
L-Serine Influences Epigenetic Modifications to Improve Cognition and Behaviors in Growth Hormone-Releasing Hormone Knockout Mice
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibody and Chemicals
2.2. Mouse Model and L-Serine Administration
2.3. Animal Behavioral Tests
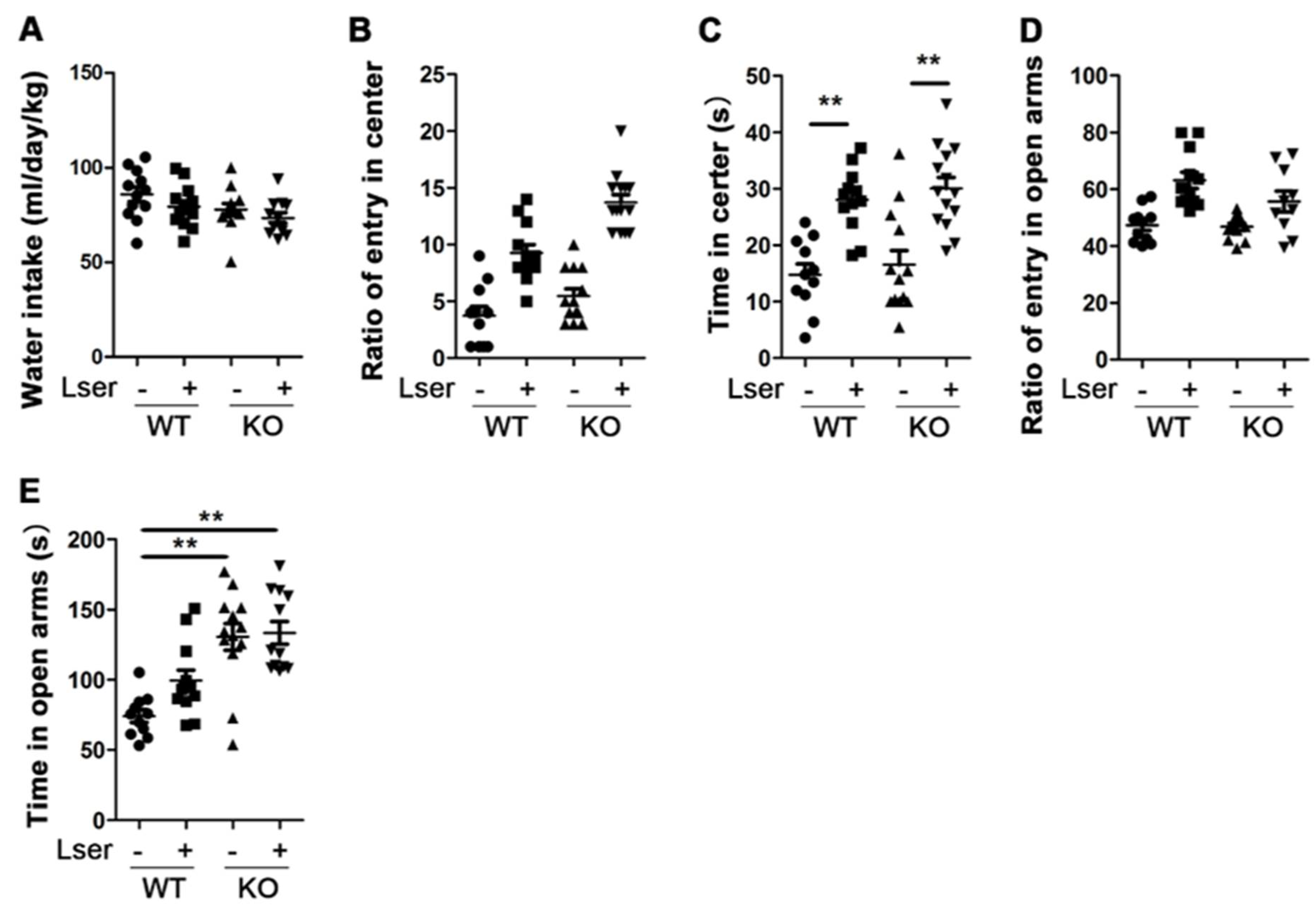
2.4. Elevated Zero Maze (EZM) and Open Field Test (OFT)
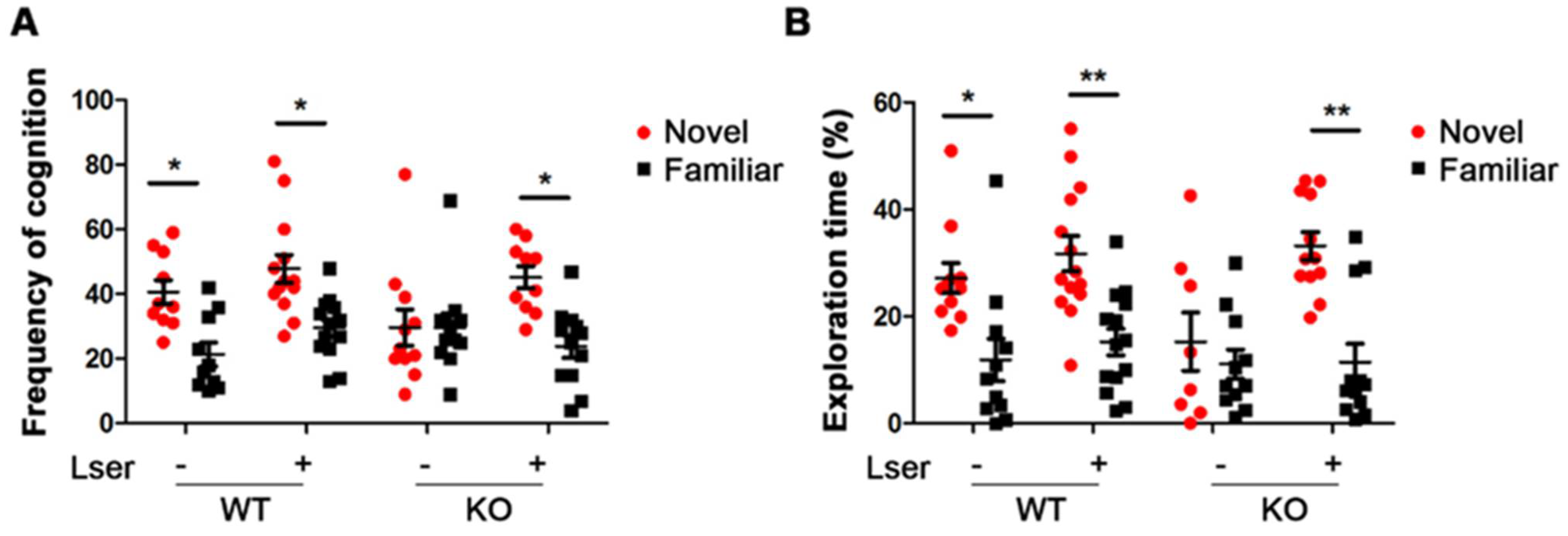
2.5. Novel Objective Recognition (NOR)
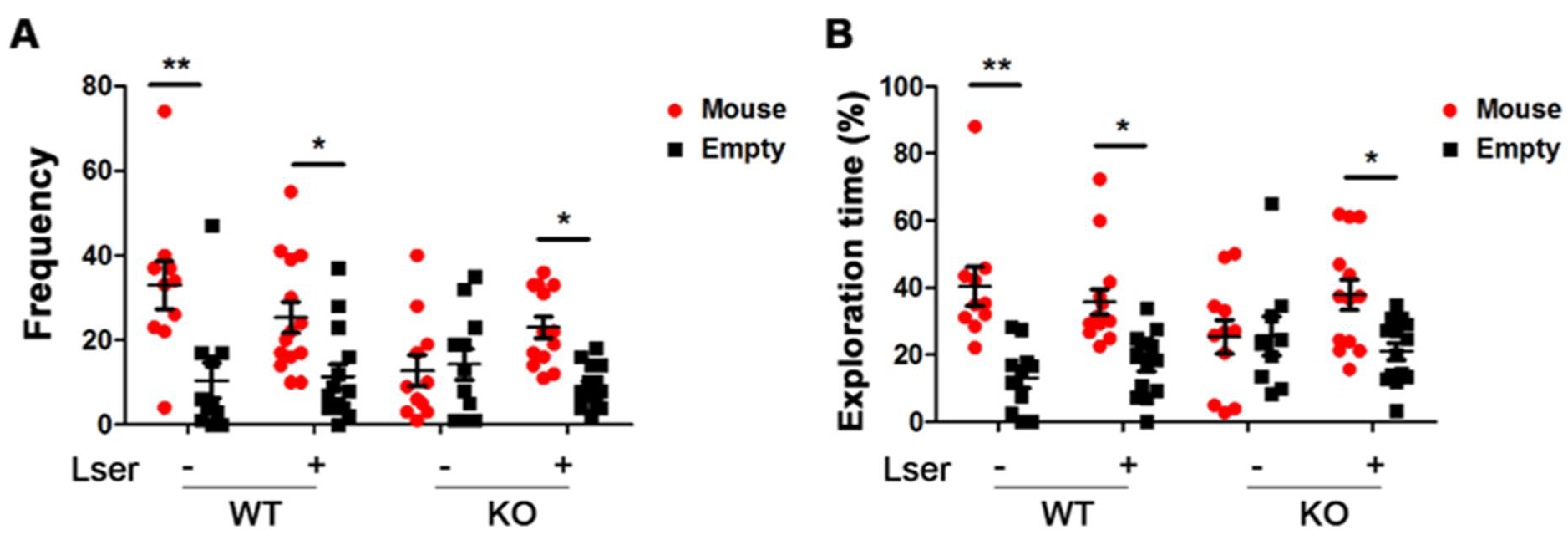
2.6. Three-Chamber Sociability
2.7. RNA Extraction and Real-Time PCR
2.8. Crude Synaptosome Preparation and Immunoblotting
2.9. Statistical Analysis
2.10. Study Flowchart

3. Results
3.1. L-Serine Intervention Decreased Anxiety-Related Behavior
3.2. L-Serine Intervention Improved Novel Object Recognition in GHRH-KO Mice
3.3. L-Serine Intervention Enhanced Sociability in GHRH-KO Mice
3.4. DNA Methylation Was Slightly Altered by Oral Administration of L-Serine
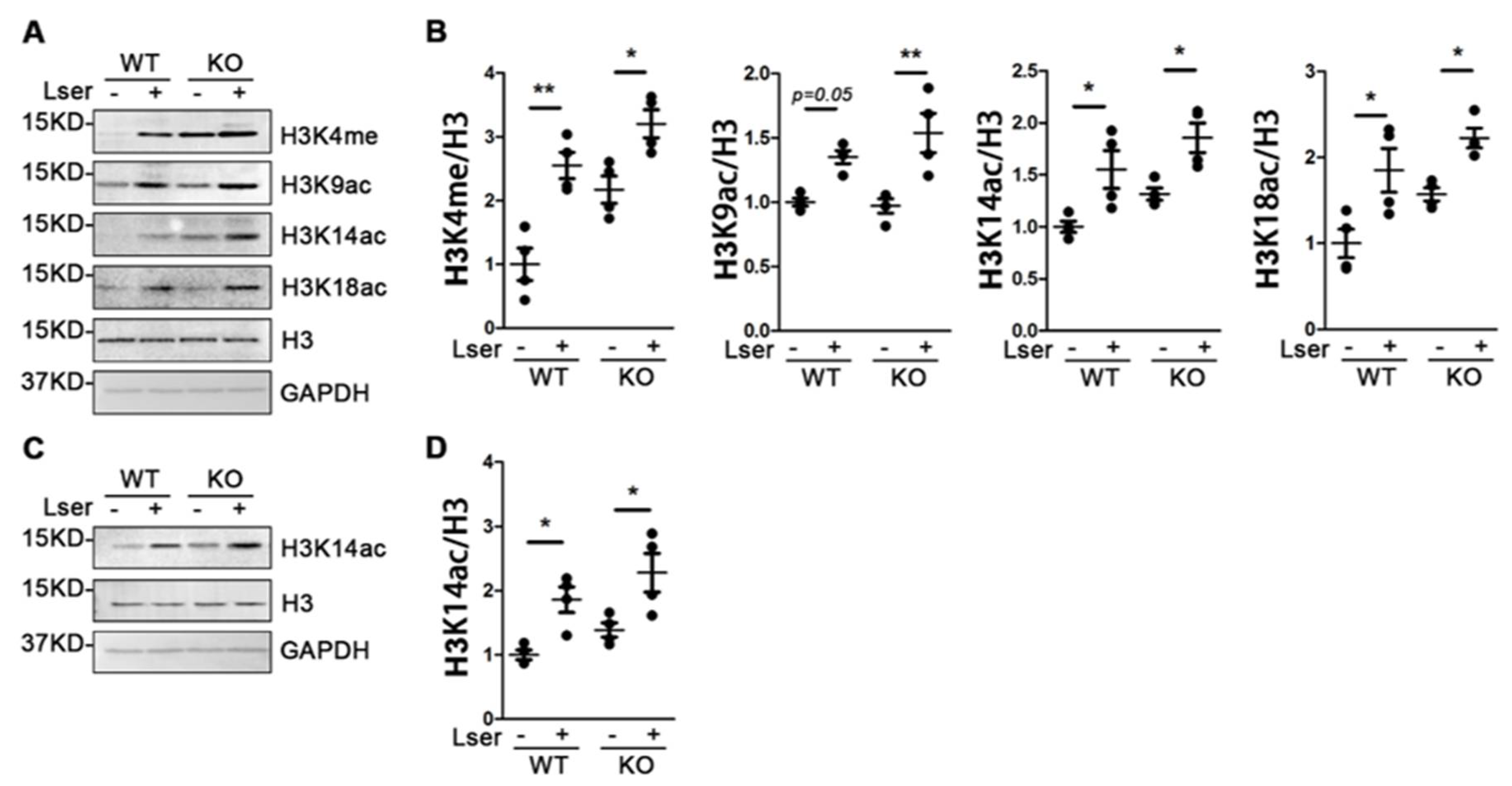
3.5. Oral Administration of L-Serine Changed Profiles of Histone H3 Modification
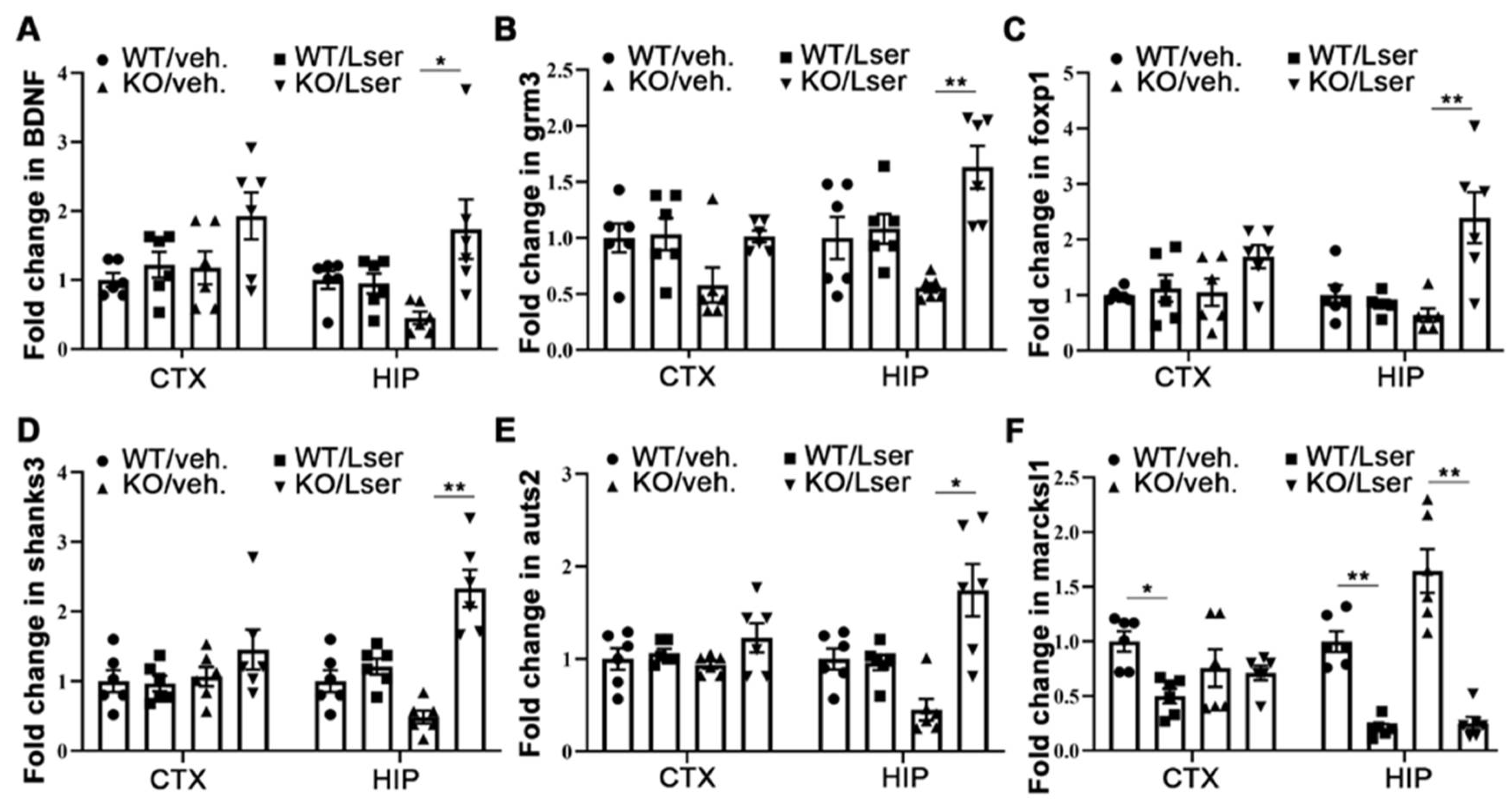
3.6. Epigenetic Modification-Regulated Genes Were Changes by L-Serine to Improve Behavioral Deficits
4. Discussion
4.1. GH Deficiency Induced Cognitive Deficit and Autism-Like Behavior
4.2. L-Serine Intervention Ameliorates Cognitive Deficit and Autism-Like Behaviors
4.3. L-Serine Improves GH Deficiency-Induced Behavioral Deficits Via Epigenetic Modifications
4.4. L-Serine Intervention Has an Anxiolytic Effect on Behavior
5. Summary
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GH | growth hormone |
| GHRH-KO | growth hormone-releasing hormone knockout |
| GHR-KO | growth hormone receptor knockout |
| Dnmt | DNA methyltransferase |
| Tet | Ten-eleven translocation |
| EZM | elevated zero maze |
| OFT | open field test |
| NOR | novel object recognition |
| ALS | amyotrophic lateral sclerosis |
| H3K4me | methylation of lysine 4 on histone H3 |
| H3K9ac | acetylation of lysine 9 on histone H3 |
| H3K14ac | acetylation of lysine 14 on histone H3 |
| H3K18ac | acetylation of lysine 18 on histone H3 |
| NAc | nucleus accumbens |
| Marcksl1 | myristoylated alanine-rich C-kinase substrate like 1 |
| BDNF | brain-derived neurotrophic factor |
| Foxp1 | forkhead box p1 |
| Auts2 | autism susceptibility candidate 2 |
| Shank3 | SH3 and multiple ankyrin repeat domains 3 |
| CNS | central nervous system |
| CTX | cerebral cortex |
| HIP | hippocampus |
References
- Hodes, G.E.; Epperson, C.N. Sex Differences in Vulnerability and Resilience to Stress Across the Life Span. Biol. Psychiatry 2019, 86, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhou, Y.; Campbell, S.L.; Le, T.; Li, E.; Sweatt, J.D.; Silva, A.J.; Fan, G. Dnmt1 and Dnmt3a maintain DNA methylation and regulate synaptic function in adult forebrain neurons. Nat. Neurosci. 2010, 13, 423–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Pena, C.J.; Purushothaman, I.; Engmann, O.; Walker, D.; Brown, A.N.; Issler, O.; Doyle, M.; Harrigan, E.; Mouzon, E.; et al. Tet1 in Nucleus Accumbens Opposes Depression- and Anxiety-Like Behaviors. Neuropsychopharmacology 2017, 42, 1657–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patki, G.; Solanki, N.; Atrooz, F.; Ansari, A.; Allam, F.; Jannise, B.; Maturi, J.; Salim, S. Novel mechanistic insights into treadmill exercise based rescue of social defeat-induced anxiety-like behavior and memory impairment in rats. Physiol. Behav. 2014, 130, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinowich, K.; Hattori, D.; Wu, H.; Fouse, S.; He, F.; Hu, Y.; Fan, G.; Sun, Y.E. DNA methylation-related chromatin remodeling in activity-dependent BDNF gene regulation. Science 2003, 302, 890–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durek, P.; Nordström, K.; Gasparoni, G.; Salhab, A.; Kressler, C.; de Almeida, M.; Bassler, K.; Ulas, T.; Schmidt, F.; Xiong, J.; et al. Epigenomic Profiling of Human CD4(+) T Cells Supports a Linear Differentiation Model and Highlights Molecular Regulators of Memory Development. Immunity 2016, 45, 1148–1161. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Wang, X.; Li, X.L.; Towers, A.; Cao, X.; Wang, P.; Bowman, R.; Yang, H.; Goldstein, J.; Li, Y.J.; et al. Epigenetic dysregulation of SHANK3 in brain tissues from individuals with autism spectrum disorders. Hum. Mol. Genet. 2014, 23, 1563–1578. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.A.; Jin, S.G.; Li, A.X.; Liu, J.; Huang, Z.; Wu, X.; Kim, B.W.; Johnson, J.; Bilbao, A.V.; Tao, S.; et al. Reprogramming of DNA methylation at NEUROD2-bound sequences during cortical neuron differentiation. Sci. Adv. 2019, 5, eaax0080. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ichida, J.K. Directly Reprogrammed Neurons Still Have Some Growing Up to Do. Cell Stem Cell 2019, 25, 1–2. [Google Scholar] [CrossRef]
- Bartke, A. Healthspan and longevity can be extended by suppression of growth hormone signaling. Mamm. Genome 2016, 27, 289–299. [Google Scholar] [CrossRef]
- Kinney, B.A.; Meliska, C.J.; Steger, R.W.; Bartke, A. Evidence that Ames dwarf mice age differently from their normal siblings in behavioral and learning and memory parameters. Horm. Behav. 2001, 39, 277–284. [Google Scholar] [CrossRef]
- Ransome, M.I.; Goldshmit, Y.; Bartlett, P.F.; Waters, M.J.; Turnley, A.M. Comparative analysis of CNS populations in knockout mice with altered growth hormone responsiveness. Eur. J. Neurosci. 2004, 19, 2069–2079. [Google Scholar] [CrossRef] [PubMed]
- Hascup, K.N.; Lynn, M.K.; Fitzgerald, P.J.; Randall, S.; Kopchick, J.J.; Boger, H.A.; Bartke, A.; Hascup, E.R. Enhanced Cognition and Hypoglutamatergic Signaling in a Growth Hormone Receptor Knockout Mouse Model of Successful Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leone, S.; Recinella, L.; Chiavaroli, A.; Ferrante, C.; Orlando, G.; Vacca, M.; Salvatori, R.; Brunetti, L. Behavioural phenotyping, learning and memory in young and aged growth hormone-releasing hormone-knockout mice. Endocr. Connect. 2018, 7, 924–931. [Google Scholar] [CrossRef] [Green Version]
- Leone, S.; Shohreh, R.; Manippa, F.; Recinella, L.; Ferrante, C.; Orlando, G.; Salvatori, R.; Vacca, M.; Brunetti, L. Behavioural phenotyping of male growth hormone-releasing hormone (GHRH) knockout mice. Growth Horm. IGF Res. 2014, 24, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Coleman, K.; Dahl, R.E.; Ryan, N.D.; Cameron, J.L. Growth hormone response to growth hormone-releasing hormone and clonidine in young monkeys: Correlation with behavioral characteristics. J. Child. Adolesc. Psychopharmacol. 2003, 13, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Arwert, L.I.; Deijen, J.B.; Müller, M.; Drent, M.L. Long-term growth hormone treatment preserves GH-induced memory and mood improvements: A 10-year follow-up study in GH-deficient adult men. Horm. Behav. 2005, 47, 343–349. [Google Scholar] [CrossRef]
- Sonntag, W.E.; Lynch, C.; Thornton, P.; Khan, A.; Bennett, S.; Ingram, R. The effects of growth hormone and IGF-1 deficiency on cerebrovascular and brain ageing. J. Anat. 2000, 197, 575–585. [Google Scholar] [CrossRef]
- Ramsey, M.M.; Weiner, J.L.; Moore, T.P.; Carter, C.S.; Sonntag, W.E. Growth hormone treatment attenuates age-related changes in hippocampal short-term plasticity and spatial learning. Neuroscience 2004, 129, 119–127. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Dunlop, R.A.; Powell, J.T.; Banack, S.A.; Cox, P.A. L-Serine: A Naturally-Occurring Amino Acid with Therapeutic Potential. Neurotox. Res. 2018, 33, 213–221. [Google Scholar] [CrossRef]
- Auranen, M.; Toppila, J.; Suriyanarayanan, S.; Lone, M.A.; Paetau, A.; Tyynismaa, H.; Hornemann, T.; Ylikallio, E. Clinical and metabolic consequences of L-serine supplementation in hereditary sensory and autonomic neuropathy type 1C. Cold Spring Harb. Mol. Case Stud. 2017, 3, a002212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, D.; Olivella, M.; Grau, C.; Armstrong, J.; Alcon, C.; Gasull, X.; Santos-Gómez, A.; Locubiche, S.; Gómez de Salazar, M.; García-Díaz, R.; et al. L-Serine dietary supplementation is associated with clinical improvement of loss-of-function GRIN2B-related pediatric encephalopathy. Sci. Signal. 2019, 12, eaaw0936. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, K.; Penno, A.; Schmidt, B.P.; Lee, H.J.; Frosch, M.P.; von Eckardstein, A.; Brown, R.H.; Hornemann, T.; Eichler, F.S. Oral L-serine supplementation reduces production of neurotoxic deoxysphingolipids in mice and humans with hereditary sensory autonomic neuropathy type 1. J. Clin. Invest. 2011, 121, 4735–4745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, W.C.; Yin, H.Q.; Choi, H.S.; Choi, Y.J.; Kwak, H.C.; Kim, S.K.; Lee, B.H. L-serine supplementation attenuates alcoholic fatty liver by enhancing homocysteine metabolism in mice and rats. J. Nutr. 2015, 145, 260–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, P.A.; Davis, D.A.; Mash, D.C.; Metcalf, J.S.; Banack, S.A. Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain. Proc. Biol. Sci. 2016, 283, 20152397. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Cox, P.A.; Banack, S.A.; Lecusay, P.D.; Garamszegi, S.P.; Hagan, M.J.; Powell, J.T.; Metcalf, J.S.; Palmour, R.M.; Beierschmitt, A.; et al. l-Serine Reduces Spinal Cord Pathology in a Vervet Model of Preclinical ALS/MND. J. Neuropathol. Exp. Neurol. 2020, 79, 393–406. [Google Scholar] [CrossRef]
- Icyuz, M.; Fitch, M.; Zhang, F.; Challa, A.; Sun, L.Y. Physiological and metabolic features of mice with CRISPR/Cas9-mediated loss-of-function in growth hormone-releasing hormone. Aging 2020, 12, 9761–9780. [Google Scholar] [CrossRef]
- Braun, A.A.; Skelton, M.R.; Vorhees, C.V.; Williams, M.T. Comparison of the elevated plus and elevated zero mazes in treated and untreated male Sprague-Dawley rats: Effects of anxiolytic and anxiogenic agents. Pharmacol. Biochem. Behav. 2011, 97, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Polter, A.; Yang, S.; Zmijewska, A.A.; van Groen, T.; Paik, J.H.; Depinho, R.A.; Peng, S.L.; Jope, R.S.; Li, X. Forkhead box, class O transcription factors in brain: Regulation and behavioral manifestation. Biol. Psychiatry 2009, 65, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Leger, M.; Quiedeville, A.; Bouet, V.; Haelewyn, B.; Boulouard, M.; Schumann-Bard, P.; Freret, T. Object recognition test in mice. Nat. Protoc. 2013, 8, 2531–2537. [Google Scholar] [CrossRef]
- Zhang, F.; Icyuz, M.; Liu, Z.; Fitch, M.; Sun, L.Y. Insulin sensitivity in long-lived growth hormone-releasing hormone knockout mice. Aging 2020, 12, 18033–18051. [Google Scholar] [CrossRef]
- Icyuz, M.; Zhang, F.; Fitch, M.P.; Joyner, M.R.; Challa, A.K.; Sun, L.Y. Physiological and metabolic characteristics of novel double-mutant female mice with targeted disruption of both growth hormone-releasing hormone and growth hormone receptor. Aging Cell 2021, 20, e13339. [Google Scholar] [CrossRef]
- Cummings, C.M.; Caporino, N.E.; Kendall, P.C. Comorbidity of anxiety and depression in children and adolescents: 20 years after. Psychol. Bull. 2014, 140, 816–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, R. Chronic stress, drug use, and vulnerability to addiction. Ann. N. Y. Acad. Sci. 2008, 1141, 105–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigemi, K.; Tsuneyoshi, Y.; Yamada, S.; Kabuki, Y.; Hayamizu, K.; Denbow, D.M.; Furuse, M. Oral administration of L-serine reduces the locomotor activity of socially isolated rats. Neurosci. Lett. 2010, 468, 75–79. [Google Scholar] [CrossRef]
- Holm, L.J.; Haupt-Jorgensen, M.; Larsen, J.; Giacobini, J.D.; Bilgin, M.; Buschard, K. L-serine supplementation lowers diabetes incidence and improves blood glucose homeostasis in NOD mice. PLoS ONE 2018, 13, e0194414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutcliffe, J.S.; Marshall, K.M.; Neill, J.C. Influence of gender on working and spatial memory in the novel object recognition task in the rat. Behav. Brain Res. 2007, 177, 117–125. [Google Scholar] [CrossRef]
- Rajagopal, L.; Massey, B.W.; Huang, M.; Oyamada, Y.; Meltzer, H.Y. The novel object recognition test in rodents in relation to cognitive impairment in schizophrenia. Curr. Pharm. Des. 2014, 20, 5104–5114. [Google Scholar] [CrossRef]
- Lai, M.C.; Szatmari, P. Sex and gender impacts on the behavioural presentation and recognition of autism. Curr. Opin. Psychiatry 2020, 33, 117–123. [Google Scholar] [CrossRef]
- Moy, S.S.; Nadler, J.J.; Perez, A.; Barbaro, R.P.; Johns, J.M.; Magnuson, T.R.; Piven, J.; Crawley, J.N. Sociability and preference for social novelty in five inbred strains: An approach to assess autistic-like behavior in mice. Genes Brain Behav. 2004, 3, 287–302. [Google Scholar] [CrossRef]
- Armstrong, V.L.; Rakoczy, S.; Rojanathammanee, L.; Brown-Borg, H.M. Expression of DNA methyltransferases is influenced by growth hormone in the long-living Ames dwarf mouse in vivo and in vitro. J. Gerontology. Ser. A Biol. Sci. Med. Sci. 2014, 69, 923–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultan, F.A.; Day, J.J. Epigenetic mechanisms in memory and synaptic function. Epigenomics 2011, 3, 157–181. [Google Scholar] [CrossRef] [Green Version]
- Matzel, L.D.; Grossman, H.; Light, K.; Townsend, D.; Kolata, S. Age-related declines in general cognitive abilities of Balb/C mice are associated with disparities in working memory, body weight, and general activity. Learn Mem. 2008, 15, 733–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattison, J.A.; Wright, C.; Bronson, R.T.; Roth, G.S.; Ingram, D.K.; Bartke, A. Studies of aging in ames dwarf mice: Effects of caloric restriction. J. Am. Aging Assoc. 2000, 23, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinney-Forshee, B.A.; Kinney, N.E.; Steger, R.W.; Bartke, A. Could a deficiency in growth hormone signaling be beneficial to the aging brain? Physiol. Behav. 2004, 80, 589–594. [Google Scholar] [CrossRef]
- Arwert, L.I.; Veltman, D.J.; Deijen, J.B.; van Dam, P.S.; Delemarre-van deWaal, H.A.; Drent, M.L. Growth hormone deficiency and memory functioning in adults visualized by functional magnetic resonance imaging. Neuroendocrinology 2005, 82, 32–40. [Google Scholar] [CrossRef]
- Sharma, S.; Haselton, J.; Rakoczy, S.; Branshaw, S.; Brown-Borg, H.M. Spatial memory is enhanced in long-living Ames dwarf mice and maintained following kainic acid induced neurodegeneration. Mech. Ageing Dev. 2010, 131, 422–435. [Google Scholar] [CrossRef] [Green Version]
- Ganote, C.E.; Peterson, D.R.; Carone, F.A. The nature of D-serine--induced nephrotoxicity. Am. J. Pathol. 1974, 77, 269–282. [Google Scholar]
- Wang, G.H.; Jiang, Z.L.; Chen, Z.Q.; Li, X.; Peng, L.L. Neuroprotective effect of L-serine against temporary cerebral ischemia in rats. J. Neurosci. Res. 2010, 88, 2035–2045. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Lobner, D.; Banack, S.A.; Cox, G.A.; Nunn, P.B.; Wyatt, P.B.; Cox, P.A. Analysis of BMAA enantiomers in cycads, cyanobacteria, and mammals: In vivo formation and toxicity of D-BMAA. Amino. Acids 2017, 49, 1427–1439. [Google Scholar] [CrossRef]
- Manns, J.R.; Eichenbaum, H. A cognitive map for object memory in the hippocampus. Learn Mem. 2009, 16, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Bredy, T.W.; Wu, H.; Crego, C.; Zellhoefer, J.; Sun, Y.E.; Barad, M. Histone modifications around individual BDNF gene promoters in prefrontal cortex are associated with extinction of conditioned fear. Learn Mem. 2007, 14, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Moghaddam, B. Targeting metabotropic glutamate receptors for treatment of the cognitive symptoms of schizophrenia. Psychopharmacology 2004, 174, 39–44. [Google Scholar] [CrossRef]
- De Filippis, B.; Lyon, L.; Taylor, A.; Lane, T.; Burnet, P.W.; Harrison, P.J.; Bannerman, D.M. The role of group II metabotropic glutamate receptors in cognition and anxiety: Comparative studies in GRM2(-/-), GRM3(-/-) and GRM2/3(-/-) knockout mice. Neuropharmacology 2015, 89, 19–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacon, C.; Schneider, M.; Le Magueresse, C.; Froehlich, H.; Sticht, C.; Gluch, C.; Monyer, H.; Rappold, G.A. Brain-specific Foxp1 deletion impairs neuronal development and causes autistic-like behaviour. Mol. Psychiatry 2015, 20, 632–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peça, J.; Feliciano, C.; Ting, J.T.; Wang, W.; Wells, M.F.; Venkatraman, T.N.; Lascola, C.D.; Fu, Z.; Feng, G. Shank3 mutant mice display autistic-like behaviours and striatal dysfunction. Nature 2011, 472, 437–442. [Google Scholar] [CrossRef] [Green Version]
- Oksenberg, N.; Stevison, L.; Wall, J.D.; Ahituv, N. Function and regulation of AUTS2, a gene implicated in autism and human evolution. PLoS Genet. 2013, 9, e1003221. [Google Scholar] [CrossRef] [Green Version]
- Salk, R.H.; Hyde, J.S.; Abramson, L.Y. Gender differences in depression in representative national samples: Meta-analyses of diagnoses and symptoms. Psychol. Bull. 2017, 143, 783–822. [Google Scholar] [CrossRef]
- Tanaka, T.; Shimizu, S.; Ueno, M.; Fujihara, Y.; Ikawa, M.; Miyata, S. MARCKSL1 Regulates Spine Formation in the Amygdala and Controls the Hypothalamic-Pituitary-Adrenal Axis and Anxiety-Like Behaviors. EBioMedicine 2018, 30, 62–73. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Protein Name | Monoclonal or Polyclonal | Host Species | Commercial Supplier | Catalogue Number |
|---|---|---|---|---|---|
| 1 | H3K4me | polyclonal | Rabbit | Abcam | ab272143 |
| 2 | H3K27me | monoclonal | Rabbit | Abcam | ab192985 |
| 3 | H3K9ac | Monoclonal | Rabbit | Cell Signaling Technology | #9649 |
| 4 | H3K14ac | Monoclonal | Rabbit | Cell Signaling Technology | #7627 |
| 5 | H3K18ac | Monoclonal | Rabbit | Cell Signaling Technology | #13998 |
| 6 | H3K27ac | Monoclonal | Rabbit | Cell Signaling Technology | #8173 |
| 7 | H3 | Monoclonal | Rabbit | Cell Signaling Technology | #4499 |
| 8 | GAPDH | Monoclonal | Mouse | Cell Signaling Technology | #97166 |
| Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| Dnmt1 | TTGAAACTTCACCTAGTTCCGTGGC | CTGCAGCACCACTCTCTGTGTCTAC |
| Dnmt3α | GAAAGGGTATGGGAGTTACATAGAG | CGGTGTGAAATATCGCAGTTTAAG |
| Dnmt3β | CACACTCTGGAGAAAGCCAGGGTTC | AGTCATTGGTTGTGCGTCTTCGACT |
| Tet1 | GACCCTCATAAGCAGAGAGGAAAA | TCTTCATTTCCAAGTCGACAGTCT |
| Tet2 | GGAGCAGAAGGAAGCAAGATGG | ATGAATCCAGCAGCACCGTCC |
| Tet3 | AAATGCTCGTGAAGGAACGGG | CTGCCTTGAATCTCCATGGTACAC |
| BDNF | CGAGATCGGGGCTGGAGA | GGTCATCACTCTTCTCACCTGG |
| Grm3 | TCATTGGCGGTTCGTACAGC | ACTGGTGGAGGCGTAGCTTAT |
| Foxp1 | CACCTCAGGTTATCACTCCTCA | AGCTGCAACTGTTCCTGTTGT |
| Shank3 | CAACATGGGTGCTCAGAATGC | ACGACATCGGAGTCTTTGTGG |
| Auts2 | TCAGCCACTCACACCACTACA | AGCCCTTGGATTCTCCGCT |
| Marcksl1 | GCCAACGGACAGGAGAATGG | CTCGATGGCATCACCAGTAGC |
| β-actin | TCTTTGCAGCTCCTTCGTTGCC | CTGACCCATTCCCACCATCACAC |
| GAPDH | CCTGGAGAAACCTGCCAAGTATGATG | AAGAGTGGGAGTTGCTGTTGAAGTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Icyuz, M.; Tollefsbol, T.; Cox, P.A.; Banack, S.A.; Sun, L.Y. L-Serine Influences Epigenetic Modifications to Improve Cognition and Behaviors in Growth Hormone-Releasing Hormone Knockout Mice. Biomedicines 2023, 11, 104. https://doi.org/10.3390/biomedicines11010104
Zhang F, Icyuz M, Tollefsbol T, Cox PA, Banack SA, Sun LY. L-Serine Influences Epigenetic Modifications to Improve Cognition and Behaviors in Growth Hormone-Releasing Hormone Knockout Mice. Biomedicines. 2023; 11(1):104. https://doi.org/10.3390/biomedicines11010104
Chicago/Turabian StyleZhang, Fang, Mert Icyuz, Trygve Tollefsbol, Paul Alan Cox, Sandra Anne Banack, and Liou Y. Sun. 2023. "L-Serine Influences Epigenetic Modifications to Improve Cognition and Behaviors in Growth Hormone-Releasing Hormone Knockout Mice" Biomedicines 11, no. 1: 104. https://doi.org/10.3390/biomedicines11010104