
Subtype-Specific Tumour Immune Microenvironment in Risk of Recurrence of Ductal Carcinoma In Situ: Prognostic Value of HER2

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Molecular Subtypes of Breast Carcinoma: Immune Characterization
3. Pathological Evaluation of TILs in DCIS: Approaches and Current Clinical Utility

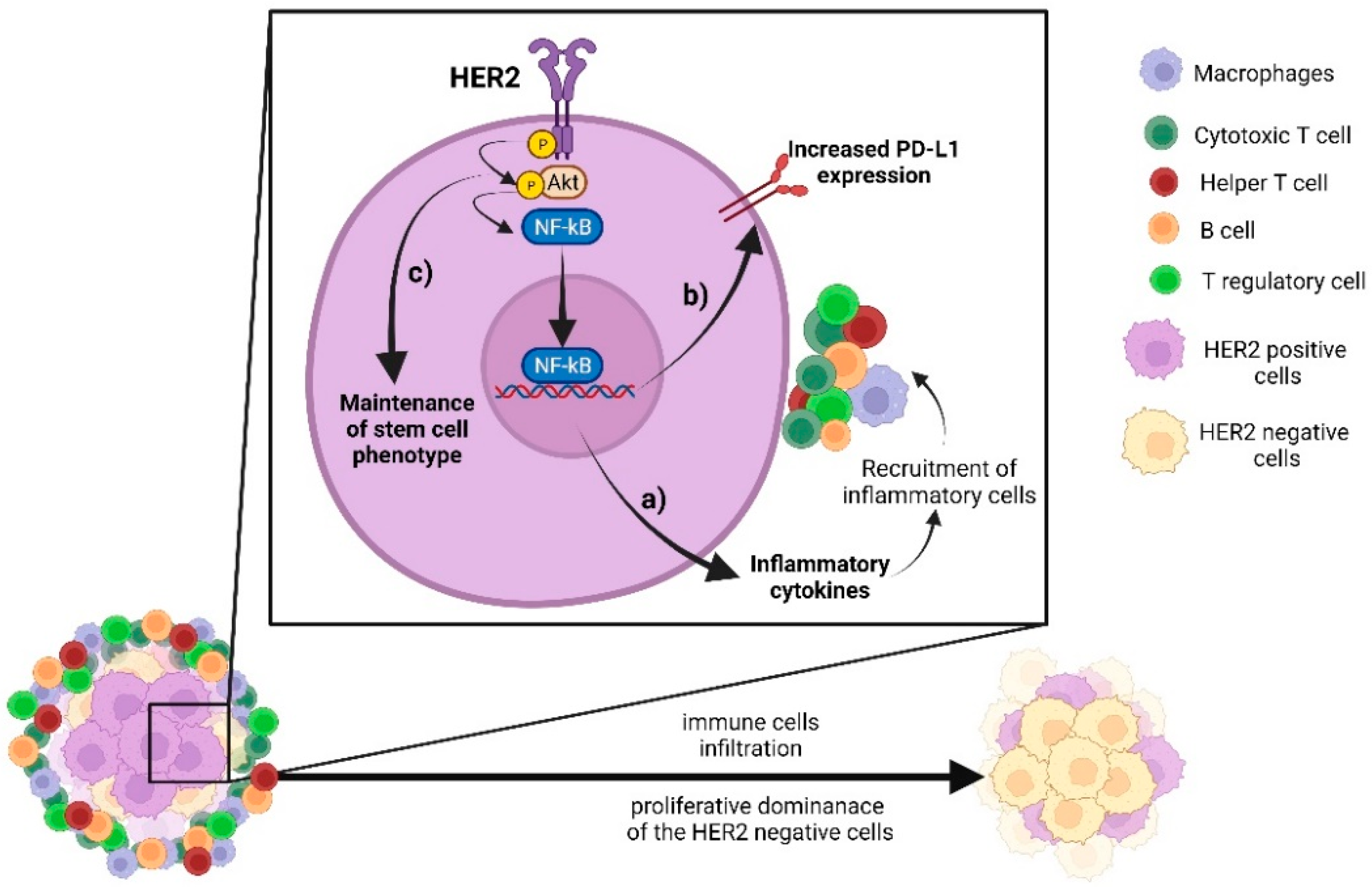{kind=link}
| TIME: Related Risk of Recurrence | Clinico-Pathological Variables | High TILs | High CD8 | High CD4 | High FoxP3 | High PD-1/PD-L1 | High TILB | High Macrophages |
|---|---|---|---|---|---|---|---|---|
| increased | HER2 + | Pruneri [27]; N = 1488 (2016), Thike [37]; N = 198 (2019) | Campbell [49]; N = 117 (2017), Thike [37]; N = 198 (2019), Semeraro [35]; N = 248 (2016) | Thike [37]; N = 198 (2019) | Toss [45]; N = 700 (2020) | Toss [45]; N = 700 (2020) | Milgy [36]; N = 80 (2017) | |
| ER - | Darvihian [28]; N = 688; (2019), Toss [34]; N = 816 (2017) | Chen [46]; N = 80 (2020) | ||||||
| Not reported | High grade | Darvishian [28]; N = 688; (2019), Pruneri [27]; N = 1488 (2016), Toss [34]; N = 816 (2017), Morita [38]; N = 46 (2017), Beguinot [40]; N = 129 (2018), Thike [37]; N = 198 (2019) Chen [41]; N = 198 (2021), Toss [45]; N = 700 (2020) Semeraro [35]; N = 248 (2016) Hendry [32]; N = 138 (2017) | Campbell [49]; N = 117 (2017), Beguinot [40]; N = 129 (2018), Semeraro [35]; N = 248 (2016) | Campbell [49]; N = 117 (2017), Beguinot [40]; N = 129 (2018), Chen [41]; N = 198 (2021) | Campbell [49]; N = 117 (2017), Chen [41]; N = 198 (2021), Toss [45]; N = 700 (2020) | Chen [41]; N = 198 (2021), Toss [45]; N = 700 (2020) Thompson [29]; N = 27 (2016), Hendry [32]; N = 138 (2017) | Campbell [49]; N = 117 (2017), Beguinot [40]; N = 129 (2018) | Capmbell [49]; N = 117 (2017), Chen [46]; N = 80 (2020), Chen [41]; N = 198 (2021) |
| Increased tumour size | Campbell [49]; N = 117 (2017), Darvishian [28]; N = 688; (2019) | Campbell [49]; N = 117 (2017) | Milgy [36]; N = 80 (2017) Campbell [49]; N = 117 (2017) | |||||
| Presence of microinvasion | Morita [38]; N = 46 (2017), Toss [34]; N = 816 (2017) | Lv [39]; N = 85 (2019), Alcazar [22]; N = 36 (2017), Beguinot [40]; N = 129 (2018) | Beguinot [40]; N = 129 (2018) | Chen [41]; N = 198 (2021), Beguinot [40]; N = 129 (2018) | Lv [39]; N = 85 (2019), Beguinot [40]; N = 129 (2018) | Milgy [36]; N = 80 (2017) | Chen [46]; N = 80 (2020), Beguinot [40]; N = 129 (2018) | |
| High mitotic index | Campbell [49]; N = 117 (2017) | Beguinot [40]; N = 129 (2018) | Beguinot [40]; N = 129 (2018), Campbell [49]; N = 117 (2017) | Beguinot [40]; N = 129 (2018) | Beguinot [40]; N = 129 (2018) | Campbell [49]; N = 117 (2017) | Beguinot [40]; N = 129 (2018) Campbell [49]; N = 117 (2017) |
4. Standard Parameters of TIME
4.1. CD8+ T Cytotoxic Cells
4.2. CD4+ T Helper Cells (CD4+ Th Cells)
4.3. CD4+CD25+FOXP3+ Regulatory T Cells (T Reg Cells)
4.4. Markers of Immune Exhaustion of T Cells
4.5. Tumour Infiltrating B Cells (TILBs)
4.6. Macrophages
5. Crosstalk between HER2 Pathway and Immune Signalling
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salvatorelli, L.; Puzzo, L.; Vecchio, G.M.; Caltabiano, R.; Virzì, V.; Magro, G. Ductal Carcinoma In Situ of the Breast: An Update with Emphasis on Radiological and Morphological Features as Predictive Prognostic Factors. Cancers 2020, 12, 609. [Google Scholar] [CrossRef]
- Habel, L.A.; Daling, J.R.; Newcomb, P.A.; Self, S.G.; Porter, P.L.; Stanford, J.L.; Seidel, K.; Weiss, N.S. Risk of Recurrence after Ductal Carcinoma in Situ of the Breast. Cancer Epidemiol. Prev. Biomark. 1998, 7, 689–696. [Google Scholar]
- Elshof, L.E.; Schaapveld, M.; Schmidt, M.K.; Rutgers, E.J.; van Leeuwen, F.E.; Wesseling, J. Subsequent Risk of Ipsilateral and Contralateral Invasive Breast Cancer after Treatment for Ductal Carcinoma in Situ: Incidence and the Effect of Radiotherapy in a Population-Based Cohort of 10,090 Women. Breast Cancer Res. Treat. 2016, 159, 553–563. [Google Scholar] [CrossRef]
- Nofech-Mozes, S.; Spayne, J.; Rakovitch, E.; Kahn, H.J.; Seth, A.; Pignol, J.-P.; Lickley, L.; Paszat, L.; Hanna, W. Biological Markers Predictive of Invasive Recurrence in DCIS. Clin. Med. Oncol. 2008, 2, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Simpson, P.T.; Da Silva, L.M.; Lakhani, S.R. In Situ Carcinoma—Can We Predict Which Patient Will Come Back with a Recurrence? Cancer Cell 2007, 12, 409–411. [Google Scholar] [CrossRef]
- Allen, M.D.; Marshall, J.F.; Jones, J.L. Avβ6 Expression in Myoepithelial Cells: A Novel Marker for Predicting DCIS Progression with Therapeutic Potential. Cancer Res. 2014, 74, 5942–5947. [Google Scholar] [CrossRef] [PubMed]
- De Roos, M.A.; de Bock, G.H.; de Vries, J.; van der Vegt, B.; Wesseling, J. P53 Overexpression Is a Predictor of Local Recurrence after Treatment for Both in Situ and Invasive Ductal Carcinoma of the Breast. J. Surg. Res. 2007, 140, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Jirström, K.; Ringberg, A.; Fernö, M.; Anagnostaki, L.; Landberg, G. Tissue Microarray Analyses of G1/S-Regulatory Proteins in Ductal Carcinoma in Situ of the Breast Indicate That Low Cyclin D1 Is Associated with Local Recurrence. Br. J. Cancer 2003, 89, 1920–1926. [Google Scholar] [CrossRef]
- Miligy, I.M.; Gorringe, K.L.; Toss, M.S.; Al-Kawaz, A.A.; Simpson, P.; Diez-Rodriguez, M.; Nolan, C.C.; Ellis, I.O.; Green, A.R.; Rakha, E.A. Thioredoxin-Interacting Protein Is an Independent Risk Stratifier for Breast Ductal Carcinoma In Situ. Mod. Pathol. 2018, 31, 1807–1815. [Google Scholar] [CrossRef]
- Poulakaki, N.; Makris, G.-M.; Papanota, A.-M.; Marineli, F.; Marinelis, A.; Battista, M.-J.; Boehm, D.; Psyrri, A.; Sergentanis, T.N. Ki-67 Expression as a Factor Predicting Recurrence of Ductal Carcinoma In Situ of the Breast: A Systematic Review and Meta-Analysis. Clin. Breast Cancer 2018, 18, 157–167.e6. [Google Scholar] [CrossRef]
- Toss, M.S.; Miligy, I.M.; Gorringe, K.L.; Aleskandarany, M.A.; Alkawaz, A.; Mittal, K.; Aneja, R.; Ellis, I.O.; Green, A.R.; Rakha, E.A. Collagen (XI) Alpha-1 Chain Is an Independent Prognostic Factor in Breast Ductal Carcinoma in Situ. Mod. Pathol. 2019, 32, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Toss, M.S.; Miligy, I.M.; Haj-Ahmad, R.; Gorringe, K.L.; AlKawaz, A.; Mittal, K.; Ellis, I.O.; Green, A.R.; Rakha, E.A. The Prognostic Significance of Lysosomal Protective Protein (Cathepsin A) in Breast Ductal Carcinoma in Situ. Histopathology 2019, 74, 1025–1035. [Google Scholar] [CrossRef]
- Gauthier, M.L.; Berman, H.K.; Miller, C.; Kozakeiwicz, K.; Chew, K.; Moore, D.; Rabban, J.; Chen, Y.Y.; Kerlikowske, K.; Tlsty, T.D. Abrogated Response to Cellular Stress Identifies DCIS Associated with Subsequent Tumor Events and Defines Basal-like Breast Tumors. Cancer Cell 2007, 12, 479–491. [Google Scholar] [CrossRef]
- Oda, G.; Nakagawa, T.; Ogawa, A.; Kumaki, Y.; Hosoya, T.; Sugimoto, H.; Ishiba, T.; Mio, M.; Fujioka, T.; Kubota, K.; et al. Predictors for Upstaging of Ductal Carcinoma in Situ (DCIS) to Invasive Carcinoma in Non-Mass-Type DCIS. Mol. Clin. Oncol. 2020, 13, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Latta, E.K.; Tjan, S.; Parkes, R.K.; O’Malley, F.P. The Role of HER2/Neu Overexpression/Amplification in the Progression of Ductal Carcinoma in Situ to Invasive Carcinoma of the Breast. Mod. Pathol. 2002, 15, 1318–1325. [Google Scholar] [CrossRef]
- Roses, R.E.; Paulson, E.C.; Sharma, A.; Schueller, J.E.; Nisenbaum, H.; Weinstein, S.; Fox, K.R.; Zhang, P.J.; Czerniecki, B.J. HER-2/Neu Overexpression as a Predictor for the Transition from in Situ to Invasive Breast Cancer. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1385–1389. [Google Scholar] [CrossRef] [PubMed]
- Borgquist, S.; Zhou, W.; Jirström, K.; Amini, R.M.; Sollie, T.; Sørlie, T.; Blomqvist, C.; Butt, S.; Wärnberg, F. The Prognostic Role of HER2 Expression in Ductal Breast Carcinoma in Situ (DCIS); a Population-Based Cohort Study. BMC Cancer 2015, 15, 468. [Google Scholar] [CrossRef]
- Mustafa, R.E.; DeStefano, L.M.; Bahng, J.; Yoon-Flannery, K.; Fisher, C.S.; Zhang, P.J.; Tchou, J.; Czerniecki, B.J.; De La Cruz, L.M. Evaluating the Risk of Upstaging HER2-Positive DCIS to Invasive Breast Cancer. Ann. Surg. Oncol. 2017, 24, 2999–3003. [Google Scholar] [CrossRef]
- Thorat, M.A.; Levey, P.M.; Jones, J.L.; Pinder, S.E.; Bundred, N.J.; Fentiman, I.S.; Cuzick, J. Prognostic and Predictive Value of HER2 Expression in Ductal Carcinoma In Situ: Results from the UK/ANZ DCIS Randomized Trial. Clin. Cancer Res. 2021, 27, 5317–5324. [Google Scholar] [CrossRef]
- Thorat, M.A.; Levey, P.M.; Jones, J.L.; Pinder, S.E.; Bundred, N.J.; Fentiman, I.S.; Cuzick, J.; Louise Jones, J.; Pinder, S.E.; Bundred, N.J.; et al. Prognostic Value of ER and PgR Expression and the Impact of Multi-Clonal Expression for Recurrence in Ductal Carcinoma In Situ: Results from the UK/ANZ DCIS Trial. Clin. Cancer Res. 2021, 2, 2861–2868. [Google Scholar] [CrossRef]
- Trinh, A.; Gil Del Alcazar, C.R.; Shukla, S.A.; Chin, K.; Chang, Y.H.; Thibault, G.; Eng, J.; Jovanovic, B.; Aldaz, C.M.; Park, S.Y.; et al. Genomic Alterations during the in Situ to Invasive Ductal Breast Carcinoma Transition Shaped by the Immune System. Mol. Cancer Res. 2021, 19, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Gil Del Alcazar, C.R.; Huh, S.J.; Ekram, M.B.; Trinh, A.; Liu, L.L.; Beca, F.; Zi, X.; Kwak, M.; Bergholtz, H.; Su, Y.; et al. Immune Escape in Breast Cancer During In Situ to Invasive Carcinoma Transition. Cancer Discov. 2017, 7, 1098–1115. [Google Scholar] [CrossRef] [PubMed]
- Piasecka, D.; Braun, M.; Mieszkowska, M.; Kowalczyk, L.; Kopczynski, J.; Kordek, R.; Sadej, R.; Romanska, H.M. Upregulation of HIF1-α via an NF-ΚB/COX2 Pathway Confers Proliferative Dominance of HER2-Negative Ductal Carcinoma In Situ Cells in Response to Inflammatory Stimuli. Neoplasia 2020, 22, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Sadeghalvad, M.; Mohammadi-Motlagh, H.-R.; Rezaei, N. Immune Microenvironment in Different Molecular Subtypes of Ductal Breast Carcinoma. Breast Cancer Res. Treat. 2021, 185, 261–279. [Google Scholar] [CrossRef]
- Chen, X.-Y.Y.; Yeong, J.; Thike, A.A.; Bay, B.H.; Tan, P.H.; Aye, A.; Boon, T.; Bay, H.; Hoon, P. Prognostic Role of Immune Infiltrates in Breast Ductal Carcinoma in Situ. Breast Cancer Res. Treat. 2019, 177, 17–27. [Google Scholar] [CrossRef]
- Gil Del Alcazar, C.R.; Alečković, M.; Polyak, K. Immune Escape during Breast Tumor Progression. Cancer Immunol. Res. 2020, 8, 422–427. [Google Scholar] [CrossRef]
- Pruneri, G.; Lazzeroni, M.; Bagnardi, V.; Tiburzio, G.B.; Rotmensz, N.; DeCensi, A.; Guerrieri-Gonzaga, A.; Vingiani, A.; Curigliano, G.; Zurrida, S.; et al. The Prevalence and Clinical Relevance of Tumor-Infiltrating Lymphocytes (TILs) in Ductal Carcinoma in Situ of the Breast. Ann. Oncol. 2017, 28, 321–328. [Google Scholar] [CrossRef]
- Darvishian, F.; Ozerdem, U.; Adams, S.; Chun, J.; Pirraglia, E.; Kaplowitz, E.; Guth, A.; Axelrod, D.; Shapiro, R.; Price, A.; et al. Tumor-Infiltrating Lymphocytes in a Contemporary Cohort of Women with Ductal Carcinoma In Situ (DCIS). Ann. Surg. Oncol. 2019, 26, 3337–3343. [Google Scholar] [CrossRef]
- Thompson, E.; Taube, J.M.; Elwood, H.; Sharma, R.; Meeker, A.; Warzecha, H.N.; Argani, P.; Cimino-Mathews, A.; Emens, L.A.; Cimino-Mathews, A.; et al. The Immune Microenvironment of Breast Ductal Carcinoma in Situ. Mod. Pathol. 2016, 29, 249–258. [Google Scholar] [CrossRef]
- Abba, M.C.; Takata, Y.; Gaddis, S.; Shen, J.; Estecio, M.R.; Sahin, A.A. A Molecular Portrait of High-Grade Ductal Carcinoma In Situ. Cancer Res. 2015, 75, 3980–3990. [Google Scholar] [CrossRef]
- Ubago, J.M.; Blanco, L.Z.; Shen, T.; Siziopikou, K.P. The PD-1/PD-L1 Axis in HER2+ Ductal Carcinoma In Situ (DCIS) of the Breast. Am. J. Clin. Pathol. 2019, 152, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Hendry, S.; Pang, J.-M.M.B.; Byrne, D.J.; Lakhani, S.R.; Cummings, M.C.; Campbell, I.G.; Mann, G.B.; Gorringe, K.L.; Fox, S.B.; BruceMann, G.; et al. Relationship of the Breast Ductal Carcinoma In Situ Immune Microenvironment with Clinicopathological and Genetic Features. Clin. Cancer Res. 2017, 23, 5210–5217. [Google Scholar] [CrossRef] [PubMed]
- Hoskoppal, D.; Reisenbichler, E.S. Can Tumor-Associated Macrophages in Ductal Carcinoma in Situ on Biopsy Predict Invasive Carcinoma on Excision? Hum. Pathol. 2018, 82, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Toss, M.S.; Miligy, I.; Al-Kawaz, A.; Alsleem, M.; Khout, H.; Rida, P.C.; Aneja, R.; Green, A.R.; Ellis, I.O.; Rakha, E.A. Prognostic Significance of Tumor-Infiltrating Lymphocytes in Ductal Carcinoma in Situ of the Breast. Mod. Pathol. 2018, 31, 1226–1236. [Google Scholar] [CrossRef]
- Semeraro, M.; Adam, J.; Stoll, G.; Louvet, E.; Chaba, K.; Poirier-Colame, V.; Sauvat, A.; Senovilla, L.; Vacchelli, E.; Bloy, N.; et al. The Ratio of CD8+/FOXP3 T Lymphocytes Infiltrating Breast Tissues Predicts the Relapse of Ductal Carcinoma in Situ. Oncoimmunology 2016, 5, e1218106. [Google Scholar] [CrossRef]
- Miligy, I.; Mohan, P.; Gaber, A.; Aleskandarany, M.A.; Nolan, C.C.; Diez-Rodriguez, M.; Mukherjee, A.; Chapman, C.; Ellis, I.O.; Green, A.R.; et al. Prognostic Significance of Tumour Infiltrating B Lymphocytes in Breast Ductal Carcinoma in Situ. Histopathology 2017, 71, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Thike, A.A.; Chen, X.; Koh, V.C.Y.; Binte Md Nasir, N.D.; Yeong, J.P.S.; Bay, B.H.; Tan, P.H. Higher Densities of Tumour-Infiltrating Lymphocytes and CD4+ T Cells Predict Recurrence and Progression of Ductal Carcinoma in Situ of the Breast. Histopathology 2020, 76, 852–864. [Google Scholar] [CrossRef]
- Morita, M.; Yamaguchi, R.; Tanaka, M.; Tse, G.M.; Yamaguchi, M.; Kanomata, N.; Naito, Y.; Akiba, J.; Hattori, S.; Minami, S.; et al. CD8+ Tumor- Infiltrating Lymphocytes Contribute to Spontaneous “Healing” in HER2- Positive Ductal Carcinoma in Situ. Cancer Med. 2016, 5, 1607–1618. [Google Scholar] [CrossRef]
- Lv, S.; Wang, S.; Qiao, G.; Wang, X.; Zhou, X.; Yan, F.; Li, Y.; Wang, S.; Morse, M.A.; Hobeika, A.; et al. Functional CD3+ CD8+ PD1− T Cell Accumulation and PD-L1 Expression Increases During Tumor Invasion in DCIS of the Breast. Clin. Breast Cancer 2019, 19, e617–e623. [Google Scholar] [CrossRef]
- Beguinot, M.; Dauplat, M.-M.; Kwiatkowski, F.; Lebouedec, G.; Tixier, L.; Pomel, C.; Penault-Llorca, F.; Radosevic-robin, N. Analysis of Tumour-Infiltrating Lymphocytes Reveals Two New Biologically Different Subgroups of Breast Ductal Carcinoma in Situ. BMC Cancer 2018, 18, 129. [Google Scholar] [CrossRef]
- Chen, X.-Y.Y.; Thike, A.A.; Koh, V.C.Y.; Nasir, N.D.M.; Bay, B.H.; Tan, P.H.; Cui, V.; Koh, Y.; Diyana, N.; Nasir, M.D.; et al. Breast Ductal Carcinoma in Situ Associated with Microinvasion Induces Immunological Response and Predicts Ipsilateral Invasive Recurrence. Virchows Arch. 2021, 478, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Perou, C.M.; Sùrlie, T.; Eisen, M.B.; Van De Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular Portraits of Human Breast Tumours. Nature 2000, 533, 747–752. [Google Scholar] [CrossRef]
- Simpson, P.T.; Reis-Filho, J.S.; Gale, T.; Lakhani, S.R. Molecular Evolution of Breast Cancer. J. Pathol. 2005, 205, 248–254. [Google Scholar] [CrossRef]
- Munkácsy, G.; Santarpia, L.; Győrffy, B. Gene Expression Profiling in Early Breast Cancer—Patient Stratification Based on Molecular and Tumor Microenvironment Features. Biomedicines 2022, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Toss, M.S.; Abidi, A.; Lesche, D.; Joseph, C.; Mahale, S.; Saunders, H.; Kader, T.; Miligy, I.M.; Green, A.R.; Gorringe, K.L.; et al. The Prognostic Significance of Immune Microenvironment in Breast Ductal Carcinoma in Situ. Br. J. Cancer 2020, 122, 1496–1506. [Google Scholar] [CrossRef]
- Chen, X.-Y.Y.; Thike, A.A.; Md Nasir, N.D.; Koh, V.C.Y.; Bay, B.H.; Tan, P.H. Higher Density of Stromal M2 Macrophages in Breast Ductal Carcinoma in Situ Predicts Recurrence. Virchows Arch. 2020, 476, 825–833. [Google Scholar] [CrossRef]
- Morita, M.; Yamaguchi, R.; Tanaka, M.; Tse, G.M.; Yamaguchi, M.; Otsuka, H.; Kanomata, N.; Minami, S.; Eguchi, S.; Yano, H. Two Progressive Pathways of Microinvasive Carcinoma: Low-Grade Luminal Pathway and High-Grade HER2 Pathway Based on High Tumour-Infiltrating Lymphocytes. J. Clin. Pathol. 2016, 69, 890–898. [Google Scholar] [CrossRef]
- Agahozo, M.C.; Westenend, P.J.; van Bockstal, M.R.; Hansum, T.; Giang, J.; Matlung, S.E.; van Deurzen, C.H.M. Immune Response and Stromal Changes in Ductal Carcinoma in Situ of the Breast Are Subtype Dependent. Mod. Pathol. 2020, 33, 1773–1782. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.J.; Baehner, F.; O’Meara, T.; Ojukwu, E.; Han, B.; Mukhtar, R.; Tandon, V.; Endicott, M.; Zhu, Z.; Wong, J.; et al. Characterizing the Immune Microenvironment in High-Risk Ductal Carcinoma in Situ of the Breast. Breast Cancer Res. Treat. 2018, 161, 17–28. [Google Scholar] [CrossRef]
- Tawara, K.; Scott, H.; Emathinger, J.; Ide, A.; Fox, R.; Greiner, D.; LaJoie, D.; Hedeen, D.; Nandakumar, M.; Oler, A.J.; et al. Co-Expression of VEGF and IL-6 Family Cytokines Is Associated with Decreased Survival in HER2 Negative Breast Cancer Patients: Subtype-Specific IL-6 Family Cytokine-Mediated VEGF Secretion. Transl. Oncol. 2019, 12, 245–255. [Google Scholar] [CrossRef]
- Liu, S.; Lee, J.S.; Jie, C.; Park, M.H.; Iwakura, Y.; Patel, Y.; Soni, M.; Reisman, D.; Chen, H. HER2 Overexpression Triggers an IL1α Proinflammatory Circuit to Drive Tumorigenesis and Promote Chemotherapy Resistance. Cancer Res. 2018, 78, 2040–2051. [Google Scholar] [CrossRef]
- Fridman, W.H.; Galon, J.; Pagès, F.; Tartour, E.; Sautès-Fridman, C.; Kroemer, G. Prognostic and Predictive Impact of Intra- and Peritumoral Immune Infiltrates. Cancer Res. 2011, 71, 5601–5605. [Google Scholar] [CrossRef]
- Fridman, W.H.; Pagès, F.; Sautès-Fridman, C.; Galon, J. The Immune Contexture in Human Tumours: Impact on Clinical Outcome. Nat. Rev. Cancer 2012, 12, 298–306. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Classification of Tumours Editorial Board WHO Classification of Breast Tumours; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Galon, J.; Fridman, W.H.; Pages, F. The Adaptive Immunologic Microenvironment in Colorectal Cancer: A Novel Perspective. Cancer Res. 2007, 67, 1883–1886. [Google Scholar] [CrossRef]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; Van Den Eynden, G.; Baehner, F.L.; Perez, E.A.; et al. In Breast Cancer: Recommendations by an International TILs Working Group. Ann. Oncol. 2014, 2018, 259–271. [Google Scholar] [CrossRef]
- Available online: Https://Www.Tilsinbreastcancer.Org/ (accessed on 21 March 2022).
- Yeong, J.; Chun, J.; Lim, T.; Lee, B.; Li, H.; Chong, C.; Ong, H.; Thike, A.A.; Yeap, W.H.; Yang, Y.; et al. Prognostic Value of CD8+ PD-1+ Immune Infiltrates and PDCD1 Gene Expression in Triple Negative Breast Cancer. J. Immunother. Cancer 2019, 7, 34. [Google Scholar] [CrossRef]
- Jin, Y.W.; Hu, P. Tumor-Infiltrating CD8 T Cells Predict Clinical Breast Cancer Outcomes in Young Women. Cancers 2020, 12, 1076. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Chung, Y.R.; Kim, H.J.; Woo, J.W.; Ahn, S.; Park, S.Y. Immune Microenvironment in Ductal Carcinoma in Situ: A Comparison with Invasive Carcinoma of the Breast. Breast Cancer Res. 2020, 22, 32. [Google Scholar] [CrossRef]
- Datta, J.; Rosemblit, C.; Berk, E.; Showalter, L.; Namjoshi, P.; Mick, R.; Lee, K.P.; Brod, A.M.; Yang, R.L.; Kelz, R.R.; et al. Progressive Loss of Anti-HER2 CD4+ T-Helper Type 1 Response in Breast Tumorigenesis and the Potential for Immune Restoration. Oncoimmunology 2015, 4, e1022301. [Google Scholar] [CrossRef]
- Egelston, C.A.; Avalos, C.; Tu, T.Y.; Rosario, A.; Wang, R.; Solomon, S.; Srinivasan, G.; Nelson, M.S.; Huang, Y.; Lim, M.H.; et al. Resident Memory CD8+ T Cells within Cancer Islands Mediate Survival in Breast Cancer Patients. JCI Insight 2019, 4, e130000. [Google Scholar] [CrossRef]
- Ostroumov, D.; Fekete-Drimusz, N.; Saborowski, M.; Kühnel, F.; Woller, N. CD4 and CD8 T Lymphocyte Interplay in Controlling Tumor Growth. Cell. Mol. Life Sci. 2018, 75, 689–713. [Google Scholar] [CrossRef]
- Caza, T.; Landas, S. Functional and Phenotypic Plasticity of CD4+ T Cell Subsets. Biomed Res. Int. 2015, 2015, 521957. [Google Scholar] [CrossRef] [PubMed]
- Beissert, S.; Schwarz, A.; Schwarz, T. Regulatory T Cells. J. Investig. Dermatol. 2006, 126, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Bettelli, E.; Carrier, Y.; Gao, W.; Korn, T.; Strom, T.B.; Oukka, M.; Weiner, H.L.; Kuchroo, V.K. Reciprocal Developmental Pathways for the Generation of Pathogenic Effector TH17 and Regulatory T Cells. Nature 2006, 441, 235–238. [Google Scholar] [CrossRef]
- Zhou, Y.; Shao, N.; Aierken, N.; Xie, C.; Ye, R.; Qian, X.; Hu, Z.; Zhang, J.; Lin, Y. Prognostic Value of Tumor-Infiltrating FoxP3+ Regulatory T Cells in Patients with Breast Cancer: A Meta-Analysis. J. Cancer 2017, 8, 4098–4105. [Google Scholar] [CrossRef]
- Syn, N.L.; Teng, M.W.L.; Mok, T.S.K.; Soo, R.A. De-Novo and Acquired Resistance to Immune Checkpoint Targeting. Lancet. Oncol. 2017, 18, e731–e741. [Google Scholar] [CrossRef]
- Yi, J.S.; Cox, M.A.; Zajac, A.J. T-Cell Exhaustion: Characteristics, Causes and Conversion. Immunology 2010, 129, 474–481. [Google Scholar] [CrossRef]
- Padmanabhan, R.; Kheraldine, H.S.; Meskin, N.; Vranic, S.; Al Moustafa, A.E. Crosstalk between HER2 and PD-1/PD-L1 in Breast Cancer: From Clinical Applications to Mathematical Models. Cancers 2020, 12, 636. [Google Scholar] [CrossRef] [PubMed]
- Lastwika, K.J.; Wilson, W.; Li, Q.K.; Norris, J.; Xu, H.; Ghazarian, S.R.; Kitagawa, H.; Kawabata, S.; Taube, J.M.; Yao, S.; et al. Control of PD-L1 Expression by Oncogenic Activation of the AKT-MTOR Pathway in Non-Small Cell Lung Cancer. Cancer Res. 2016, 76, 227–238. [Google Scholar] [CrossRef]
- Peng, S.; Wang, R.; Zhang, X.; Ma, Y.; Zhong, L.; Li, K.; Nishiyama, A.; Arai, S.; Yano, S.; Wang, W. EGFR-TKI Resistance Promotes Immune Escape in Lung Cancer via Increased PD-L1 Expression. Mol. Cancer 2019, 18, 165. [Google Scholar] [CrossRef]
- Suh, K.J.; Sung, J.H.; Kim, J.W.; Han, S.-H.; Lee, H.S.; Min, A.; Kang, M.H.; Kim, J.E.; Kim, J.-W.; Kim, S.H.; et al. EGFR or HER2 Inhibition Modulates the Tumor Microenvironment by Suppression of PD-L1 and Cytokines Release. Oncotarget 2017, 8, 63901–63910. [Google Scholar] [CrossRef] [PubMed]
- He, Q.-F.; Xu, Y.; Li, J.; Huang, Z.-M.; Li, X.-H.; Wang, X. CD8+ T-Cell Exhaustion in Cancer: Mechanisms and New Area for Cancer Immunotherapy. Brief. Funct. Genom. 2019, 18, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Muenst, S.; Schaerli, A.R.; Gao, F.; Däster, S.; Trella, E.; Droeser, R.A.; Muraro, M.G.; Zajac, P.; Zanetti, R.; Gillanders, W.E.; et al. The Presence of Programmed Death 1 (PD-1)-Positive Tumor- Infiltrating Lymphocytes Is Associated with Poor Prognosis in Human Breast Cancer. Breast Cancer Res. Treat. 2012, 76, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.Z.; Sarian, L.O.; Derchain, S.F.M.; Pinto, G.A.; Vassallo, J. Prognostic Significance of PD-L1 and PD-L2 in Breast Cancer. Hum. Pathol. 2016, 47, 78–84. [Google Scholar] [CrossRef]
- Andreatta, M.; Corria-Osorio, J.; Müller, S.; Cubas, R.; Coukos, G.; Carmona, S.J. Interpretation of T Cell States from Single-Cell Transcriptomics Data Using Reference Atlases. Nat. Commun. 2021, 12, 2965. [Google Scholar] [CrossRef]
- Cano, R.L.E.; Lopera, H.D.E. Introduction to T and B Lymphocytes; El Rosario University Press: Bogota, Colombia, 2013. [Google Scholar]
- Chen, Y.; Song, Y.; Du, W.; Gong, L.; Chang, H.; Zou, Z. Tumor-Associated Macrophages: An Accomplice in Solid Tumor Progression. J. Biomed. Sci. 2019, 26, 78. [Google Scholar] [CrossRef]
- Van Nguyen, A.; Pollard, J.W. Colony Stimulating Factor-1 Is Required to Recruit Macrophages into the Mammary Gland to Facilitate Mammary Ductal Outgrowth. Dev. Biol. 2002, 247, 11–25. [Google Scholar] [CrossRef]
- Gouon-Evans, V.; Rothenberg, M.E.; Pollard, J.W. Postnatal Mammary Gland Development Requires Macrophages and Eosinophils. Development 2000, 127, 2269–2282. [Google Scholar] [CrossRef]
- Bronte, V.; Brandau, S.; Chen, S.H.; Colombo, M.P.; Frey, A.B.; Greten, T.F.; Mandruzzato, S.; Murray, P.J.; Ochoa, A.; Ostrand-Rosenberg, S.; et al. Recommendations for Myeloid-Derived Suppressor Cell Nomenclature and Characterization Standards. Nat. Commun. 2016, 7, 12150. [Google Scholar] [CrossRef]
- Biswas, S.K.; Mantovani, A. Macrophage Plasticity and Interaction with Lymphocyte Subsets: Cancer as a Paradigm. Nat. Immunol. 2010, 11, 889–896. [Google Scholar] [CrossRef]
- Chan, T.; Pek, E.A.; Huth, K.; Ashkar, A.A. CD4+ T-Cells Are Important in Regulating Macrophage Polarization in C57BL/6 Wild-Type Mice. Cell. Immunol. 2011, 266, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Linde, N.; Casanova-Acebes, M.; Sosa, M.S.; Mortha, A.; Rahman, A.; Farias, E.; Harper, K.; Tardio, E.; Reyes Torres, I.; Jones, J.; et al. Macrophages Orchestrate Breast Cancer Early Dissemination and Metastasis. Nat. Commun. 2018, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Triulzi, T.; Forte, L.; Regondi, V.; Di Modica, M.; Ghirelli, C.; Carcangiu, M.L.; Sfondrini, L.; Balsari, A.; Tagliabue, E. HER2 Signaling Regulates the Tumor Immune Microenvironment and Trastuzumab Efficacy. Oncoimmunology 2019, 8, e1512942. [Google Scholar] [CrossRef]
- Meng, S.; Tripathy, D.; Frenkel, E.P.; Shete, S.; Naftalis, E.Z.; Huth, J.F.; Beitsch, P.D.; Leitch, M.; Hoover, S.; Euhus, D.; et al. Circulating Tumor Cells in Patients with Breast Cancer Dormancy. Clin. Cancer Res. 2004, 10, 8152–8162. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.U.; Cho, S.Y.; Jeong, H.; Han, J.; Chae, H.Y.; Yang, H.; Sung, C.O.; Choi, Y.-L.; Shin, Y.K.; Kwon, M.J. Matrix Metalloproteinase 11 (MMP11) in Macrophages Promotes the Migration of HER2-Positive Breast Cancer Cells and Monocyte Recruitment through CCL2-CCR2 Signaling. Lab. Investig. 2021, 102, 376–390. [Google Scholar] [CrossRef]
- Castiello, L.; Sestili, P.; Schiavoni, G.; Dattilo, R.; Monque, D.M.; Ciaffoni, F.; Iezzi, M.; Lamolinara, A.; Sistigu, A.; Moschella, F.; et al. Disruption of IFN-I Signaling Promotes HER2/Neu Tumor Progression and Breast Cancer Stem Cells. Cancer Immunol. Res. 2018, 6, 658–670. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solek, J.; Chrzanowski, J.; Cieslak, A.; Zielinska, A.; Piasecka, D.; Braun, M.; Sadej, R.; Romanska, H.M. Subtype-Specific Tumour Immune Microenvironment in Risk of Recurrence of Ductal Carcinoma In Situ: Prognostic Value of HER2. Biomedicines 2022, 10, 1061. https://doi.org/10.3390/biomedicines10051061
Solek J, Chrzanowski J, Cieslak A, Zielinska A, Piasecka D, Braun M, Sadej R, Romanska HM. Subtype-Specific Tumour Immune Microenvironment in Risk of Recurrence of Ductal Carcinoma In Situ: Prognostic Value of HER2. Biomedicines. 2022; 10(5):1061. https://doi.org/10.3390/biomedicines10051061
Chicago/Turabian StyleSolek, Julia, Jedrzej Chrzanowski, Adrianna Cieslak, Aleksandra Zielinska, Dominika Piasecka, Marcin Braun, Rafal Sadej, and Hanna M. Romanska. 2022. "Subtype-Specific Tumour Immune Microenvironment in Risk of Recurrence of Ductal Carcinoma In Situ: Prognostic Value of HER2" Biomedicines 10, no. 5: 1061. https://doi.org/10.3390/biomedicines10051061