
Isoliquiritigenin Inhibits Gastric Cancer Stemness, Modulates Tumor Microenvironment, and Suppresses Tumor Growth through Glucose-Regulated Protein 78 Downregulation

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients Cohort
2.2. Cell Culture and Reagent
2.3. Cell Viability Analysis
2.4. Western Blot
2.5. Flow Cytometry
2.6. Sphere Formation Analysis
2.7. Soft Agar Colony Formation Analysis
2.8. Measurement of Tumor Growth Factor-β1 Level
2.9. Measurement of Interleukin-6 Level
2.10. Xenograft Tumor Experiments
2.11. Immunohistochemistry and Hematoxylin plus Eosin Staining
2.12. Statistical Analysis
3. Results
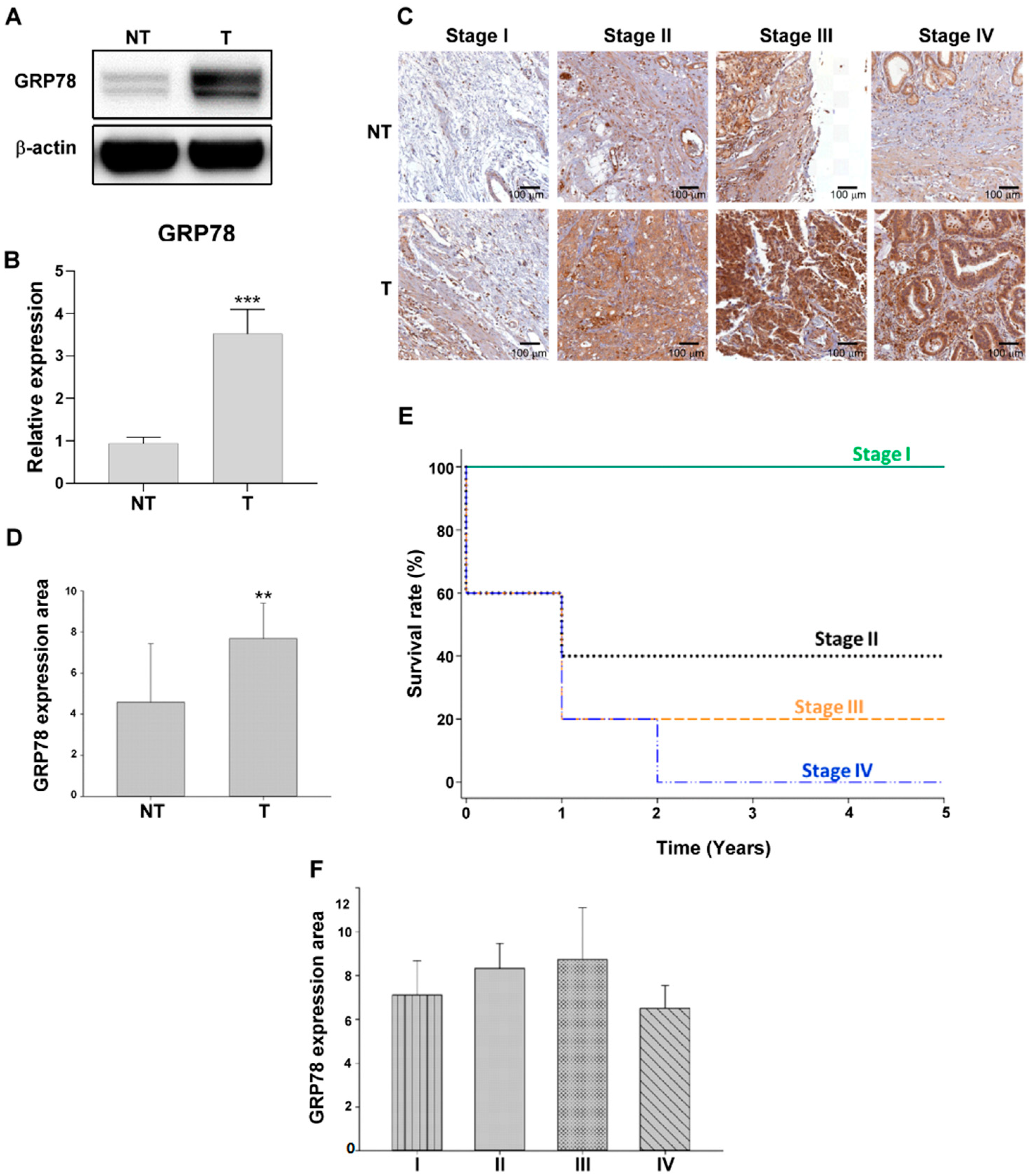
3.1. GRP78 Overexpression in Human Gastric Tumor Cells
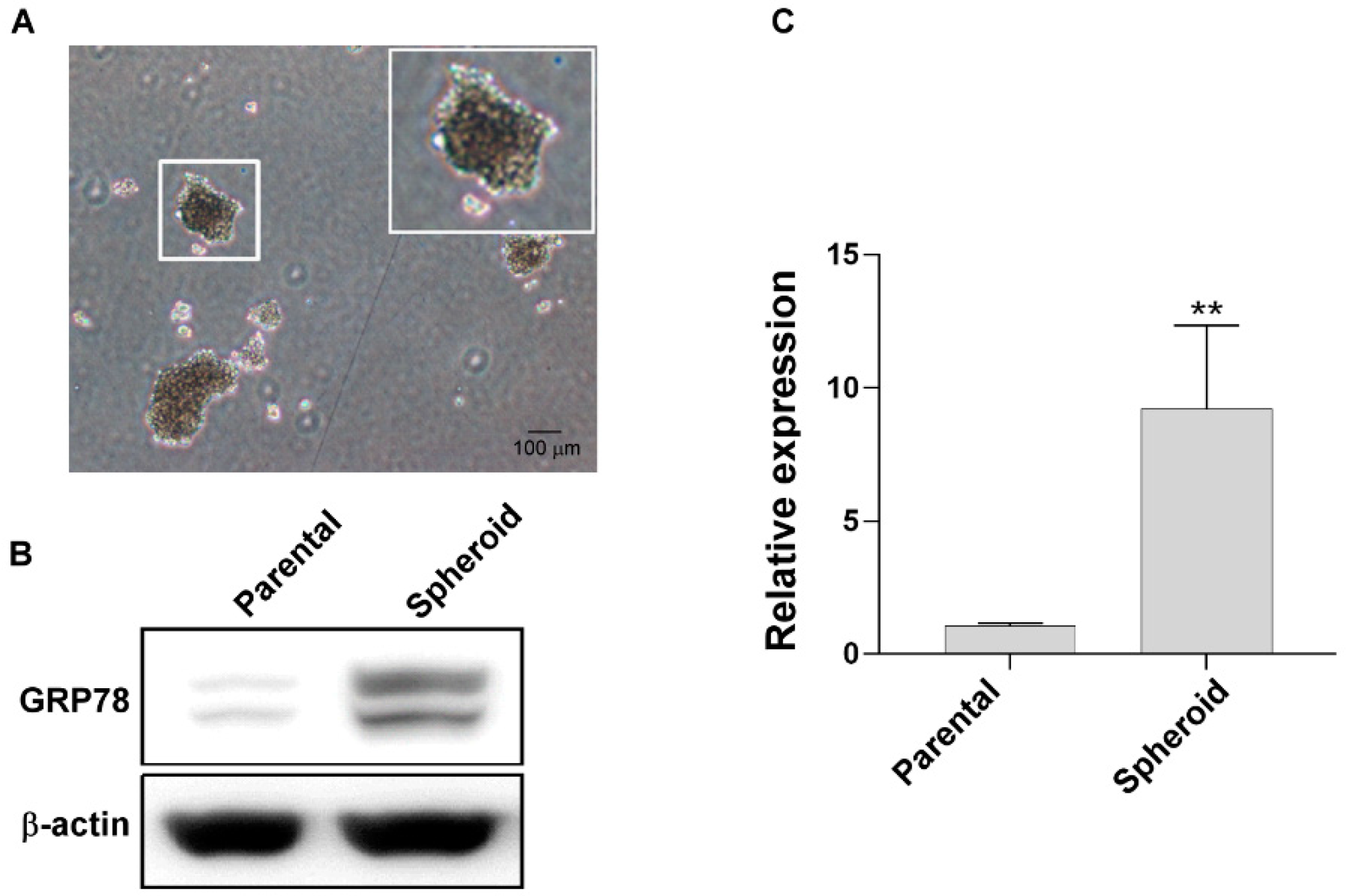
3.2. GRP78 Overexpression in Stem Cell-like Spheroid-Forming Human Gastric Cancer Cells
3.3. GRP78 Overexpression in Human Gastric Cancer Cells Promote Stem Cell-like Characteristics
3.4. ISL Inhibited GRP78 Expression and Suppressed Stem Cell-like Characteristics in Human Gastric Cancer Cells
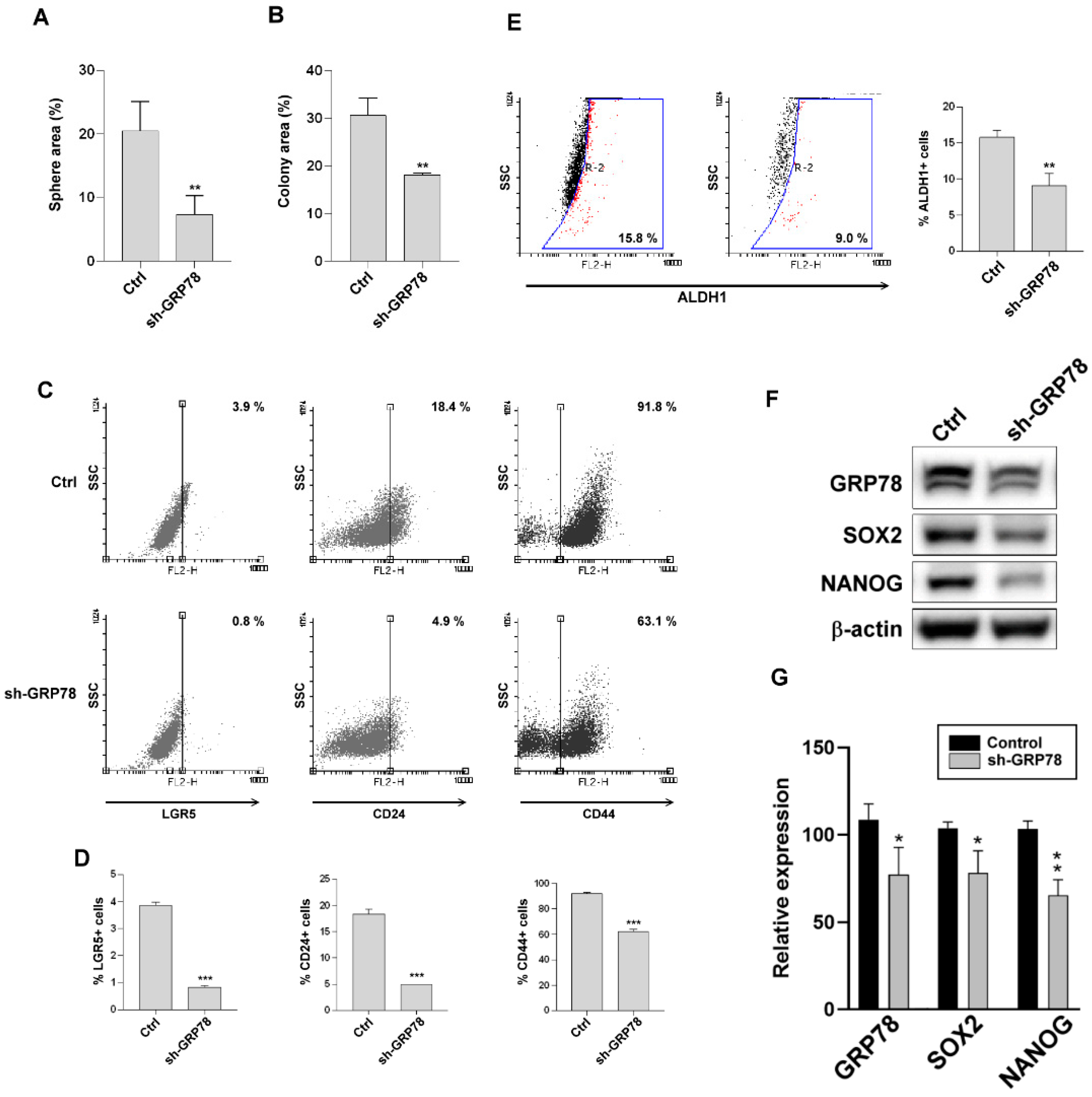
3.5. Knockdown of GRP78 Suppressed Stem Cell-like Characteristics in Human Gastric Cancer Cells
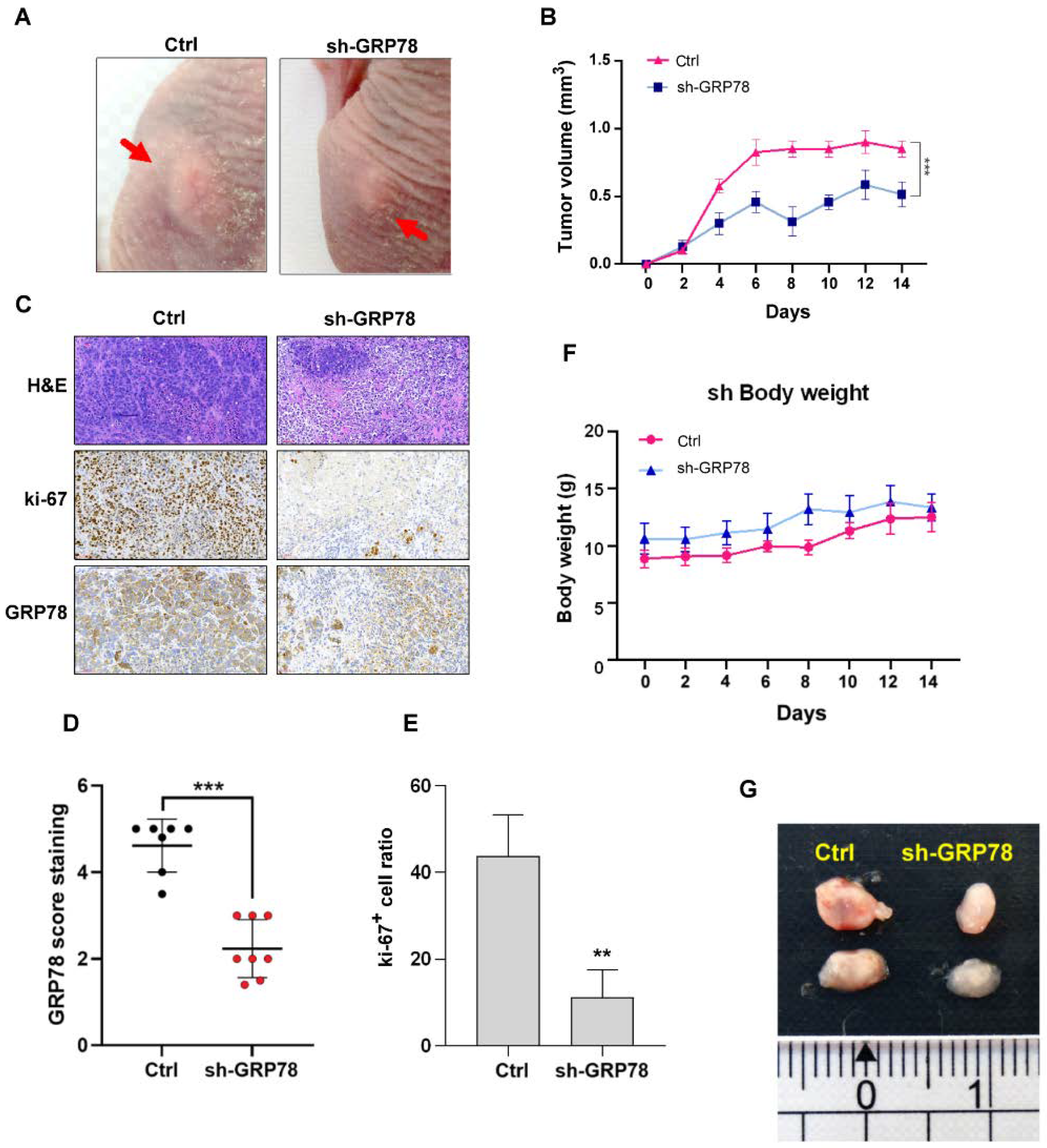
3.6. Knockdown of GRP78 Inhibited Tumor Growth in Xenograft Tumor Mice
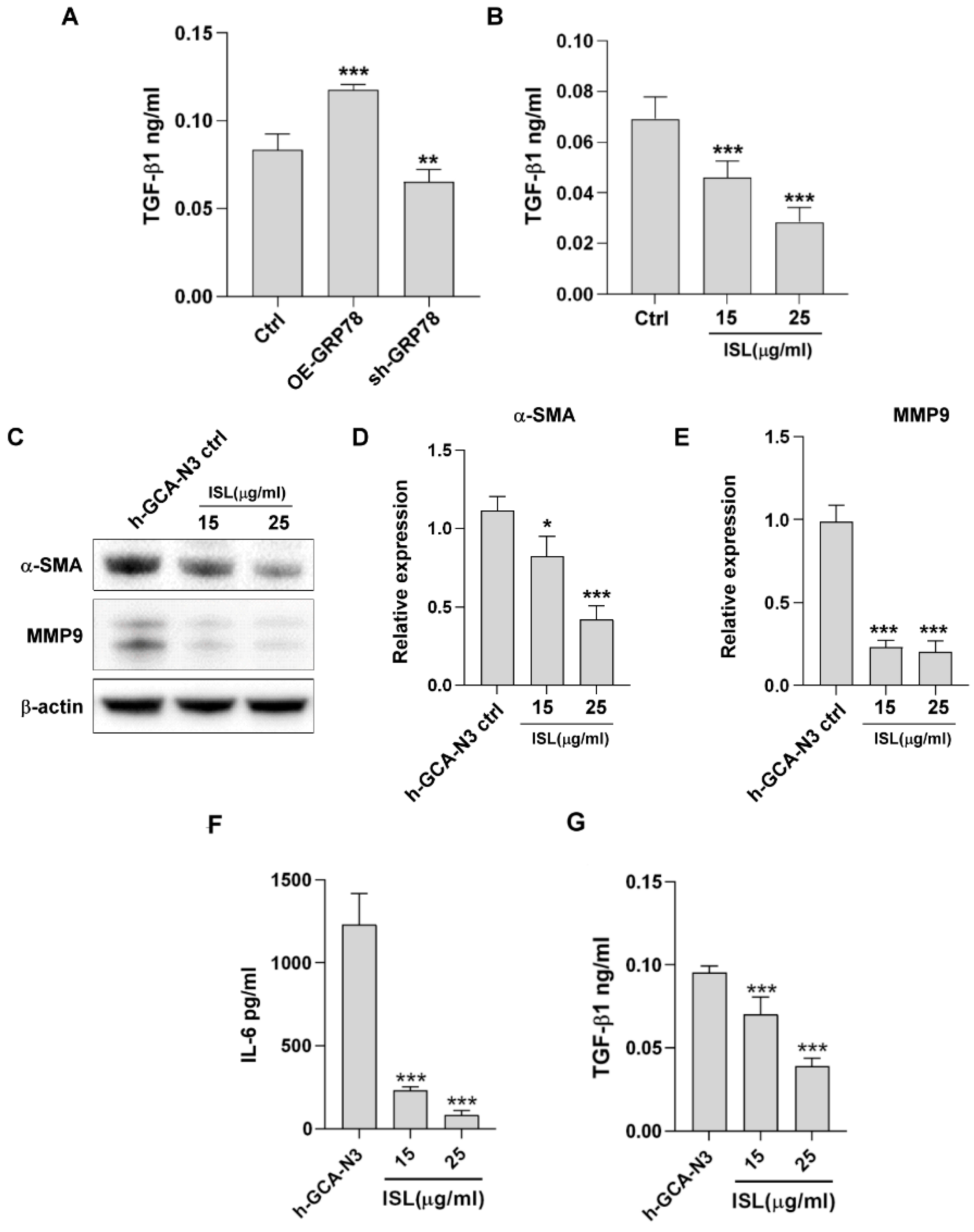
3.7. ISL Regulated the Tumor Microenvironment through GRP78-Mediated TGF-β1 Expression
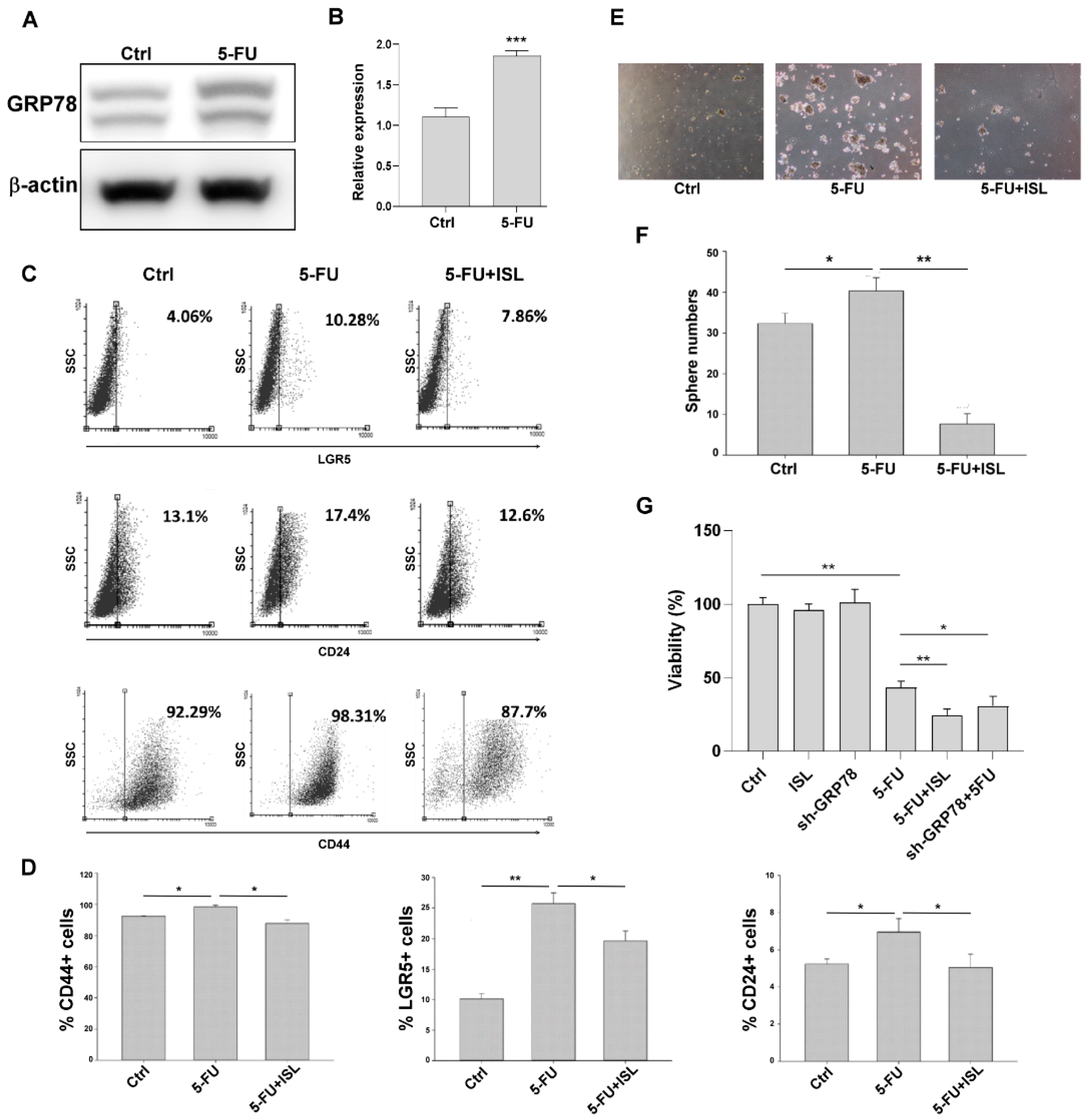
3.8. ISL Inhibited 5-FU-Induced GRP78-Mediated Gastric Cancer Stemness
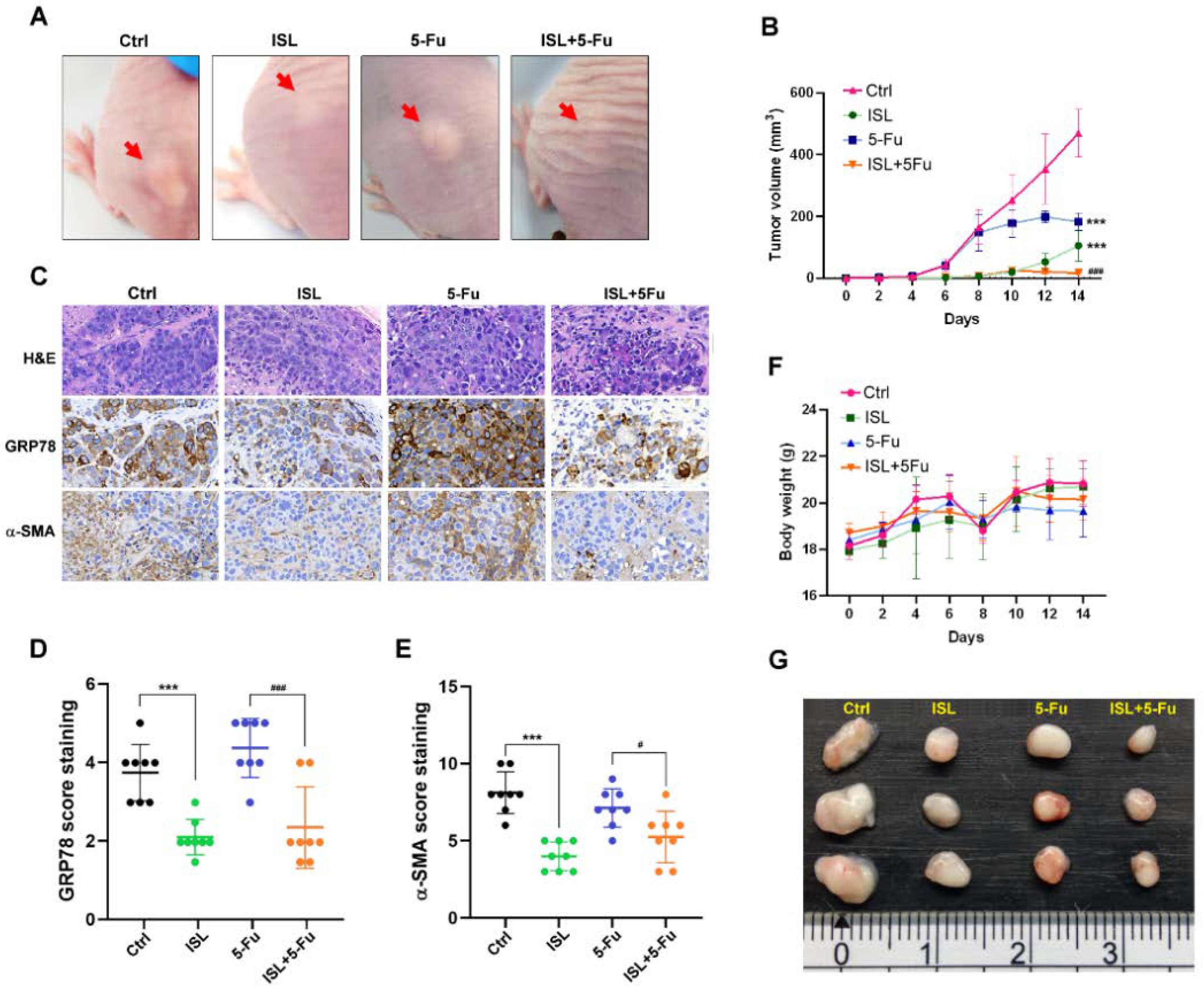
3.9. ISL Enhanced Chemosensitivity to 5-FU and Inhibited Tumor Growth in Xenograft Tumor Mice
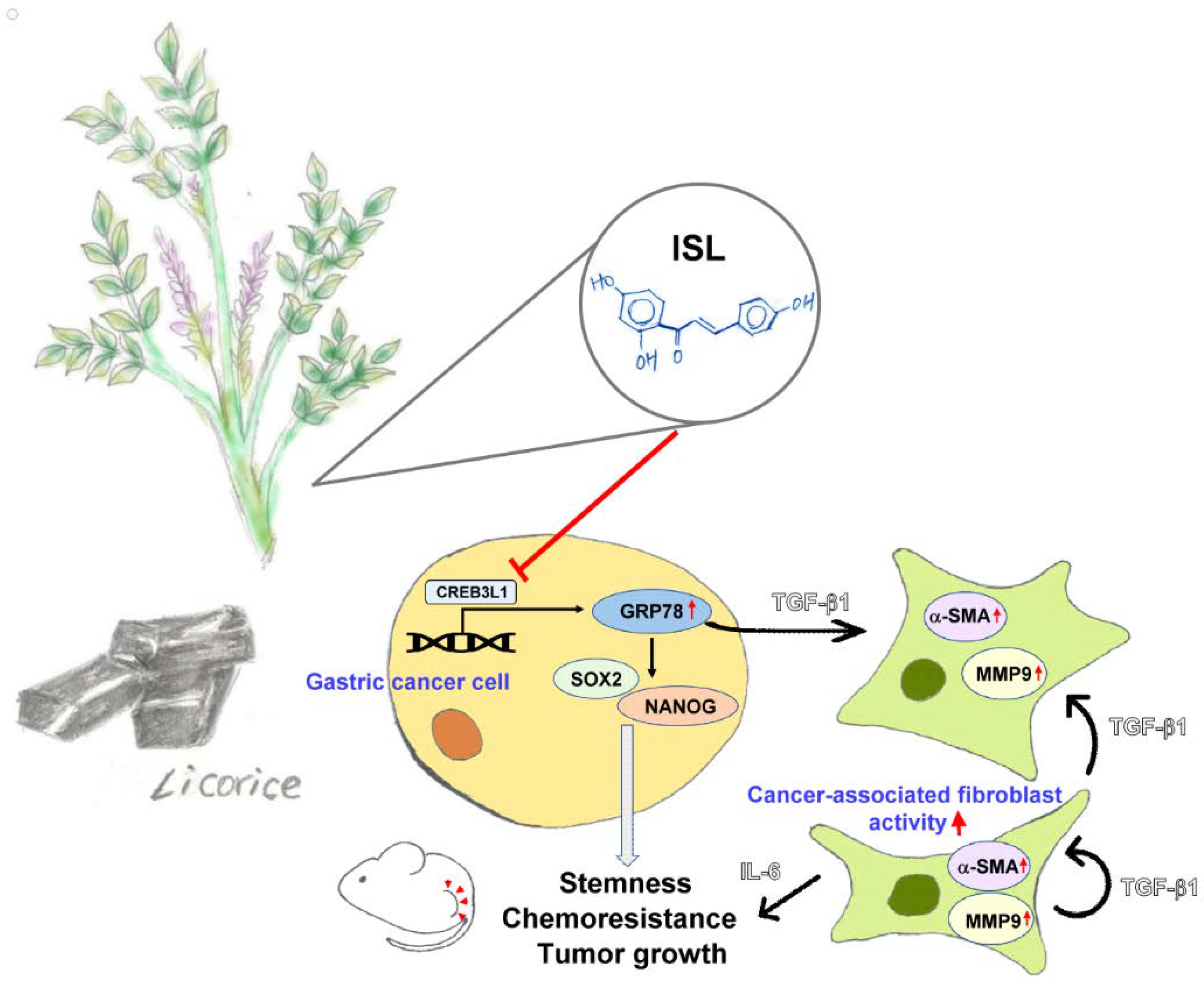
3.10. ISL Suppresses Cancer Stemness–Mediated Chemoresistance, Tumor Microenvironment, and Tumor Growth by GRP78 Inhibition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLO-BOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Otaegi-Ugartemendia, M.; Matheu, A.; Carrasco-Garcia, E. Impact of Cancer Stem Cells on Therapy Resistance in Gastric Cancer. Cancers 2022, 14, 1457. [Google Scholar] [CrossRef]
- Kharkar, P.S. Cancer Stem Cell (CSC) Inhibitors in Oncology—A Promise for a Better Therapeutic Outcome: State of the Art and Future Perspectives. J. Med. Chem. 2020, 63, 15279–15307. [Google Scholar] [CrossRef]
- Luo, B.; Lee, A.S. The critical roles of endoplasmic reticulum chaperones and unfolded protein response in tumorigenesis and anticancer therapies. Oncogene 2013, 32, 805–818. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Lee, A.S. Role of the unfolded protein response, GRP78 and GRP94 in organ homeostasis. J Cell Physiol. 2015, 230, 1413–1420. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.-Y.; Cheng, C.-C.; Wang, J.-Y.; Wu, D.-C.; Hsieh, J.-S.; Lee, S.-C.; Wang, W.-M. Discovery of Tumor Markers for Gastric Cancer by Proteomics. PLoS ONE 2014, 9, e84158. [Google Scholar] [CrossRef]
- Zheng, Y.-Z.; Cao, Z.-G.; Hu, X.; Shao, Z.-M. The endoplasmic reticulum stress markers GRP78 and CHOP predict disease-free survival and responsiveness to chemotherapy in breast cancer. Breast Cancer Res. Treat. 2014, 145, 349–358. [Google Scholar] [CrossRef]
- Kwon, D.; Koh, J.; Kim, S.; Go, H.; Min, H.S.; Kim, Y.A.; Kim, D.K.; Jeon, Y.K.; Chung, D.H. Overexpression of endoplasmic reticu-lum stress-related proteins, XBP1s and GRP78, predicts poor prognosis in pulmonary adenocarcinoma. Lung Cancer 2018, 122, 131–137. [Google Scholar] [CrossRef]
- Ninkovic, S.; Harrison, S.J.; Quach, H. Glucose-regulated protein 78 (GRP78) as a potential novel biomarker and therapeutic target in multiple myeloma. Expert Rev. Hematol. 2020, 13, 1201–1210. [Google Scholar] [CrossRef]
- Sahni, S.; Nahm, C.; Krisp, C.; Molloy, M.P.; Mehta, S.; Maloney, S.; Itchins, M.; Pavlakis, N.; Clarke, S.; Chan, D.; et al. Identification of Novel Biomarkers in Pancreatic Tumor Tissue to Predict Response to Neoadju-vant Chemotherapy. Front. Oncol. 2020, 10, 237. [Google Scholar] [CrossRef] [Green Version]
- Dauer, P.; Sharma, N.S.; Gupta, V.K.; Durden, B.; Hadad, R.; Banerjee, S.; Dudeja, V.; Saluja, A.; Banerjee, S. ER stress sensor, glu-cose regulatory protein 78 (GRP78) regulates redox status in pancreatic cancer thereby maintaining “stemness”. Cell Death Dis. 2019, 10, 132. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.J.; Jan, C.I.; Tsay, Y.G.; Yu, Y.H.; Huang, C.Y.; Lin, S.C.; Liu, C.J.; Chen, Y.S.; Lo, J.F.; Yu, C.C. Elimination of head and neck can-cer initiating cells through targeting glucose regulated protein78 signaling. Mol. Cancer 2010, 9, 283. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; He, M.; Zhao, L.; Wu, H.; Zhao, Y.; Han, L.; Wei, B.; Ye, D.; Lv, X.; Wang, Y.; et al. A novel HIF-2α targeted inhibitor suppresses hypoxia-induced breast cancer stemness via SOD2-mtROS-PDI/GPR78-UPRER axis. Cell Death Differ. 2022, 1–21. [Google Scholar] [CrossRef]
- Yang, Y.; Meng, W.-J.; Wang, Z.-Q. Cancer Stem Cells and the Tumor Microenvironment in Gastric Cancer. Front. Oncol. 2022, 11, 803974. [Google Scholar] [CrossRef]
- Wang, M.; Kaufman, R.J. The impact of the endoplasmic reticulum protein-folding environment on cancer development. Nat. Rev. Cancer 2014, 14, 581–597. [Google Scholar] [CrossRef]
- Lebeaupin, C.; Yong, J.; Kaufman, R.J. The Impact of the ER Unfolded Protein Response on Cancer Initiation and Progres-sion: Therapeutic Implications. Adv. Exp. Med. Biol. 2020, 1243, 113–131. [Google Scholar]
- Ishibashi, R.; Furusawa, Y.; Honda, H.; Watanabe, Y.; Fujisaka, S.; Nishikawa, M.; Ikushiro, S.; Kurihara, S.; Tabuchi, Y.; Tobe, K.; et al. Isoliquiritigenin Attenuates Adipose Tissue Inflammation and Metabolic Syndrome by Modifying Gut Bacteria Composition in Mice. Mol. Nutr. Food Res. 2022, 66, e2101119. [Google Scholar] [CrossRef]
- Yang, L.; Wang, D.; Zhang, Z.; Jiang, Y.; Liu, Y. Isoliquiritigenin alleviates diabetic symptoms via activating AMPK and inhib-iting mTORC1 signaling in diet-induced diabetic mice. Phytomedicine 2022, 98, 153950. [Google Scholar] [CrossRef]
- Rui-Zhi, T.; Ke-Huan, X.; Yuan, L.; Xiao, L.; Bing-Wen, Z.; Tong-Tong, L.; Li, W. Renoprotective effect of isoliquiritigenin on cis-platin-induced acute kidney injury through inhibition of FPR2 in macrophage. J. Pharmacol. Sci. 2022, 148, 56–64. [Google Scholar] [CrossRef]
- Wang, K.-L.; Yu, Y.-C.; Hsia, S.-M. Perspectives on the Role of Isoliquiritigenin in Cancer. Cancers 2021, 13, 115. [Google Scholar] [CrossRef]
- Raiter, A.; Lipovetsky, J.; Hyman, L.; Mugami, S.; Ben-Zur, T.; Yerushalmi, R. Chemotherapy Controls Metastasis Through Stimulatory Effects on GRP78 and Its Transcription Factor CREB3L1. Front. Oncol. 2020, 10, 1500. [Google Scholar] [CrossRef]
- Ishimoto, T.; Miyake, K.; Nandi, T.; Yashiro, M.; Onishi, N.; Huang, K.K.; Lin, S.J.; Kalpana, R.; Tay, S.T.; Suzuki, Y.; et al. Activation of Transforming Growth Factor Beta 1 Signaling in Gastric Cancer-associated Fibroblasts Increases Their Motility, via Expression of Rhomboid 5 Homolog 2, and Ability to Induce Invasiveness of Gastric Cancer Cells. Gastroenterology 2017, 153, 191–204.e16. [Google Scholar] [CrossRef]
- Loh, J.J.; Ma, S. The Role of Cancer-Associated Fibroblast as a Dynamic Player in Mediating Cancer Stemness in the Tumor Microenvironment. Front. Cell Dev. Biol. 2021, 9, 727640. [Google Scholar] [CrossRef]
- Wiechec, E.; Magan, M.; Matic, N.; Ansell-Schultz, A.; Kankainen, M.; Monni, O.; Johansson, A.-C.; Roberg, K. Cancer-Associated Fibroblasts Modulate Transcriptional Signatures Involved in Proliferation, Differentiation and Metastasis in Head and Neck Squamous Cell Carcinoma. Cancers 2021, 13, 3361. [Google Scholar] [CrossRef]
- Suh, J.; Kim, D.H.; Surh, Y.J. Resveratrol suppresses migration, invasion and stemness of human breast cancer cells by inter-fering with tumor-stromal cross-talk. Arch. Biochem. Biophys. 2018, 643, 62–71. [Google Scholar] [CrossRef]
- Yao, X.; Tu, Y.; Xu, Y.; Guo, Y.; Yao, F.; Zhang, X. Endoplasmic reticulum stress confers 5-fluorouracil resistance in breast can-cer cell via the GRP78/OCT4/lncRNA MIAT/AKT pathway. Am. J. Cancer Res. 2020, 10, 838–855. [Google Scholar]
- Gu, Y.J.; Li, H.D.; Zhao, L.; Zhao, S.; He, W.B.; Rui, L.; Su, C.; Zheng, H.C.; Su, R.J. GRP78 confers the resistance to 5-FU by activat-ing the c-Src/LSF/TS axis in hepatocellular carcinoma. Oncotarget 2015, 6, 33658–33674. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Wang, Z.; Peng, C.; You, J.; Shen, J.; Han, S.; Chen, J. Dietary compound isoliquiritigenin targets GRP78 to chemosen-sitize breast cancer stem cells via β-catenin/ABCG2 signaling. Carcinogenesis 2014, 35, 2544–2554. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.W.; Yu, C.C.; Hsieh, P.L.; Liao, Y.W.; Lu, M.Y.; Chu, P.M. Targeting oral cancer stemness and chemoresistance by isoliquiri-tigenin-mediated GRP78 regulation. Oncotarget 2017, 8, 93912–93923. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Khoonkari, M.; Avril, T.; Chevet, E.; Kruyt, F.A. The unfolded protein response as regulator of cancer stemness and differentiation: Mechanisms and implications for cancer therapy. Biochem. Pharmacol. 2021, 192, 114737. [Google Scholar] [CrossRef]
- Li, C.; Fan, Q.; Quan, H.; Nie, M.; Luo, Y.; Wang, L. The three branches of the unfolded protein response exhibit differential significance in breast cancer growth and stemness. Exp. Cell Res. 2018, 367, 170–185. [Google Scholar] [CrossRef]
- Chiu, C.C.; Lee, L.Y.; Li, Y.C.; Chen, Y.J.; Lu, Y.C.; Li, Y.L.; Wang, H.M.; Chang, J.T.; Cheng, A.J. Grp78 as a therapeutic target for re-fractory head-neck cancer with CD24−CD44+ stemness phenotype. Cancer Gene Ther. 2013, 20, 606–615. [Google Scholar] [CrossRef] [Green Version]
- Vasefifar, P.; Motafakkerazad, R.; Maleki, L.A.; Najafi, S.; Ghrobaninezhad, F.; Najafzadeh, B.; Alemohammad, H.; Amini, M.; Baghbanzadeh, A.; Baradaran, B. Nanog, as a key cancer stem cell marker in tumor progression. Gene 2022, 827, 146448. [Google Scholar] [CrossRef]
- Fatma, H.; Siddique, H.R.; Maurya, S.K. The multiple faces of NANOG in cancer: A therapeutic target to chemosensitize therapy-resistant cancers. Epigenomics 2021, 13, 1885–1900. [Google Scholar] [CrossRef]
- Pouremamali, F.; Vahedian, V.; Hassani, N.; Mirzaei, S.; Pouremamali, A.; Kazemzadeh, H.; Faridvand, Y.; Jafari-Gharabaghlou, D.; Nouri, M.; Maroufi, N.F. The role of SOX family in cancer stem cell maintenance: With a focus on SOX2. Pathol.-Res. Pract. 2022, 231, 153783. [Google Scholar] [CrossRef]
- Tian, T.; Zhang, Y.; Wang, S.; Zhou, J.; Xu, S. Sox2 enhances the tumorigenicity and chemoresistance of cancer stem-like cells derived from gastric cancer. J. Biomed. Res. 2012, 26, 336–345. [Google Scholar] [CrossRef]
- Poturnajova, M.; Kozovska, Z.; Matuskova, M. Aldehyde dehydrogenase 1A1 and 1A3 isoforms–mechanism of activation and regulation in cancer. Cell. Signal. 2021, 87, 110120. [Google Scholar] [CrossRef]
- Ye, Y.; Zhang, S.; Chen, Y.; Wang, X.; Wang, P. High ALDH1A1 expression indicates a poor prognosis in gastric neuroendo-crine carcinoma. Pathol. Res. Pract. 2018, 214, 268–272. [Google Scholar] [CrossRef]
- Li, K.; Guo, X.; Wang, Z.; Li, X.; Bu, Y.; Bai, X.; Zheng, L.; Huang, Y. The prognostic roles of ALDH1 isoenzymes in gastric cancer. Onco. Targets Ther. 2016, 9, 3405–3414. [Google Scholar] [CrossRef] [Green Version]
- Peñaranda-Fajardo, N.M.; Meijer, C.; Liang, Y.; Dijkstra, B.M.; Aguirre-Gamboa, R.; den Dunnen, W.F.A.; Kruyt, F.A.E. ER stress and UPR activation in glioblastoma: Identification of a noncanonical PERK mechanism regulating GBM stem cells through SOX2 modulation. Cell Death Dis. 2019, 10, 690. [Google Scholar] [CrossRef]
- Tseng, C.C.; Stanciauskas, R.; Zhang, P.; Woo, D.; Wu, K.; Kelly, K.; Gill, P.S.; Yu, M.; Pinaud, F.; Lee, A.S. GRP78 regulates CD44v membrane homeostasis and cell spreading in tamoxifen-resistant breast cance. Life Sci. Alliance 2019, 2, e201900377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, J.; Chen, Y.; Huang, S.; Cui, F.; Wang, X. Suppression of GRP78 sensitizes human colorectal cancer cells to oxaliplatin by downregulation of CD24. Oncol. Lett. 2018, 15, 9861–9867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, T.; Yashiro, M.; Nishii, T.; Matsuoka, J.; Fuyuhiro, Y.; Morisaki, T.; Fukuoka, T.; Shimizu, K.; Shimizu, T.; Miwa, A.; et al. Cancer-associated fibroblasts might sustain the stemness of scirrhous gastric cancer cells via transforming growth factor-β signaling. Int. J. Cancer 2013, 134, 1785–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, T.; Prasad, P.; Mukherjee, P.; Chaudhuri, S.R.; Chatterji, U.; Roy, S.S. Stemness and chemoresistance are imparted to the OC cells through TGFβ1 driven EMT. J. Cell Biochem. 2018, 119, 5775–5787. [Google Scholar] [CrossRef] [PubMed]
- Niu, N.; Yao, J.; Bast, R.C.; Sood, A.K.; Liu, J. IL-6 promotes drug resistance through formation of polyploid giant cancer cells and stromal fibroblast reprogramming. Oncogenesis 2021, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Narusaka, T.; Ohara, T.; Noma, K.; Nishiwaki, N.; Katsura, Y.; Kato, T.; Sato, H.; Tomono, Y.; Kikuchi, S.; Tazawa, H.; et al. Nanog is a promising chemoresistant stemness marker and therapeutic target by iron chelators for esophageal cancer. Int. J. Cancer 2021, 149, 347–357. [Google Scholar] [CrossRef]
- Ham, I.-H.; Oh, H.J.; Jin, H.; Bae, C.A.; Jeon, S.-M.; Choi, K.S.; Son, S.-Y.; Han, S.-U.; Brekken, R.A.; Lee, D.; et al. Targeting interleukin-6 as a strategy to overcome stroma-induced resistance to chemotherapy in gastric cancer. Mol. Cancer 2019, 18, 68. [Google Scholar] [CrossRef]
- Ham, I.H.; Lee, D.; Hur, H. Role of Cancer-Associated Fibroblast in Gastric Cancer Progression and Resistance to Treatments. J. Oncol. 2019, 2019, 6270784. [Google Scholar] [CrossRef]
- Kim, J.K.; Kang, K.A.; Piao, M.J.; Ryu, Y.S.; Han, X.; Fernando, P.M.D.J.; Oh, M.C.; Park, J.E.; Shilnikova, K.; Boo, S.J.; et al. Endoplasmic reticulum stress induces 5-fluorouracil resistance in human colon cancer cells. Environ. Toxicol. Pharmacol. 2016, 44, 128–133. [Google Scholar] [CrossRef]
- Zheng, H.-C.; Gong, B.-C.; Zhao, S. The meta and bioinformatics analysis of GRP78 expression in gastric cancer. Oncotarget 2017, 8, 73017–73028. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | No. Pf Patients |
|---|---|
| Normal tissue | 20 |
| Gastric carcinoma | 20 |
| Gender | |
| M | 10 |
| F | 10 |
| Age | |
| ≥60 | 13 |
| <60 | 7 |
| Histologic grade | |
| Moderately differentiated | 9 |
| Poorly differentiated | 11 |
| TNM stage | |
| I | 5 |
| II | 5 |
| III | 5 |
| IV | 5 |
| LN metastasis | |
| Absence | 6 |
| Presence | 14 |
| Survival (TNM stage) | |
| I | 5/5 |
| II | 2/5 |
| III | 1/5 |
| IV | 0/5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.-H.; Tsai, H.-Y.; Chen, C.-L.; Chen, J.-L.; Lu, C.-C.; Fang, Y.-P.; Wu, D.-C.; Huang, Y.-B.; Lin, M.-W. Isoliquiritigenin Inhibits Gastric Cancer Stemness, Modulates Tumor Microenvironment, and Suppresses Tumor Growth through Glucose-Regulated Protein 78 Downregulation. Biomedicines 2022, 10, 1350. https://doi.org/10.3390/biomedicines10061350
Lee C-H, Tsai H-Y, Chen C-L, Chen J-L, Lu C-C, Fang Y-P, Wu D-C, Huang Y-B, Lin M-W. Isoliquiritigenin Inhibits Gastric Cancer Stemness, Modulates Tumor Microenvironment, and Suppresses Tumor Growth through Glucose-Regulated Protein 78 Downregulation. Biomedicines. 2022; 10(6):1350. https://doi.org/10.3390/biomedicines10061350
Chicago/Turabian StyleLee, Chien-Hsing, Hsin-Yi Tsai, Chun-Lin Chen, Jen-Lung Chen, Chao-Chun Lu, Yi-Ping Fang, Deng-Chyang Wu, Yaw-Bin Huang, and Ming-Wei Lin. 2022. "Isoliquiritigenin Inhibits Gastric Cancer Stemness, Modulates Tumor Microenvironment, and Suppresses Tumor Growth through Glucose-Regulated Protein 78 Downregulation" Biomedicines 10, no. 6: 1350. https://doi.org/10.3390/biomedicines10061350