
Mechanistic Insights into Inorganic Nitrite-Mediated Vasodilation of Isolated Aortic Rings under Oxidative/Hypertensive Conditions and S-Nitros(yl)ation of Proteins in Germ-Free Mice

 , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Treatment
2.2. Isometric Tension Studies
2.3. Western Blotting Analysis of Xanthine Oxidase and Dehydrogenase
2.4. Protein S-Nitros(yl)ation Quantification by Dot Blot Technique
2.5. Statistical Analysis
3. Results
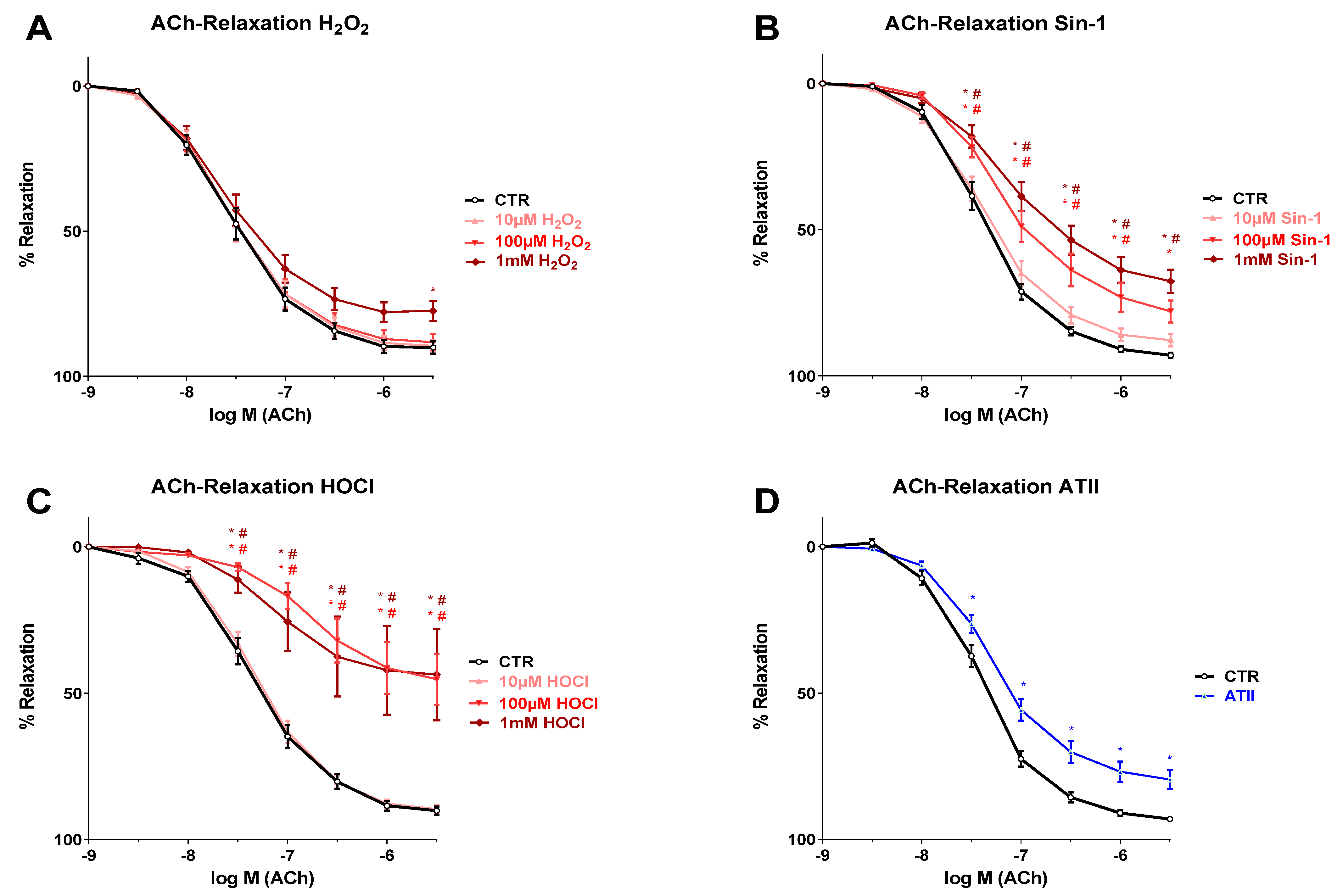
3.1. Effects of Different Oxidants on Endothelium-Dependent Vasodilation
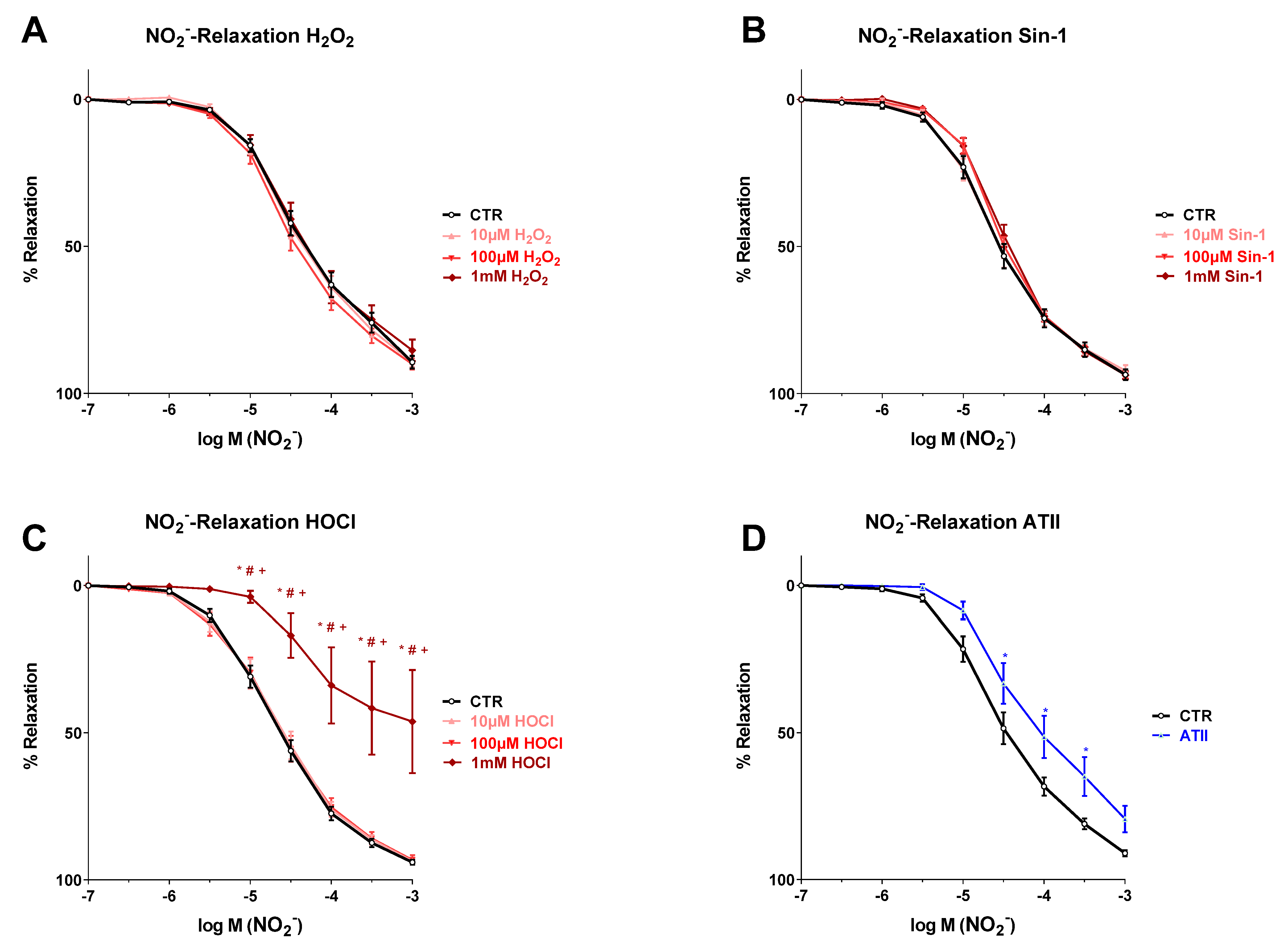
3.2. Effects of Different Oxidants on Nitrite-Induced Vasodilation
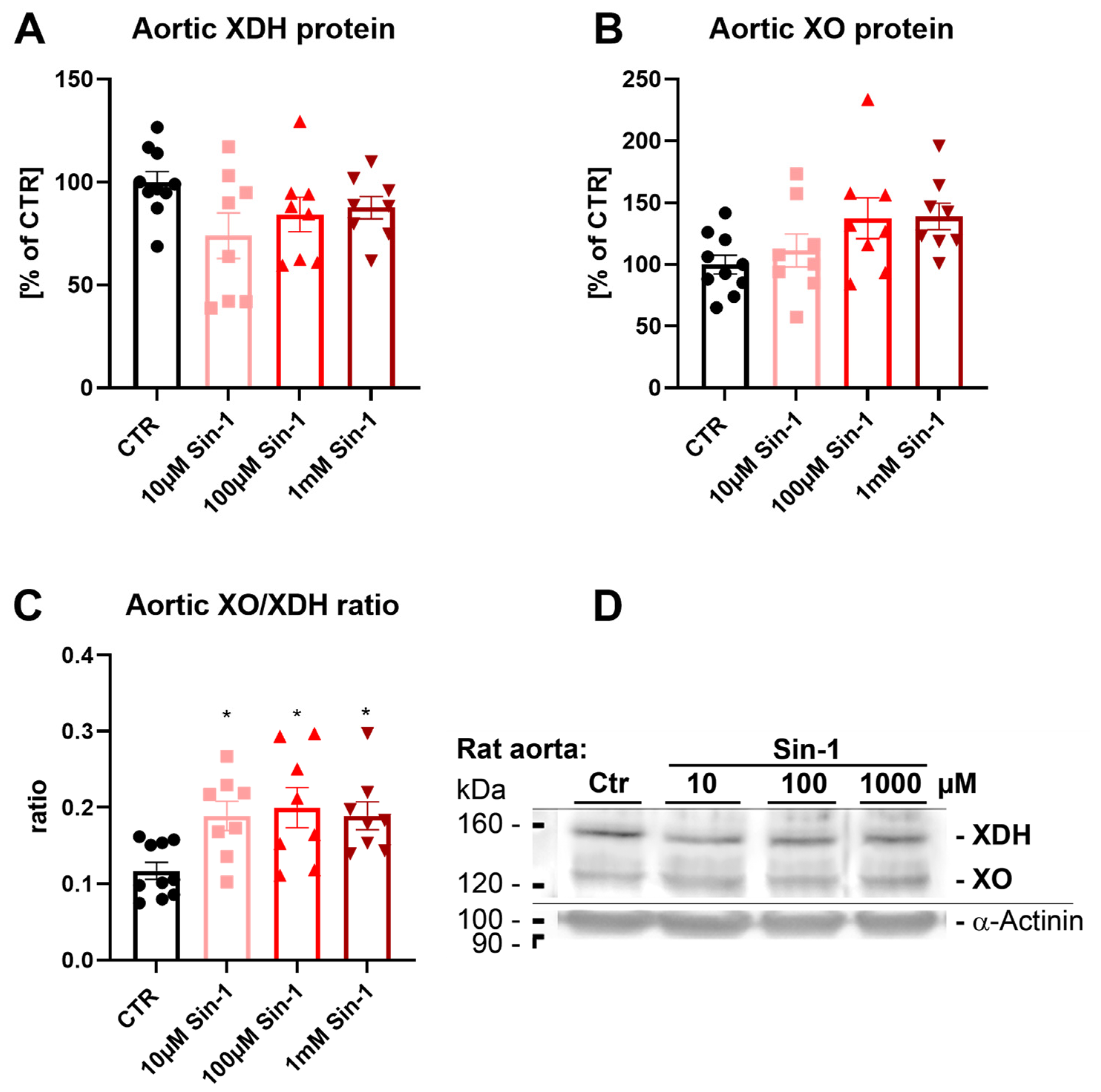
3.3. Effects of Sin-1 on Xanthine Oxidase to Dehydrogenase Conversion in Aortic Rings
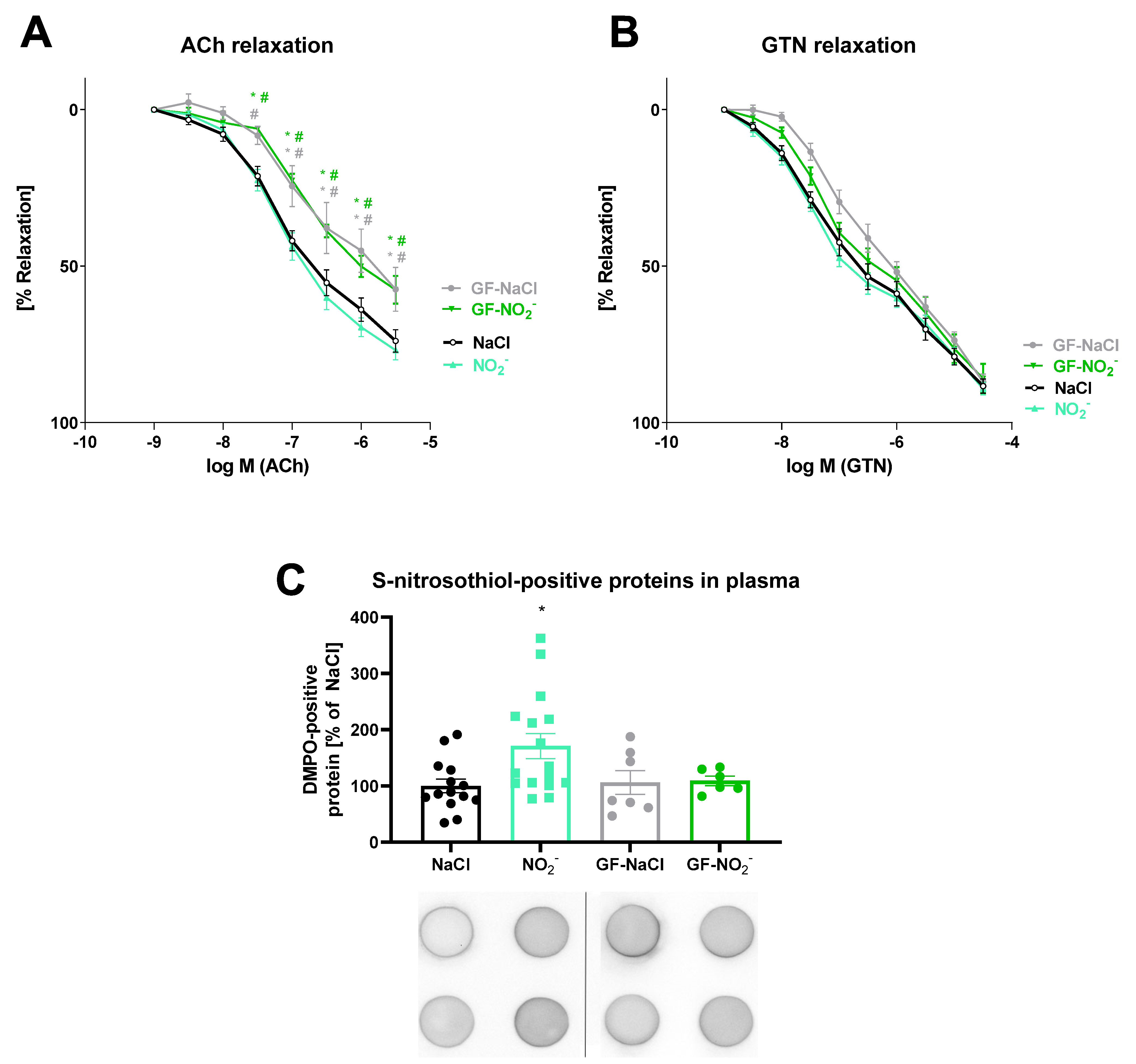
3.4. Effects of Inorganic Nitrite Treatment on Vasodilation and S-Nitros(yl)ation
4. Discussion
4.1. Mechanistic Considerations
4.2. Clinical Considerations
4.3. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perkovic, V.; Huxley, R.; Wu, Y.; Prabhakaran, D.; MacMahon, S. The burden of blood pressure-related disease: A neglected priority for global health. Hypertension 2007, 50, 991–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.; Mancia, G.; Spiering, W.; Agabiti Rosei, E.; Azizi, M.; Burnier, M.; Clement, D.L.; Coca, A.; de Simone, G.; Dominiczak, A.; et al. 2018 esc/esh guidelines for the management of arterial hypertension. Eur. Heart J. 2018, 39, 3021–3104. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.S.; Vos, T.; Flaxman, A.D.; Danaei, G.; Shibuya, K.; Adair-Rohani, H.; Amann, M.; Anderson, H.R.; Andrews, K.G.; Aryee, M.; et al. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990-2010: A systematic analysis for the global burden of disease study 2010. Lancet 2012, 380, 2224–2260. [Google Scholar] [CrossRef] [Green Version]
- Burnier, M.; Egan, B.M. Adherence in hypertension. Circ. Res. 2019, 124, 1124–1140. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Khan, H.; Heydon, E.; Shroufi, A.; Fahimi, S.; Moore, C.; Stricker, B.; Mendis, S.; Hofman, A.; Mant, J.; et al. Adherence to cardiovascular therapy: A meta-analysis of prevalence and clinical consequences. Eur. Heart J. 2013, 34, 2940–2948. [Google Scholar] [CrossRef] [Green Version]
- Luscher, T.F. Hypertension: An undertreated known risk factor revisited. Eur. Heart J. 2019, 40, 1991–1994. [Google Scholar] [CrossRef]
- Lobo, M.D.; Sobotka, P.A.; Pathak, A. Interventional procedures and future drug therapy for hypertension. Eur. Heart J. 2017, 38, 1101–1111. [Google Scholar] [CrossRef] [Green Version]
- McDonald, H.P.; Garg, A.X.; Haynes, R.B. Interventions to enhance patient adherence to medication prescriptions: Scientific review. JAMA 2002, 288, 2868–2879. [Google Scholar] [CrossRef]
- Song, P.; Wu, L.; Guan, W. Dietary nitrates, nitrites, and nitrosamines intake and the risk of gastric cancer: A meta-analysis. Nutrients 2015, 7, 9872–9895. [Google Scholar] [CrossRef]
- Habermeyer, M.; Roth, A.; Guth, S.; Diel, P.; Engel, K.H.; Epe, B.; Furst, P.; Heinz, V.; Humpf, H.U.; Joost, H.G.; et al. Nitrate and nitrite in the diet: How to assess their benefit and risk for human health. Mol. Nutr. Food Res. 2015, 59, 106–128. [Google Scholar] [CrossRef]
- Pattillo, C.B.; Bir, S.; Rajaram, V.; Kevil, C.G. Inorganic nitrite and chronic tissue ischaemia: A novel therapeutic modality for peripheral vascular diseases. Cardiovasc. Res. 2011, 89, 533–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, J.C.; Feelisch, M.; Horowitz, J.D.; Frenneaux, M.P.; Madhani, M. Pharmacology and therapeutic role of inorganic nitrite and nitrate in vasodilatation. Pharmacol. Ther. 2014, 144, 303–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacks, F.M.; Svetkey, L.P.; Vollmer, W.M.; Appel, L.J.; Bray, G.A.; Harsha, D.; Obarzanek, E.; Conlin, P.R.; Miller, E.R., 3rd; Simons-Morton, D.G.; et al. Effects on blood pressure of reduced dietary sodium and the dietary approaches to stop hypertension (dash) diet. Dash-sodium collaborative research group. N. Engl. J. Med. 2001, 344, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Woessner, M.N.; McIlvenna, L.C.; Ortiz de Zevallos, J.; Neil, C.J.; Allen, J.D. Dietary nitrate supplementation in cardiovascular health: An ergogenic aid or exercise therapeutic? Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H195–H212. [Google Scholar] [CrossRef]
- Bryan, N.S. Functional nitric oxide nutrition to combat cardiovascular disease. Curr. Atheroscler. Rep. 2018, 20, 21. [Google Scholar] [CrossRef]
- Jakubcik, E.M.; Rutherfurd-Markwick, K.; Chabert, M.; Wong, M.; Ali, A. Pharmacokinetics of nitrate and nitrite following beetroot juice drink consumption. Nutrients 2021, 13, 281. [Google Scholar] [CrossRef]
- Munzel, T.; Daiber, A. Inorganic nitrite and nitrate in cardiovascular therapy: A better alternative to organic nitrates as nitric oxide donors? Vascul. Pharmacol. 2018, 102, 1–10. [Google Scholar] [CrossRef]
- Stamm, P.; Oelze, M.; Steven, S.; Kroller-Schon, S.; Kvandova, M.; Kalinovic, S.; Jasztal, A.; Kij, A.; Kuntic, M.; Bayo Jimenez, M.T.; et al. Direct comparison of inorganic nitrite and nitrate on vascular dysfunction and oxidative damage in experimental arterial hypertension. Nitric Oxide 2021, 113–114, 57–69. [Google Scholar] [CrossRef]
- Amaral, J.H.; Ferreira, G.C.; Pinheiro, L.C.; Montenegro, M.F.; Tanus-Santos, J.E. Consistent antioxidant and antihypertensive effects of oral sodium nitrite in doca-salt hypertension. Redox Biol. 2015, 5, 340–346. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.M.; Kapil, V.; Fuentes-Calvo, I.; Bubb, K.J.; Pearl, V.; Milsom, A.B.; Khambata, R.; Maleki-Toyserkani, S.; Yousuf, M.; Benjamin, N.; et al. Enhanced vasodilator activity of nitrite in hypertension: Critical role for erythrocytic xanthine oxidoreductase and translational potential. Hypertension 2013, 61, 1091–1102. [Google Scholar] [CrossRef] [Green Version]
- Ling, W.C.; Lau, Y.S.; Murugan, D.D.; Vanhoutte, P.M.; Mustafa, M.R. Sodium nitrite causes relaxation of the isolated rat aorta: By stimulating both endothelial no synthase and activating soluble guanylyl cyclase in vascular smooth muscle. Vascul. Pharmacol. 2015, 74, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Jansson, E.A.; Huang, L.; Malkey, R.; Govoni, M.; Nihlen, C.; Olsson, A.; Stensdotter, M.; Petersson, J.; Holm, L.; Weitzberg, E.; et al. A mammalian functional nitrate reductase that regulates nitrite and nitric oxide homeostasis. Nat. Chem. Biol. 2008, 4, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Petersson, J.; Jadert, C.; Phillipson, M.; Borniquel, S.; Lundberg, J.O.; Holm, L. Physiological recycling of endogenous nitrate by oral bacteria regulates gastric mucus thickness. Free Radic. Biol. Med. 2015, 89, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Moretti, C.; Zhuge, Z.; Zhang, G.; Haworth, S.M.; Paulo, L.L.; Guimaraes, D.D.; Cruz, J.C.; Montenegro, M.F.; Cordero-Herrera, I.; Braga, V.A.; et al. The obligatory role of host microbiota in bioactivation of dietary nitrate. Free Radic. Biol. Med. 2019, 145, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, F.; Backhed, F. The gut microbiota--masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Esser, D.; Lange, J.; Marinos, G.; Sieber, M.; Best, L.; Prasse, D.; Bathia, J.; Ruhlemann, M.C.; Boersch, K.; Jaspers, C.; et al. Functions of the microbiota for the physiology of animal metaorganisms. J. Innate Immun. 2019, 11, 393–404. [Google Scholar] [CrossRef]
- Xiao, L.; Feng, Q.; Liang, S.; Sonne, S.B.; Xia, Z.; Qiu, X.; Li, X.; Long, H.; Zhang, J.; Zhang, D.; et al. A catalog of the mouse gut metagenome. Nat. Biotechnol. 2015, 33, 1103–1108. [Google Scholar] [CrossRef]
- Marques, F.Z.; Nelson, E.; Chu, P.Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-fiber diet and acetate supplementation change the gut microbiota and prevent the development of hypertension and heart failure in hypertensive mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef]
- Jie, Z.; Xia, H.; Zhong, S.L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat. Commun. 2017, 8, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karbach, S.H.; Schonfelder, T.; Brandao, I.; Wilms, E.; Hormann, N.; Jackel, S.; Schuler, R.; Finger, S.; Knorr, M.; Lagrange, J.; et al. Gut microbiota promote angiotensin ii-induced arterial hypertension and vascular dysfunction. J. Am. Heart Assoc. 2016, 5, e003698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formes, H.; Bernardes, J.P.; Mann, A.; Bayer, F.; Pontarollo, G.; Kiouptsi, K.; Schafer, K.; Attig, S.; Nikolova, T.; Hofmann, T.G.; et al. The gut microbiota instructs the hepatic endothelial cell transcriptome. iScience 2021, 24, 103092. [Google Scholar] [CrossRef]
- Vikram, A.; Kim, Y.R.; Kumar, S.; Li, Q.; Kassan, M.; Jacobs, J.S.; Irani, K. Vascular microrna-204 is remotely governed by the microbiome and impairs endothelium-dependent vasorelaxation by downregulating sirtuin1. Nat. Commun. 2016, 7, 12565. [Google Scholar] [CrossRef] [PubMed]
- Zollbrecht, C.; Persson, A.E.; Lundberg, J.O.; Weitzberg, E.; Carlstrom, M. Nitrite-mediated reduction of macrophage nadph oxidase activity is dependent on xanthine oxidoreductase-derived nitric oxide but independent of s-nitrosation. Redox Biol. 2016, 10, 119–127. [Google Scholar] [CrossRef]
- Schuhmacher, S.; Wenzel, P.; Schulz, E.; Oelze, M.; Mang, C.; Kamuf, J.; Gori, T.; Jansen, T.; Knorr, M.; Karbach, S.; et al. Pentaerythritol tetranitrate improves angiotensin ii-induced vascular dysfunction via induction of heme oxygenase-1. Hypertension 2010, 55, 897–904. [Google Scholar] [CrossRef] [Green Version]
- Backhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. USA 2007, 104, 979–984. [Google Scholar] [CrossRef] [Green Version]
- Coggan, A.R.; Racette, S.B.; Thies, D.; Peterson, L.R.; Stratford, R.E., Jr. Simultaneous pharmacokinetic analysis of nitrate and its reduced metabolite, nitrite, following ingestion of inorganic nitrate in a mixed patient population. Pharm. Res. 2020, 37, 235. [Google Scholar] [CrossRef]
- Kevil, C.G.; Kolluru, G.K.; Pattillo, C.B.; Giordano, T. Inorganic nitrite therapy: Historical perspective and future directions. Free Radic. Biol. Med. 2011, 51, 576–593. [Google Scholar] [CrossRef] [Green Version]
- Kuntic, M.; Oelze, M.; Steven, S.; Kroller-Schon, S.; Stamm, P.; Kalinovic, S.; Frenis, K.; Vujacic-Mirski, K.; Bayo Jimenez, M.T.; Kvandova, M.; et al. Short-term e-cigarette vapour exposure causes vascular oxidative stress and dysfunction: Evidence for a close connection to brain damage and a key role of the phagocytic nadph oxidase (nox-2). Eur. Heart J. 2019, 41, 2472–2483. [Google Scholar] [CrossRef] [Green Version]
- Laursen, J.B.; Rajagopalan, S.; Galis, Z.; Tarpey, M.; Freeman, B.A.; Harrison, D.G. Role of superoxide in angiotensin ii-induced but not catecholamine-induced hypertension. Circulation 1997, 95, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, R.; Billiar, T.R.; Stoyanovsky, D.A. Studies toward the analysis of s-nitrosoproteins. Org. Biomol. Chem. 2009, 7, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Crow, J.P.; Beckman, J.S. Reaction between nitric oxide, superoxide, and peroxynitrite: Footprints of peroxynitrite in vivo. Adv. Pharmacol. 1995, 35, 17–43. [Google Scholar]
- Beckman, J.S.; Koppenol, W.H. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol. 1996, 271, C1424–C1437. [Google Scholar] [CrossRef] [Green Version]
- Koppenol, W.H.; Moreno, J.J.; Pryor, W.A.; Ischiropoulos, H.; Beckman, J.S. Peroxynitrite, a cloaked oxidant formed by nitric oxide and superoxide. Chem. Res. Toxicol. 1992, 5, 834–842. [Google Scholar] [CrossRef]
- Gryglewski, R.J.; Palmer, R.M.; Moncada, S. Superoxide anion is involved in the breakdown of endothelium-derived vascular relaxing factor. Nature 1986, 320, 454–456. [Google Scholar] [CrossRef]
- Zou, M.H.; Cohen, R.; Ullrich, V. Peroxynitrite and vascular endothelial dysfunction in diabetes mellitus. Endothelium 2004, 11, 89–97. [Google Scholar] [CrossRef]
- Munzel, T.; Daiber, A.; Ullrich, V.; Mulsch, A. Vascular consequences of endothelial nitric oxide synthase uncoupling for the activity and expression of the soluble guanylyl cyclase and the cgmp-dependent protein kinase. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1551–1557. [Google Scholar] [CrossRef]
- Tropea, T.; Wareing, M.; Greenwood, S.L.; Feelisch, M.; Sibley, C.P.; Cottrell, E.C. Nitrite mediated vasorelaxation in human chorionic plate vessels is enhanced by hypoxia and dependent on the no-sgc-cgmp pathway. Nitric Oxide 2018, 80, 82–88. [Google Scholar] [CrossRef]
- Amdahl, M.B.; DeMartino, A.W.; Gladwin, M.T. Inorganic nitrite bioactivation and role in physiological signaling and therapeutics. Biol. Chem. 2019, 401, 201–211. [Google Scholar] [CrossRef]
- Godber, B.L.; Doel, J.J.; Sapkota, G.P.; Blake, D.R.; Stevens, C.R.; Eisenthal, R.; Harrison, R. Reduction of nitrite to nitric oxide catalyzed by xanthine oxidoreductase. J. Biol. Chem. 2000, 275, 7757–7763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ormerod, J.O.; Ashrafian, H.; Maher, A.R.; Arif, S.; Steeples, V.; Born, G.V.; Egginton, S.; Feelisch, M.; Watkins, H.; Frenneaux, M.P. The role of vascular myoglobin in nitrite-mediated blood vessel relaxation. Cardiovasc. Res. 2011, 89, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.G.; Balendran, A.; Harrison, R.; Wolstenholme, A.; Bulkley, G.B. Xanthine oxidoreductase: Dehydrogenase to oxidase conversion. Biochem. Soc. Trans. 1997, 25, 530S. [Google Scholar] [CrossRef]
- Schulz, E.; Wenzel, P.; Munzel, T.; Daiber, A. Mitochondrial redox signaling: Interaction of mitochondrial reactive oxygen species with other sources of oxidative stress. Antioxid. Redox Signal. 2014, 20, 308–324. [Google Scholar] [CrossRef] [PubMed]
- Maia, L.B.; Moura, J.J.G. Putting xanthine oxidoreductase and aldehyde oxidase on the no metabolism map: Nitrite reduction by molybdoenzymes. Redox Biol. 2018, 19, 274–289. [Google Scholar] [CrossRef] [PubMed]
- Carlstrom, M.; Larsen, F.J.; Nystrom, T.; Hezel, M.; Borniquel, S.; Weitzberg, E.; Lundberg, J.O. Dietary inorganic nitrate reverses features of metabolic syndrome in endothelial nitric oxide synthase-deficient mice. Proc. Natl. Acad. Sci. USA 2010, 107, 17716–17720. [Google Scholar] [CrossRef] [Green Version]
- Cordero-Herrera, I.; Kozyra, M.; Zhuge, Z.; McCann Haworth, S.; Moretti, C.; Peleli, M.; Caldeira-Dias, M.; Jahandideh, A.; Huirong, H.; Cruz, J.C.; et al. Amp-activated protein kinase activation and nadph oxidase inhibition by inorganic nitrate and nitrite prevent liver steatosis. Proc. Natl. Acad. Sci. USA 2019, 116, 217–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, A.; Bond, R.; McLean, P.; Uppal, R.; Benjamin, N.; Ahluwalia, A. Reduction of nitrite to nitric oxide during ischemia protects against myocardial ischemia-reperfusion damage. Proc. Natl. Acad. Sci. USA 2004, 101, 13683–13688. [Google Scholar] [CrossRef] [Green Version]
- Duranski, M.R.; Greer, J.J.; Dejam, A.; Jaganmohan, S.; Hogg, N.; Langston, W.; Patel, R.P.; Yet, S.F.; Wang, X.; Kevil, C.G.; et al. Cytoprotective effects of nitrite during in vivo ischemia-reperfusion of the heart and liver. J. Clin. Investig. 2005, 115, 1232–1240. [Google Scholar] [CrossRef] [Green Version]
- Raat, N.J.; Noguchi, A.C.; Liu, V.B.; Raghavachari, N.; Liu, D.; Xu, X.; Shiva, S.; Munson, P.J.; Gladwin, M.T. Dietary nitrate and nitrite modulate blood and organ nitrite and the cellular ischemic stress response. Free Radic. Biol. Med. 2009, 47, 510–517. [Google Scholar] [CrossRef] [Green Version]
- Nossaman, V.E.; Nossaman, B.D.; Kadowitz, P.J. Nitrates and nitrites in the treatment of ischemic cardiac disease. Cardiol. Rev. 2010, 18, 190–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaskonen, T.; Mervaala, E.; Teravainen, T.L.; Laakso, J.; Karppanen, H.; Vapaatalo, H. Cardiovascular effects of dietary salts and isosorbide-5-mononitrate in spontaneously hypertensive rats. Blood Press. 1998, 7, 184–192. [Google Scholar] [PubMed]
- Weber, M.; Lauer, N.; Mulsch, A.; Kojda, G. The effect of peroxynitrite on the catalytic activity of soluble guanylyl cyclase. Free Radic. Biol. Med. 2001, 31, 1360–1367. [Google Scholar] [CrossRef]
- Brune, B.; Schmidt, K.U.; Ullrich, V. Activation of soluble guanylate cyclase by carbon monoxide and inhibition by superoxide anion. Eur. J. Biochem. 1990, 192, 683–688. [Google Scholar] [CrossRef]
- Montenegro, M.F.; Amaral, J.H.; Pinheiro, L.C.; Sakamoto, E.K.; Ferreira, G.C.; Reis, R.I.; Marcal, D.M.; Pereira, R.P.; Tanus-Santos, J.E. Sodium nitrite downregulates vascular nadph oxidase and exerts antihypertensive effects in hypertension. Free Radic. Biol. Med. 2011, 51, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Rizzi, E.; Amaral, J.H.; Guimaraes, D.A.; Conde-Tella, S.O.; Pinheiro, L.C.; Gerlach, R.F.; Castro, M.M.; Tanus-Santos, J.E. Nitrite treatment downregulates vascular mmp-2 activity and inhibits vascular remodeling in hypertension independently of its antihypertensive effects. Free Radic. Biol. Med. 2019, 130, 234–243. [Google Scholar] [CrossRef]
- Carlstrom, M.; Persson, A.E.; Larsson, E.; Hezel, M.; Scheffer, P.G.; Teerlink, T.; Weitzberg, E.; Lundberg, J.O. Dietary nitrate attenuates oxidative stress, prevents cardiac and renal injuries, and reduces blood pressure in salt-induced hypertension. Cardiovasc. Res. 2011, 89, 574–585. [Google Scholar] [CrossRef]
- Hoggatt, A.F.; Hoggatt, J.; Honerlaw, M.; Pelus, L.M. A spoonful of sugar helps the medicine go down: A novel technique to improve oral gavage in mice. J. Am. Assoc. Lab. Anim. Sci. 2010, 49, 329–334. [Google Scholar]
- Oliveira-Paula, G.H.; Pinheiro, L.C.; Tanus-Santos, J.E. Mechanisms impairing blood pressure responses to nitrite and nitrate. Nitric Oxide 2019, 85, 35–43. [Google Scholar] [CrossRef]
- Wang, D.D.; Nguyen, L.H.; Li, Y.; Yan, Y.; Ma, W.; Rinott, E.; Ivey, K.L.; Shai, I.; Willett, W.C.; Hu, F.B.; et al. The gut microbiome modulates the protective association between a mediterranean diet and cardiometabolic disease risk. Nat. Med. 2021, 27, 333–343. [Google Scholar] [CrossRef]
- Avery, E.G.; Bartolomaeus, H.; Maifeld, A.; Marko, L.; Wiig, H.; Wilck, N.; Rosshart, S.P.; Forslund, S.K.; Muller, D.N. The gut microbiome in hypertension: Recent advances and future perspectives. Circ. Res. 2021, 128, 934–950. [Google Scholar] [CrossRef] [PubMed]
- Zhurakivska, K.; Troiano, G.; Caponio, V.C.A.; Dioguardi, M.; Laino, L.; Maffione, A.B.; Lo Muzio, L. Do changes in oral microbiota correlate with plasma nitrite response? A systematic review. Front. Physiol. 2019, 10, 1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhatalo, A.; L’Heureux, J.E.; Kelly, J.; Blackwell, J.R.; Wylie, L.J.; Fulford, J.; Winyard, P.G.; Williams, D.W.; van der Giezen, M.; Jones, A.M. Network analysis of nitrate-sensitive oral microbiome reveals interactions with cognitive function and cardiovascular health across dietary interventions. Redox Biol. 2021, 41, 101933. [Google Scholar] [CrossRef] [PubMed]
- Pignatelli, P.; Fabietti, G.; Ricci, A.; Piattelli, A.; Curia, M.C. How periodontal disease and presence of nitric oxide reducing oral bacteria can affect blood pressure. Int. J. Mol. Sci. 2020, 21, 7538. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D. Dietary and policy priorities for cardiovascular disease, diabetes, and obesity: A comprehensive review. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef]
- Dixon, A.; Robertson, K.; Yung, A.; Que, M.; Randall, H.; Wellalagodage, D.; Cox, T.; Robertson, D.; Chi, C.; Sun, J. Efficacy of probiotics in patients of cardiovascular disease risk: A systematic review and meta-analysis. Curr. Hypertens. Rep. 2020, 22, 74. [Google Scholar] [CrossRef]
- Kapil, V.; Milsom, A.B.; Okorie, M.; Maleki-Toyserkani, S.; Akram, F.; Rehman, F.; Arghandawi, S.; Pearl, V.; Benjamin, N.; Loukogeorgakis, S.; et al. Inorganic nitrate supplementation lowers blood pressure in humans: Role for nitrite-derived no. Hypertension 2010, 56, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Kevil, C.G.; Lefer, D.J. Review focus on inorganic nitrite and nitrate in cardiovascular health and disease. Cardiovasc. Res. 2011, 89, 489–491. [Google Scholar] [CrossRef] [Green Version]
- Bahadoran, Z.; Mirmiran, P.; Ghasemi, A.; Carlstrom, M.; Azizi, F.; Hadaegh, F. Association between dietary intakes of nitrate and nitrite and the risk of hypertension and chronic kidney disease: Tehran lipid and glucose study. Nutrients 2016, 8, 811. [Google Scholar] [CrossRef]
- Henrohn, D.; Bjorkstrand, K.; Lundberg, J.O.; Granstam, S.O.; Baron, T.; Ingimarsdottir, I.J.; Hedenstrom, H.; Malinovschi, A.; Wernroth, M.L.; Jansson, M.; et al. Effects of oral supplementation with nitrate-rich beetroot juice in patients with pulmonary arterial hypertension-results from beet-pah, an exploratory randomized, double-blind, placebo-controlled, crossover study. J. Card. Fail. 2018, 24, 640–653. [Google Scholar] [CrossRef]
- Aboulgheit, A.; Karbasiafshar, C.; Zhang, Z.; Sabra, M.; Shi, G.; Tucker, A.; Sodha, N.; Abid, M.R.; Sellke, F.W. Lactobacillus plantarum probiotic induces nrf2-mediated antioxidant signaling and enos expression resulting in improvement of myocardial diastolic function. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H839–H849. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.K.; Idris-Khodja, N.; Auger, C.; Alhosin, M.; Boehm, N.; Oswald-Mammosser, M.; Schini-Kerth, V.B. Probiotics (vsl#3) prevent endothelial dysfunction in rats with portal hypertension: Role of the angiotensin system. PLoS ONE 2014, 9, e97458. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stamm, P.; Kalinovic, S.; Oelze, M.; Steven, S.; Czarnowski, A.; Kvandova, M.; Bayer, F.; Reinhardt, C.; Münzel, T.; Daiber, A. Mechanistic Insights into Inorganic Nitrite-Mediated Vasodilation of Isolated Aortic Rings under Oxidative/Hypertensive Conditions and S-Nitros(yl)ation of Proteins in Germ-Free Mice. Biomedicines 2022, 10, 730. https://doi.org/10.3390/biomedicines10030730
Stamm P, Kalinovic S, Oelze M, Steven S, Czarnowski A, Kvandova M, Bayer F, Reinhardt C, Münzel T, Daiber A. Mechanistic Insights into Inorganic Nitrite-Mediated Vasodilation of Isolated Aortic Rings under Oxidative/Hypertensive Conditions and S-Nitros(yl)ation of Proteins in Germ-Free Mice. Biomedicines. 2022; 10(3):730. https://doi.org/10.3390/biomedicines10030730
Chicago/Turabian StyleStamm, Paul, Sanela Kalinovic, Matthias Oelze, Sebastian Steven, Alexander Czarnowski, Miroslava Kvandova, Franziska Bayer, Christoph Reinhardt, Thomas Münzel, and Andreas Daiber. 2022. "Mechanistic Insights into Inorganic Nitrite-Mediated Vasodilation of Isolated Aortic Rings under Oxidative/Hypertensive Conditions and S-Nitros(yl)ation of Proteins in Germ-Free Mice" Biomedicines 10, no. 3: 730. https://doi.org/10.3390/biomedicines10030730