
Carbonic Anhydrase VIII (CAVIII) Gene Mediated Colorectal Cancer Growth and Angiogenesis through Mediated miRNA 16-5p

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
2.3. Construction of Stable Expression CA8 shRNA Cell Line
2.4. Transient Transfection
2.5. Conditioned Medium (CM) Preparation
2.6. ELISA Assay
2.7. Migration Assay
2.8. Tube Formation Assay
2.9. Immunohistochemical (IHC) Staining
2.10. Western Blot Analysis
2.11. Quantitative Real-Time PCR
2.12. In Vivo Tumor Angiogenesis Assay
2.13. Mice Xenograft Assay
2.14. Hemoglobin Assay
2.15. Statistical Analysis
3. Results
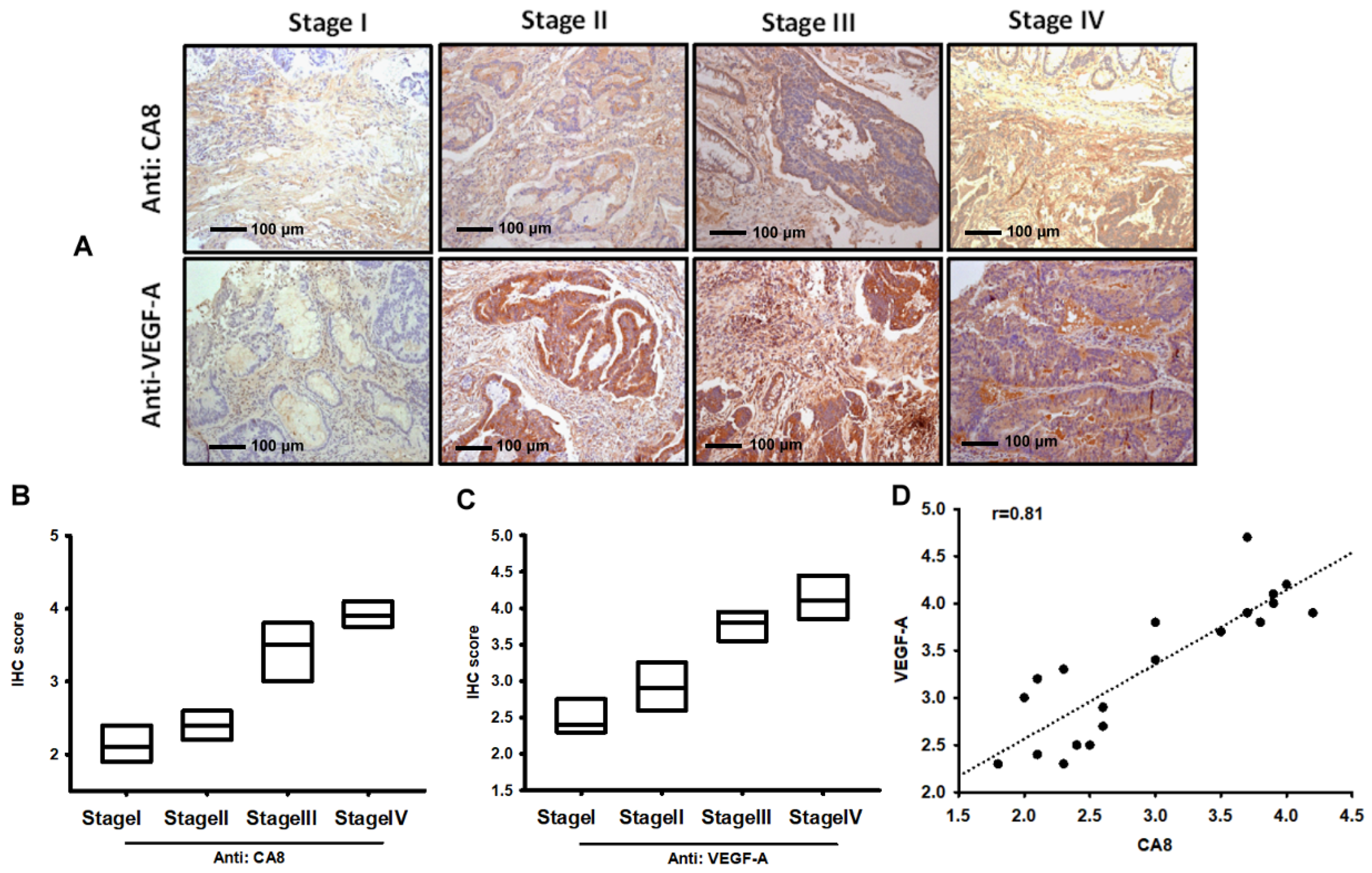
3.1. Identification of CAVIII and VEGF-A Expression in Colorectal Cancer Tissues
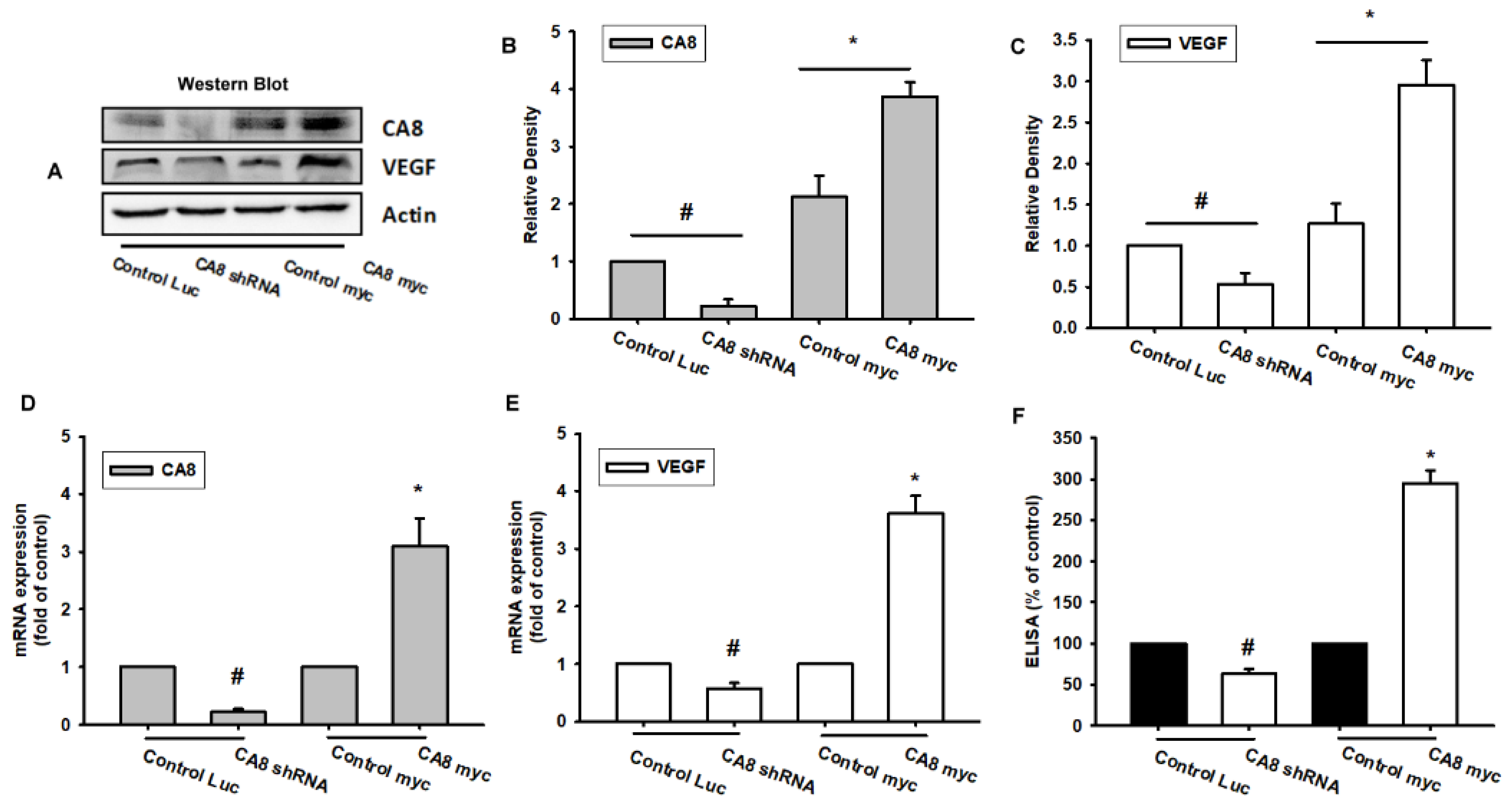
3.2. Knockdown of CAVIII or Over-Expression of CAVIII Directly Regulates VEGF-A Expression in Human Colon Cancer Cell Lines
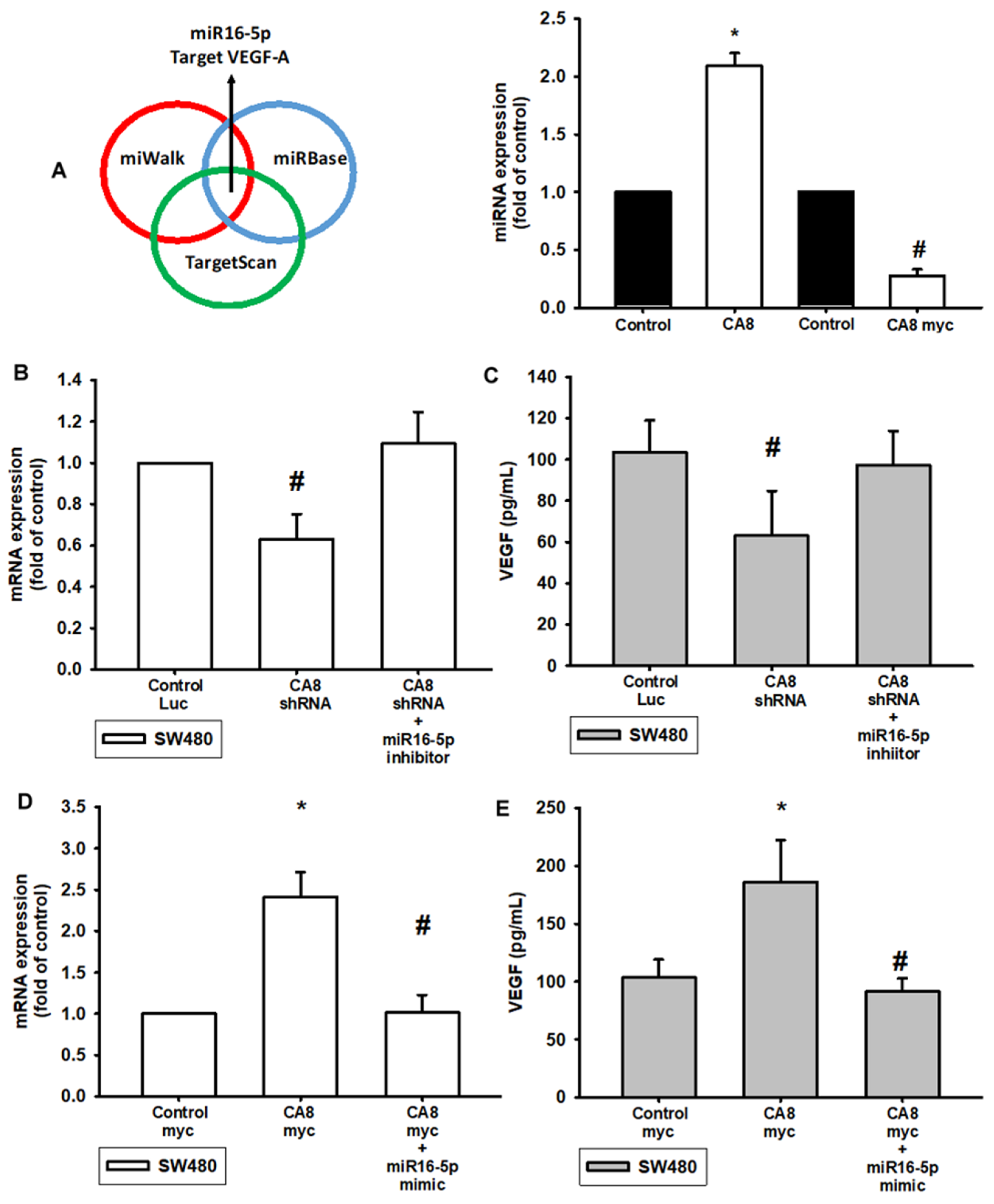
3.3. Knockdown of CA8 Over-Expression of CAVIII Directly Regulates VEGF-A Expression through Regulated by miR16-5p
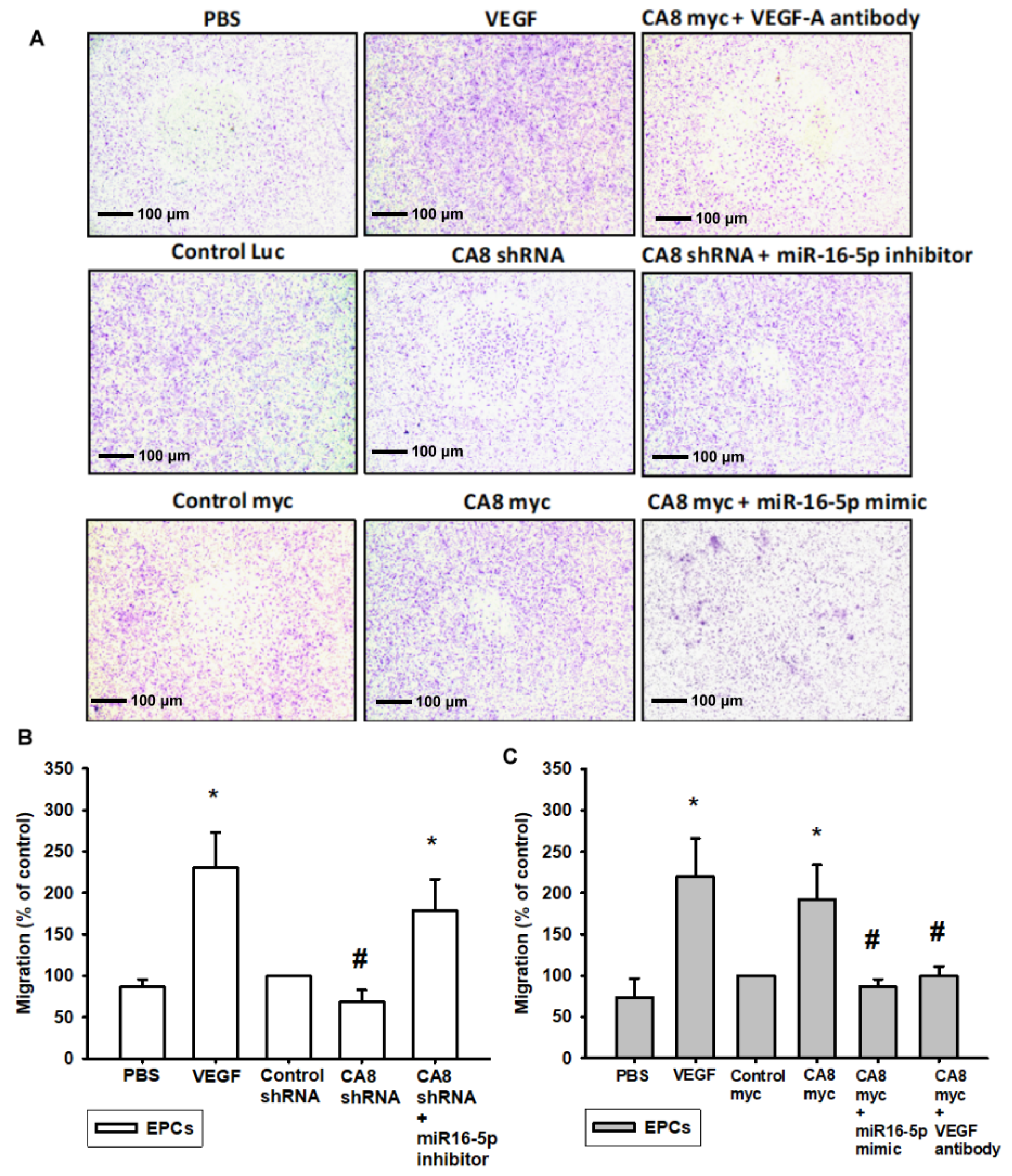
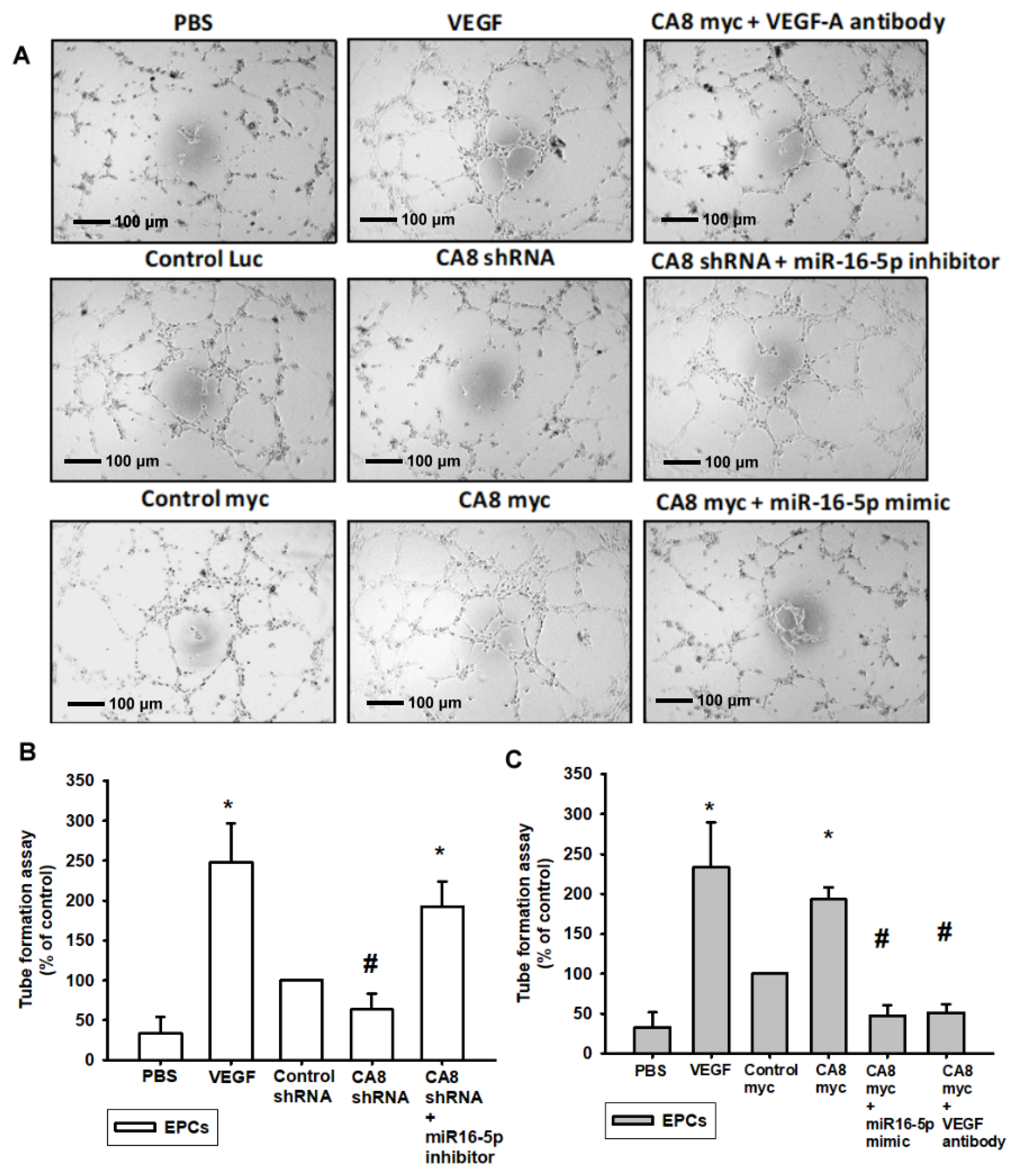
3.4. CAVIII Influences VEGFA Production and Affects In Vitro Angiogenesis by Regulating miR-16-5p
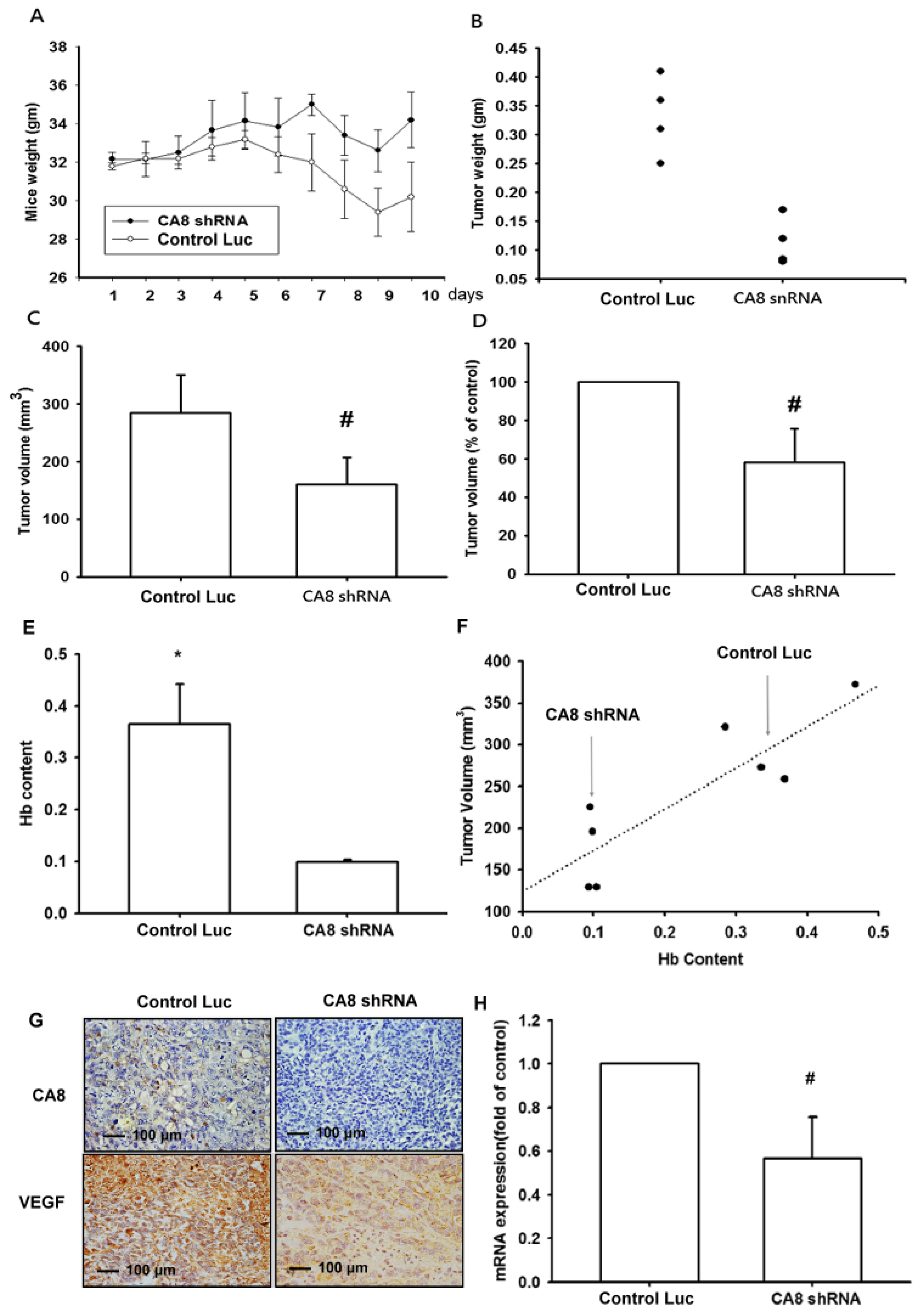
3.5. Inhibition of Tumor Growth and Angiogenesis by CAVIII Expression in a Mouse Xenograft Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hung, Y.H.; Chung, Y.C.; Lee, P.Y.; Kao, H.Y. Exploration of Preventable Hospitalizations for Colorectal Cancer with the National Cancer Control Program in Taiwan. Int. J. Environ. Res. Public Health 2021, 18, 9327. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Chen, T.H.; Wu, Y.C.; Fang, C.Y.; Wang, J.Y.; Chen, C.P.; Huang, K.W.; Jiang, J.K. Taiwan Society of Colon and Rectal Surgeons (TSCRS) Consensus for Cytoreduction Selection in Metastatic Colorectal Cancer. Ann. Surg. Oncol. 2021, 28, 1762–1776. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Xiang, R.; Ren, J.; Song, W.; Lu, W.; Fu, T. A Nomogram for Predicting Multiple Metastases in Metastatic Colorectal Cancer Patients: A Large Population-Based Study. Front. Oncol. 2021, 11, 633995. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, Y.; Kopetz, S.; Panettieri, E.; Hwang, H.; Wang, X.; Cao, H.S.T.; Tzeng, C.D.; Chun, Y.S.; Aloia, T.A.; Vauthey, J.N. Improved Survival over Time After Resection of Colorectal Liver Metastases and Clinical Impact of Multigene Alteration Testing in Patients with Metastatic Colorectal Cancer. J. Gastrointest. Surg. 2022, 26, 583–593. [Google Scholar] [CrossRef]
- Lakkis, M.M.; Bergenhem, N.C.; O’Shea, K.S.; Tashian, R.E. Expression of the acatalytic VIII gene, Car8, during mouse embryonic development. Histochem. J. 1997, 29, 135–141. [Google Scholar] [CrossRef]
- Chi, T.H.; Hsieh, B.Y.; Liang, P.S.; Han, T.H.; Hsieh, M. Expression and Functional Study of Single Mutations of 8 in Neuronal Cells. Cell Mol. Neurobiol. 2021, 41, 1355–1371. [Google Scholar] [CrossRef]
- Lo, C.M.; Ma, Y.S.; Wei, Y.H.; Hsieh, B.Y.T.; Hsieh, M. Promoter analysis and transcriptional regulation of human VIII gene in a MERRF disease cell model. Arch. Biochem. Biophys. 2018, 641, 50–61. [Google Scholar] [CrossRef]
- Ishihara, T.; Takeuchi, T.; Nishimori, I.; Adachi, Y.; Minakuchi, T.; Fujita, J.; Sonobe, H.; Ohtsuki, Y.; Onishi, S. Carbonic anhydrase-related protein VIII increases invasiveness of non-small cell lung adenocarcinoma. Virchows. Arch. 2006, 448, 830–837. [Google Scholar] [CrossRef]
- Miyaji, E.; Nishimori, I.; Taniuchi, K.; Takeuchi, T.; Ohtsuki, Y.; Onishi, S. Overexpression of -related protein VIII in human colorectal cancer. J. Pathol. 2003, 201, 37–45. [Google Scholar] [CrossRef]
- Nishikata, M.; Nishimori, I.; Taniuchi, K.; Takeuchi, T.; Minakuchi, T.; Kohsaki, T.; Adachi, Y.; Ohtsuki, Y.; Onishi, S. Carbonic anhydrase-related protein VIII promotes colon cancer cell growth. Mol. Carcinog. 2007, 46, 208–214. [Google Scholar] [CrossRef]
- Ma, H.L.; Yu, S.J.; Chen, J.; Ding, X.F.; Chen, G.; Liang, Y.; Pan, J.L. CA8 promotes RCC proliferation and migration though its expression level is lower in tumor compared to adjacent normal tissue. Biomed. Pharmacother. 2020, 121, 109578. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cho, Y.B.; Hong, H.K.; Wu, S.; Ebert, P.J.; Bray, S.M.; Wong, S.S.; Ting, J.C.; Calley, J.N.; Whittington, C.F.; et al. Molecular dissection of CRC primary tumors and their matched liver metastases reveals critical role of immune microenvironment, EMT and angiogenesis in cancer metastasis. Sci. Rep. 2020, 10, 10725. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.F.; Qvortrup, C.; Pfeiffer, P. Angiogenesis Inhibitors for Colorectal Cancer. A Review of the Clinical Data. Cancers 2021, 13, 1031. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cai, B.; Shen, L.; Dong, Y.; Lu, Q.; Sun, S.; Liu, S.; Ma, S.; Ma, P.X.; Chen, J. MiRNA-29b suppresses tumor growth through simultaneously inhibiting angiogenesis and tumorigenesis by targeting Akt3. Cancer Lett. 2017, 397, 111–119. [Google Scholar] [CrossRef]
- Qun, L.; Wenda, X.; Weihong, S.; Jianyang, M.; Wei, C.; Fangzhou, L.; Zhenyao, X.; Pingjin, G. miRNA-27b modulates endothelial cell angiogenesis by directly targeting Naa15 in atherogenesis. Atherosclerosis 2016, 254, 184–192. [Google Scholar] [CrossRef]
- Joladarashi, D.; Krishnamurthy, P. Assessment of MiRNA Regulation of Endothelial Progenitor Cell Mediated Angiogenesis. Methods Mol. Biol. 2017, 1553, 305–314. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, X.J. Circ0021205 aggravates the progression of non-small cell lung cancer by targeting miRNA-16-5p/VEGFA. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 3148. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Zheng, Z.; Sun, J.; Zhang, Y.; Wei, C.; Ke, X.; Liu, Y.; Deng, L.; Wang, H. MiR-16-5p mediates a positive feedback loop in EV71-induced apoptosis and suppresses virus replication. Sci. Rep. 2017, 7, 16422. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Dong, X.; Wu, D.; Zhao, H.; Xu, C.; Li, H. Long noncoding RNA small nucleolar RNA host gene 12 promotes papillary thyroid carcinoma cell growth and invasion by targeting miR-16-5p. Histol. Histopathol. 2020, 35, 217–224. [Google Scholar] [CrossRef]
- Wang, F.; Wang, W.; Lu, L.; Xie, Y.; Yan, J.; Chen, Y.; Di, C.; Gan, L.; Si, J.; Zhang, H.; et al. MicroRNA165p regulates cell survival, cell cycle and apoptosis by targeting AKT3 in prostate cancer cells. Oncol. Rep. 2020, 44, 1282–1292. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Liu, S.C.; Chung, W.H.; Wang, S.W.; Wu, M.H.; Tang, C.H. Visfatin Increases VEGF-dependent Angiogenesis of Endothelial Progenitor Cells during Osteoarthritis Progression. Cells 2020, 9, 1315. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Chung, C.H.; Lu, Y.C.; Wu, M.H.; Chou, P.H.; Yen, J.Y.; Lai, Y.W.; Wang, G.S.; Liu, S.C.; Cheng, J.K.; et al. BMP-2 induces angiogenesis by provoking integrin alpha6 expression in human endothelial progenitor cells. Biochem. Pharmacol. 2018, 150, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.K.; Lin, Y.M.; Lo, C.M.; Tang, C.H.; Teng, C.L.; Chao, W.T.; Wu, M.H.; Liu, C.S.; Hsieh, M. Oncogenic roles of 8 in human osteosarcoma cells. Tumour. Biol. 2016, 37, 7989–8005. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, H.E.; Tang, C.H.; Tsai, C.H.; Chiu, C.H.; Wu, M.H.; Yen, Y. ET-1 Promotes Epithelial-Mesenchymal Transition in Oral Squamous Cell Carcinoma Cells via the microRNA-489-3p /TWIST Axis. Oncol. Targets Ther. 2021, 14, 5005–5018. [Google Scholar] [CrossRef]
- Wu, M.H.; Huang, C.Y.; Lin, J.A.; Wang, S.W.; Peng, C.Y.; Cheng, H.C.; Tang, C.H. Endothelin-1 promotes vascular endothelial growth factor-dependent angiogenesis in human chondrosarcoma cells. Oncogene 2014, 33, 1725–1735. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Wang, Y.; Yuan, M.; Zhao, Q.; Zhang, Y.; Yao, Y.; Duan, Y. Angiogenesis, Anti-Tumor, and Anti-Metastatic Activity of Novel alpha-Substituted Hetero-Aromatic Chalcone Hybrids as Inhibitors of Microtubule Polymerization. Front. Chem. 2021, 9, 766201. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, J.; Shen, L. Extracellular vesicle-mediated regulation of tumor angiogenesis- implications for anti-angiogenesis therapy. J. Cell Mol. Med. 2021, 25, 2776–2785. [Google Scholar] [CrossRef]
- Wang, K.; Chen, Q.; Liu, N.; Zhang, J.; Pan, X. Recent advances in, and challenges of, anti-angiogenesis agents for tumor chemotherapy based on vascular normalization. Drug Discov. Today 2021, 26, 2743–2753. [Google Scholar] [CrossRef]
- Lai, V.; Neshat, S.Y.; Rakoski, A.; Pitingolo, J.; Doloff, J.C. Drug delivery strategies in maximizing anti-angiogenesis and anti-tumor immunity. Adv. Drug Deliv. Rev. 2021, 179, 113920. [Google Scholar] [CrossRef]
- Diaz-Rubio, E.; Schmoll, H.J. Introduction. Critical role of anti-angiogenesis and VEGF inhibition in colorectal cancer. Oncology 2005, 69 (Suppl. S3), 1–3. [Google Scholar] [CrossRef] [PubMed]
- Ciombor, K.K.; Goldberg, R.M. Update on Anti-Angiogenesis Therapy in Colorectal Cancer. Curr. Colorectal. Cancer Rep. 2015, 11, 378–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winum, J.Y.; Innocenti, A.; Nasr, J.; Montero, J.L.; Scozzafava, A.; Vullo, D.; Supuran, C.T. Carbonic anhydrase inhibitors: Synthesis and inhibition of cytosolic/tumor-associated isozymes I, II, IX, and XII with N-hydroxysulfamides—A new zinc-binding function in the design of inhibitors. Bioorg. Med. Chem. Lett. 2005, 15, 2353–2358. [Google Scholar] [CrossRef]
- Queen, A.; Bhutto, H.N.; Yousuf, M.; Syed, M.A.; Hassan, M.I. IX: A tumor acidification switch in heterogeneity and chemokine regulation. Semin. Cancer Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Yamali, C.; Inci Gul, H.; Ozli, G.; Angeli, A.; Ballar Kirmizibayrak, P.; Erbaykent Tepedelen, B.; Sakagami, H.; Bua, S.; Supuran, C.T. Exploring of tumor-associated isoenzyme IX and XII inhibitory effects and cytotoxicities of the novel N-aryl-1-(4-sulfamoylphenyl)-5-(thiophen-2-yl)-1H-pyrazole-3-carboxamides. Bioorg. Chem. 2021, 115, 105194. [Google Scholar] [CrossRef] [PubMed]
- Pickering, L.M.; Larkin, J. Kidney cancer: IX in resected clear cell RCC. Nat. Rev. Urol. 2015, 12, 309–310. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Tang, C.H.; Chie, M.J.; Tsai, C.H.; Fong, Y.C.; Lu, Y.C.; Chen, W.C.; Lai, C.T.; Wei, C.Y.; Tai, H.C.; et al. Resistin facilitates VEGF-A-dependent angiogenesis by inhibiting miR-16-5p in human chondrosarcoma cells. Cell Death Dis. 2019, 10, 31. [Google Scholar] [CrossRef]
- Annese, T.; Tamma, R.; De Giorgis, M.; Ribatti, D. microRNAs Biogenesis, Functions and Role in Tumor Angiogenesis. Front. Oncol. 2020, 10, 581007. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, M.; Huang, P.-J.; Chou, P.-Y.; Wang, S.-W.; Lu, H.-C.; Su, W.-W.; Chung, Y.-C.; Wu, M.-H. Carbonic Anhydrase VIII (CAVIII) Gene Mediated Colorectal Cancer Growth and Angiogenesis through Mediated miRNA 16-5p. Biomedicines 2022, 10, 1030. https://doi.org/10.3390/biomedicines10051030
Hsieh M, Huang P-J, Chou P-Y, Wang S-W, Lu H-C, Su W-W, Chung Y-C, Wu M-H. Carbonic Anhydrase VIII (CAVIII) Gene Mediated Colorectal Cancer Growth and Angiogenesis through Mediated miRNA 16-5p. Biomedicines. 2022; 10(5):1030. https://doi.org/10.3390/biomedicines10051030
Chicago/Turabian StyleHsieh, Mingli, Pei-Ju Huang, Pei-Yu Chou, Shih-Wei Wang, Hsi-Chi Lu, Wei-Wen Su, Yuan-Chiang Chung, and Min-Huan Wu. 2022. "Carbonic Anhydrase VIII (CAVIII) Gene Mediated Colorectal Cancer Growth and Angiogenesis through Mediated miRNA 16-5p" Biomedicines 10, no. 5: 1030. https://doi.org/10.3390/biomedicines10051030