinal cortex of Alzheimer’s disease brain. Neurochem. Res. 2013, 38, 1777–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, C.C.; Vadhwana, B.; Verkhratsky, A.; Rodríguez, J.J. Early astrocytic atrophy in the entorhinal cortex of a triple transgenic animal model of Alzheimer’s disease. ASN Neuro 2011, 3, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Brooks, D.D.; Okello, A.; Edison, P. An early and late peak in microglial activation in Alzheimer’s disease trajectory. Brain 2017, 140, 792–803. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, J.; Jiang, W.; Cao, Z.; Zhao, F.; Cai, T.; Aschner, M.; Luo, W. The role of NLRP3-CASP1 in inflammasome-mediated neuroinflammation and autophagy dysfunction in manganese-induced, hippocampal-dependent impairment of learning and memory ability. Autophagy 2017, 13, 914–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolezzi, J.J.; Santos, M.M.; Bastias-Candia, S.; Pinto, C.; Godoy, J.J.; Inestrosa, N.C. PPARs in the central nervous system: Roles in neurodegeneration and neuroinflammation. Biol. Rev. Camb. Philos. Soc. 2017, 92, 2046–2069. [Google Scholar] [CrossRef]
- Feng, B.; Hu, P.; Chen, J.; Liu, Q.; Li, X.; Du, Y. Analysis of Differentially Expressed Genes Associated with Alzheimer’s Disease Based on Bioinformatics Methods. Am. J. Alzheimer’s Dis. Other Dement. 2015, 30, 746–751. [Google Scholar] [CrossRef]
- Wu, Y.; Liang, S.; Zhu, H.; Zhu, Y. Analysis of immune-related key genes in Alzheimer’s disease. Bioengineered 2021, 12, 9610–9624. [Google Scholar] [CrossRef]
- Qiu, H.; Weng, Q. Screening of Crucial Differentially-Methylated/Expressed Genes for Alzheimer’s Disease. Am. J. Alzheimer’s Dis. Other Dement. 2022, 37, 15333175221116220. [Google Scholar] [CrossRef]
- Shippy, D.D.; Watters, J.J.; Ulland, T.K. Transcriptional response of murine microglia in Alzheimer’s disease and inflammation. BMC Genom. 2022, 23, 183. [Google Scholar] [CrossRef]
- Liang, W.W.; Dunckley, T.; Beach, T.T.; Grover, A.; Mastroeni, D.; Walker, D.G.; Caselli, R.J.; Kukull, W.A.; McKeel, D.; Morris, J.C.; et al. Gene expression profiles in anatomically and functionally distinct regions of the normal aged human brain. Physiol. Genom. 2007, 28, 311–322. [Google Scholar] [CrossRef]
- Bottero, V.; Powers, D.; Yalamanchi, A.; Quinn, J.J.; Potashkin, J.A. Key Disease Mechanisms Linked to Alzheimer’s Disease in the Entorhinal Cortex. Int. J. Mol. Sci. 2021, 22, 3915. [Google Scholar] [CrossRef]
- Dunckley, T.; Beach, T.T.; Ramsey, K.E.; Grover, A.; Mastroeni, D.; Walker, D.G.; LaFleur, B.J.; Coon, K.D.; Brown, K.M.; Caselli, R.; et al. Gene expression correlates of neurofibrillary tangles in Alzheimer’s disease. Neurobiol. Aging 2006, 27, 1359–1371. [Google Scholar] [CrossRef] [Green Version]
- Abdel Rassoul, R.; Alves, S.; Pantesco, V.; De Vos, J.; Michel, B.; Perret, M.; Mestre-Francés, N.; Verdier, J.-M.; Devau, G. Distinct transcriptome expression of the temporal cortex of the primate Microcebus murinus during brain aging versus Alzheimer’s disease-like pathology. PLoS ONE 2010, 5, e12770. [Google Scholar] [CrossRef]
- Ritchie, M.M.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.L.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Zhe, W.; Ryan, M. TModel-free posterior inference on the area under the receiver operating characteristic curve. J. Stat. Plan. Inference 2020, 209, 174–186. [Google Scholar]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Jiang, Y.; Xu, K.; Cui, M.; Ye, W.; Zhao, G.; Jin, L.; Chen, X. The progress of gut microbiome research related to brain disorders. J. Neuroinflamm. 2020, 17, 25. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, K.; Mulak, A. Brain-Gut-Microbiota Axis in Alzheimer’s Disease. J. Neurogastroenterol. Motil. 2019, 25, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Roy Sarkar, S.; Banerjee, S. Gut microbiota in neurodegenerative disorders. J. Neuroimmunol. 2019, 328, 98–104. [Google Scholar] [CrossRef]
- Borrego, A.; Colombo, F.; de Souza, J.G.; Jensen, J.R.; Dassano, A.; Piazza, R.; dos Santos, B.A.R.; Ribeiro, O.G.; De Franco, M.; Cabrera, W.H.K.; et al. Pycard and BC017158 Candidate Genes of Irm1 Locus Modulate Inflammasome Activation for IL-1β Production. Front. Immunol. 2022, 13, 899569. [Google Scholar] [CrossRef] [PubMed]
- Sborgi, L.; Ravotti, F.; Dandey, V.V.; Dick, M.M.; Mazur, A.; Reckel, S.; Chami, M.; Scherer, S.; Huber, M.; Böckmann, A.; et al. Structure and assembly of the mouse ASC inflammasome by combined NMR spectroscopy and cryo-electron microscopy. Proc. Natl. Acad. Sci. USA 2015, 112, 13237–13242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, J.J.; Vafeiadou, K.; Williams, R.R.; Vauzour, D. Neuroinflammation: Modulation by flavonoids and mechanisms of action. Mol. Asp. Med. 2012, 33, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Si, Z.Z.; Zou, C.C.; Mei, X.; Li, X.X.; Luo, H.; Shen, Y.; Hu, J.; Li, X.-X.; Wu, L. Targeting neuroinflammation in Alzheimer’s disease: From mechanisms to clinical applications. Neural Regen. Res. 2022, 18, 708–715. [Google Scholar] [CrossRef]
- Hensley, K. Neuroinflammation in Alzheimer’s disease: Mechanisms, pathologic consequences, and potential for therapeutic manipulation. J. Alzheimer’s Dis. 2010, 21, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wyss-Coray, T.; Rogers, J. Inflammation in Alzheimer disease-a brief review of the basic science and clinical literature. Cold Spring Harb. Perspect. Med. 2012, 2, a006346. [Google Scholar] [CrossRef]
- Venegas, C.; Kumar, S.; Franklin, B.S.; Dierkes, T.; Brinkschulte, R.; Tejera, D.; Vieira-Saecker, A.; Schwartz, S.; Santarelli, F.; Kummer, M.P.; et al. Microglia-derived ASC specks cross-seed amyloid-β in Alzheimer’s disease. Nature 2017, 552, 355–361. [Google Scholar] [CrossRef]
- Brown, G.G.; Bal-Price, A. Inflammatory neurodegeneration mediated by nitric oxide, glutamate, and mitochondria. Mol. Neurobiol. 2003, 27, 325–355. [Google Scholar] [CrossRef]
- McGeer, E.E.; McGeer, P.L. Neuroinflammation in Alzheimer’s disease and mild cognitive impairment: A field in its infancy. J. Alzheimer’s Dis. 2010, 19, 355–361. [Google Scholar] [CrossRef]
- Zhang, F.; Jiang, L. Neuroinflammation in Alzheimer’s disease. Neuropsychiatr. Dis. Treat. 2015, 11, 243–256. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
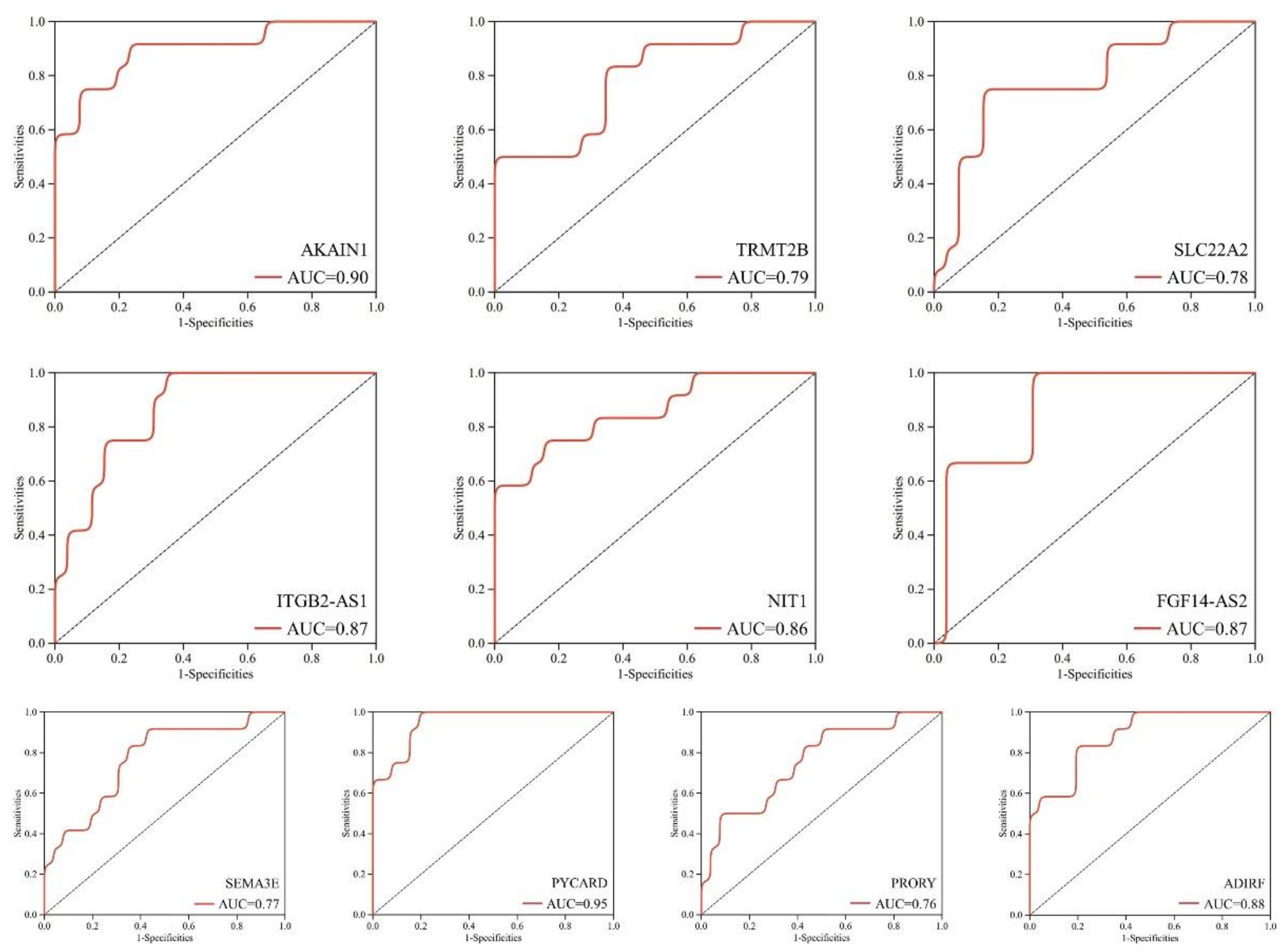{kind=link}
| DEGs | Count | ID | p Value | logFC | Gene | Description |
|---|---|---|---|---|---|---|
| Up_regulated | 10 | 207429_at | 0.042 | 1.884 | SLC22A2 | solute carrier family 22 member 2 |
| 229041_s_at | 0.013 | 2.136 | ITGB2-AS1 | ITGB2 antisense RNA 1 | ||
| 241395_at | 0.022 | 1.739 | NIT1 | nitrilase 1 | ||
| 243225_at | 0.047 | 2.123 | FGF14-AS2 | FGF14 antisense RNA 2 | ||
| 222009_at | 0.039 | 1.559 | CEMP1 | cementum protein 1 | ||
| 219580_s_at | 0.027 | 1.648 | TMC5 | transmembrane channel like 5 | ||
| 206941_x_at | 0.028 | 3.083 | SEMA3E | semaphorin 3E | ||
| 221666_s_at | 0.048 | 1.968 | PYCARD | PYD and CARD domain containing | ||
| 221179_at | 0.026 | 1.979 | PRORY | proline rich, Y-linked | ||
| 203571_s_at | 0.020 | 1.741 | ADIRF | adipogenesis regulatory factor | ||
| Down_regulated | 6 | 218657_at | 0.020 | −1.624 | RAPGEFL1 | Rap guanine nucleotide exchange factor like 1 |
| 243506_at | 0.001 | −2.416 | AKAIN1 | A kinase (PRKA) anchor inhibitor 1 | ||
| 1554279_a_at | 0.048 | −1.434 | TRMT2B | tRNA methyltransferase 2 homolog B | ||
| 213338_at | 0.048 | −1.780 | TMEM158 | transmembrane protein 158 (gene/pseudogene) | ||
| 227264_at | 0.031 | −2.005 | TRAF6 | TNF receptor associated factor 6 | ||
| 243624_at | 0.040 | −1.769 | PIAS2 | protein inhibitor of activated STAT 2 |
| GO Analysis | ID | Description | p Value | Gene |
|---|---|---|---|---|
| BP | GO:0001773 | myeloid dendritic cell activation | 0.0001 | TRAF6, PYCARD |
| GO:0006919 | activation of cysteine-type endopeptidase activity involved in apoptotic process | 0.001 | SLC22A2, PYCARD | |
| GO:0042102 | positive regulation of T cell proliferation | 0.002 | TRAF6, PYCARD | |
| GO:0002495 | antigen processing and presentation of peptide antigen via MHC class II | 0.002 | TRAF6, PYCARD | |
| GO:0002504 | antigen processing and presentation of peptide or polysaccharide antigen via MHC class II | 0.002 | TRAF6, PYCARD | |
| GO:0002821 | positive regulation of adaptive immune response | 0.002 | TRAF6, PYCARD | |
| GO:0042476 | odontogenesis | 0.003 | TRAF6, CEMP1 | |
| GO:0043280 | positive regulation of cysteine-type endopeptidase activity involved in apoptotic process | 0.003 | SLC22A2, PYCARD | |
| GO:0050671 | positive regulation of lymphocyte proliferation | 0.003 | TRAF6, PYCARD | |
| GO:0032946 | positive regulation of mononuclear cell proliferation | 0.003 | TRAF6, PYCARD | |
| CC | GO:0035631 | CD40 receptor complex | 0.006 | TRAF6 |
| GO:0061702 | inflammasome complex | 0.010 | PYCARD | |
| GO:1902554 | serine/threonine protein kinase complex | 0.049 | PYCARD | |
| MF | GO:0031996 | thioesterase binding | 0.008 | TRAF6 |
| GO:0043422 | protein kinase B binding | 0.008 | TRAF6 | |
| GO:0070700 | BMP receptor binding | 0.009 | PYCARD | |
| GO:0015562 | efflux transmembrane transporter activity | 0.010 | SLC22A2 | |
| GO:0097153 | cysteine-type endopeptidase activity involved in apoptotic process | 0.011 | PYCARD | |
| GO:0005523 | tropomyosin binding | 0.011 | PYCARD | |
| GO:0008381 | mechanosensitive ion channel activity | 0.012 | TMC5 | |
| GO:0008656 | cysteine-type endopeptidase activator activity involved in apoptotic process | 0.012 | PYCARD | |
| GO:0015174 | basic amino acid transmembrane transporter activity | 0.012 | SLC22A2 | |
| GO:0031435 | mitogen-activated protein kinase binding | 0.013 | TRAF6 |
| ID | Description | p Value | Gene |
|---|---|---|---|
| hsa05133 | Pertussis | 0.001 | TRAF6, PYCARD |
| hsa05135 | Yersinia infection | 0.003 | TRAF6, PYCARD |
| hsa04120 | Ubiquitin mediated proteolysis pathway | 0.003 | TRAF6, PIAS2 |
| hsa04621 | NOD-like receptor signaling pathway | 0.005 | TRAF6, PYCARD |
| hsa05130 | Pathogenic Escherichia coli infection | 0.006 | TRAF6, PYCARD |
| hsa05417 | Lipid and atherosclerosis signaling pathways | 0.007 | TRAF6, PYCARD |
| hsa05131 | Shigellosis | 0.009 | TRAF6, PYCARD |
| hsa05132 | Salmonella infection | 0.009 | TRAF6, PYCARD |
| hsa05134 | Legionellosis | 0.035 | PYCARD |
| hsa04623 | Cytosolic DNA-sensing pathway | 0.038 | PYCARD |
| hsa04622 | RIG-I-like receptor signaling pathway | 0.042 | TRAF6 |
| hsa05140 | Leishmaniasis pathway | 0.047 | TRAF6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Chen, S.; Rao, X.; Yang, Y.; Chen, X.; Yu, L. The Inflammatory Gene PYCARD of the Entorhinal Cortex as an Early Diagnostic Target for Alzheimer’s Disease. Biomedicines 2023, 11, 194. https://doi.org/10.3390/biomedicines11010194
Liu W, Chen S, Rao X, Yang Y, Chen X, Yu L. The Inflammatory Gene PYCARD of the Entorhinal Cortex as an Early Diagnostic Target for Alzheimer’s Disease. Biomedicines. 2023; 11(1):194. https://doi.org/10.3390/biomedicines11010194
Chicago/Turabian StyleLiu, Wenjia, Sophia Chen, Xin Rao, Yisong Yang, Xiaodong Chen, and Liyang Yu. 2023. "The Inflammatory Gene PYCARD of the Entorhinal Cortex as an Early Diagnostic Target for Alzheimer’s Disease" Biomedicines 11, no. 1: 194. https://doi.org/10.3390/biomedicines11010194