
An Early Th1 Response Is a Key Factor for a Favorable COVID-19 Evolution

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Patients
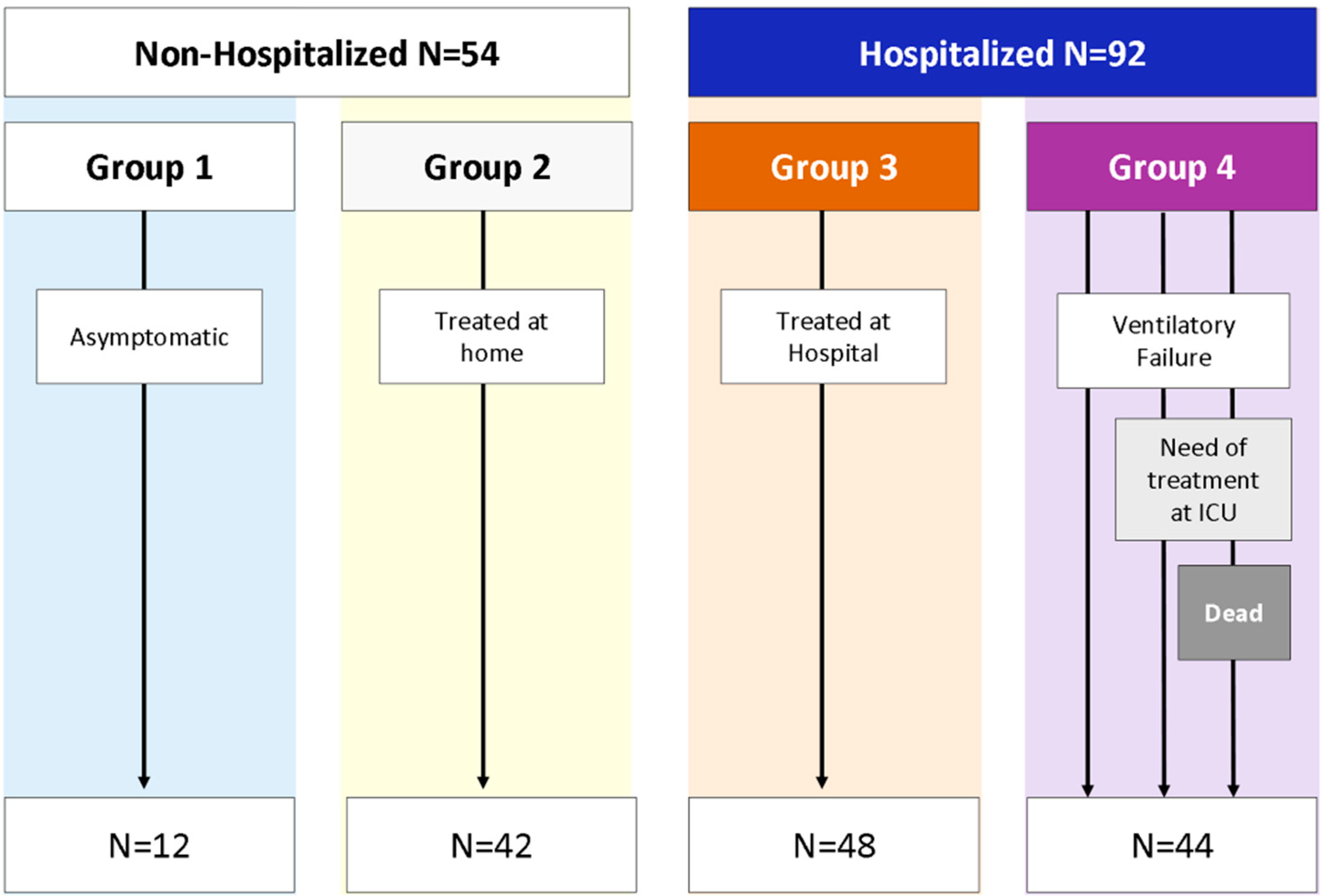
2.3. Patient Classification
2.4. Study Definitions
2.5. Data Collection
2.6. Samples
2.7. T-Cell Subsets
2.8. The Subsets
2.9. Evaluation of Anti-SARS-CoV-2 Antibodies
2.10. Statistical Analysis
3. Results
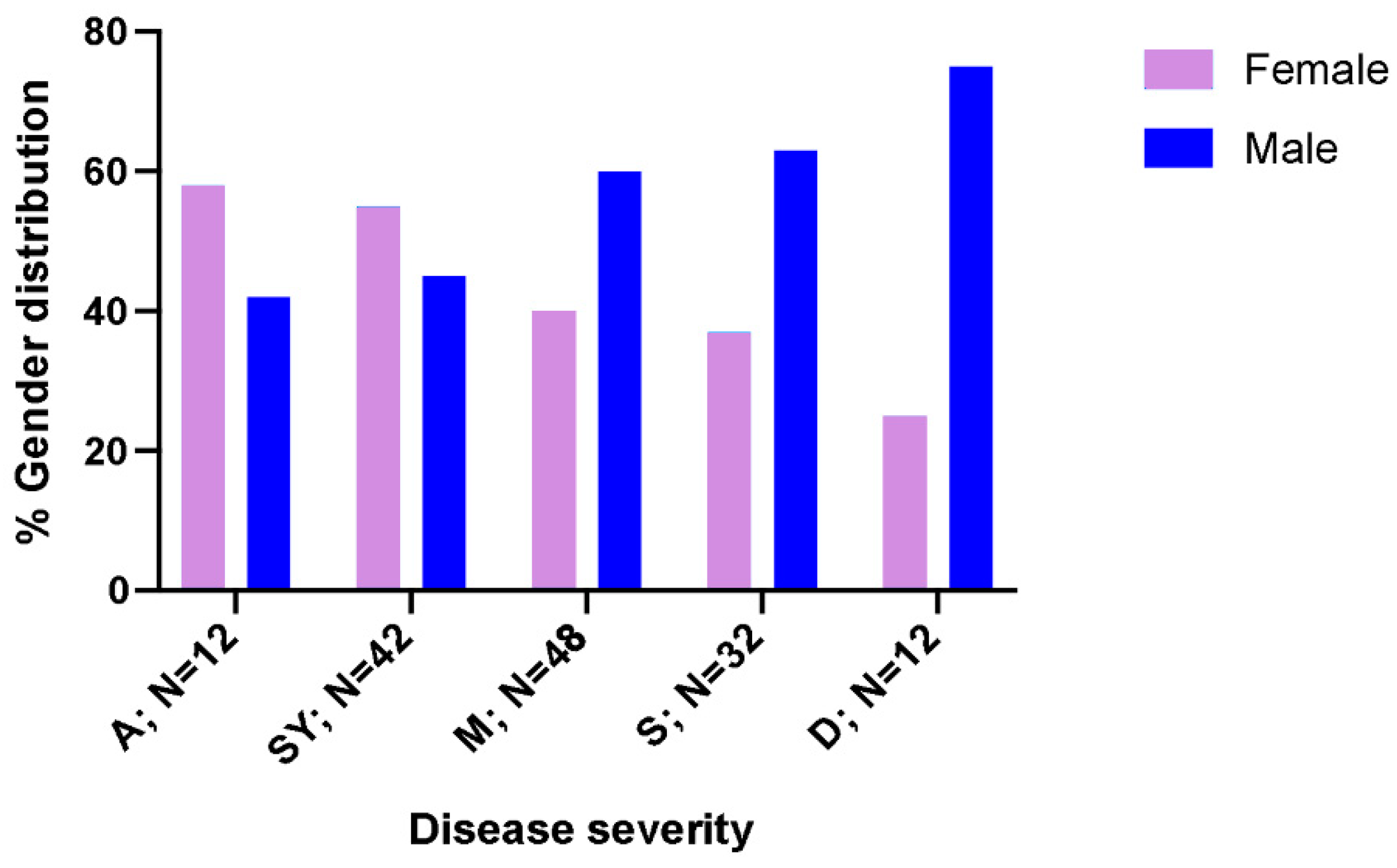
3.1. Population Characteristics and Biochemical Markers
3.2. Specificity of Anti-SARS-CoV-2 Antibodies
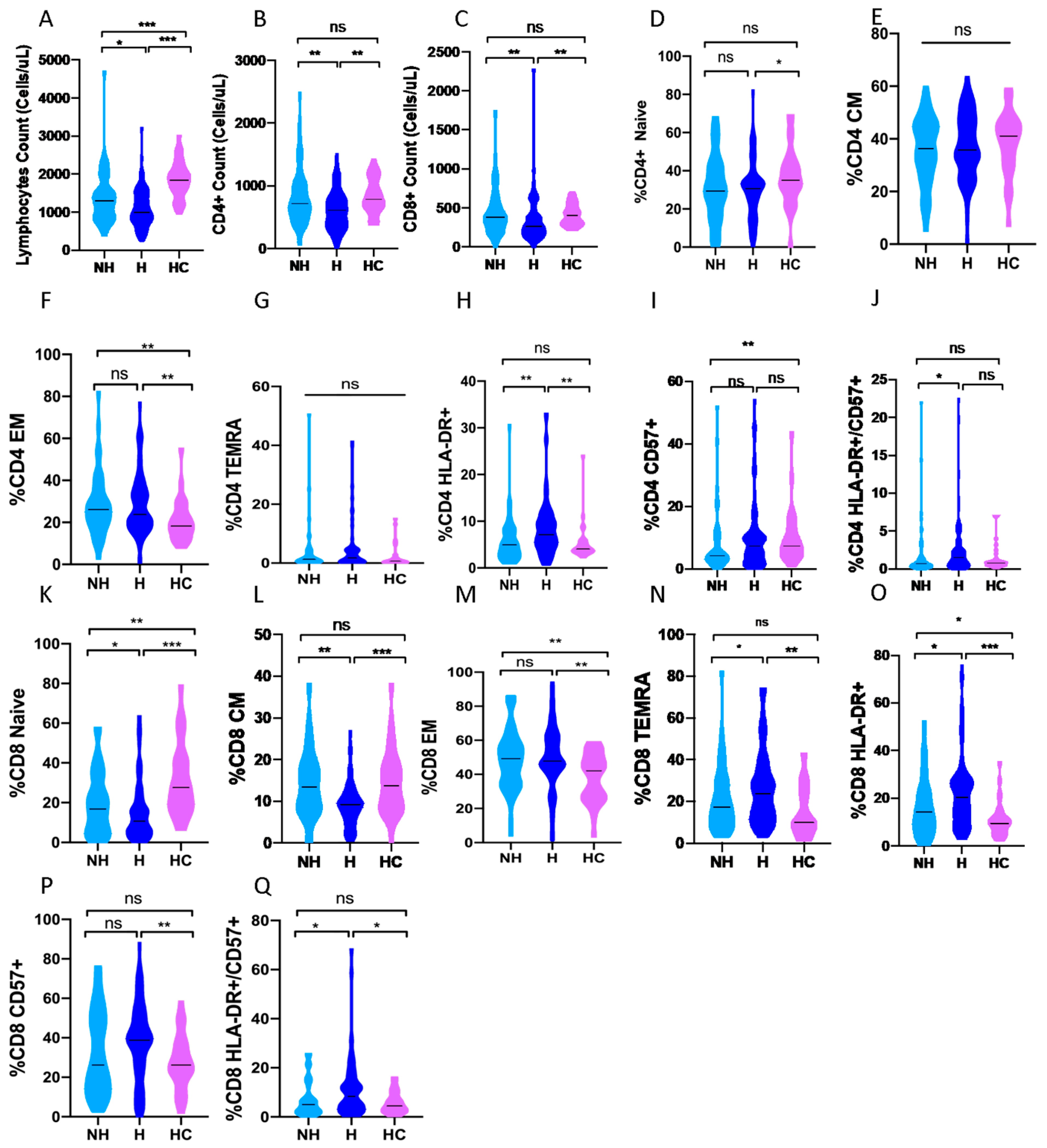
3.3. CD4 and CD8 Subpopulations and the Severity of the Disease
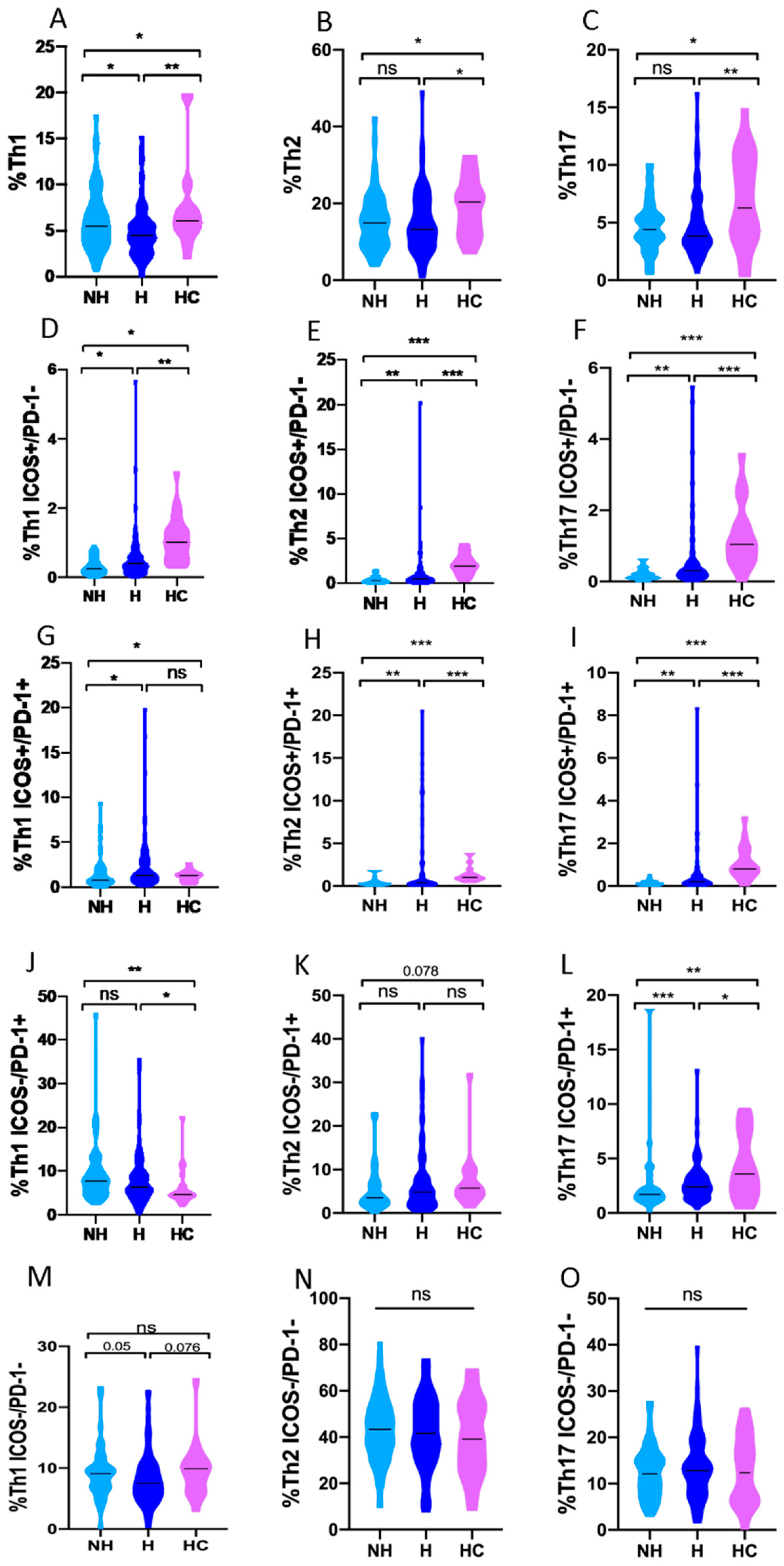
3.4. Th1, Th2 and Th17 Subsets in COVID-19 Patients and Healthy Controls
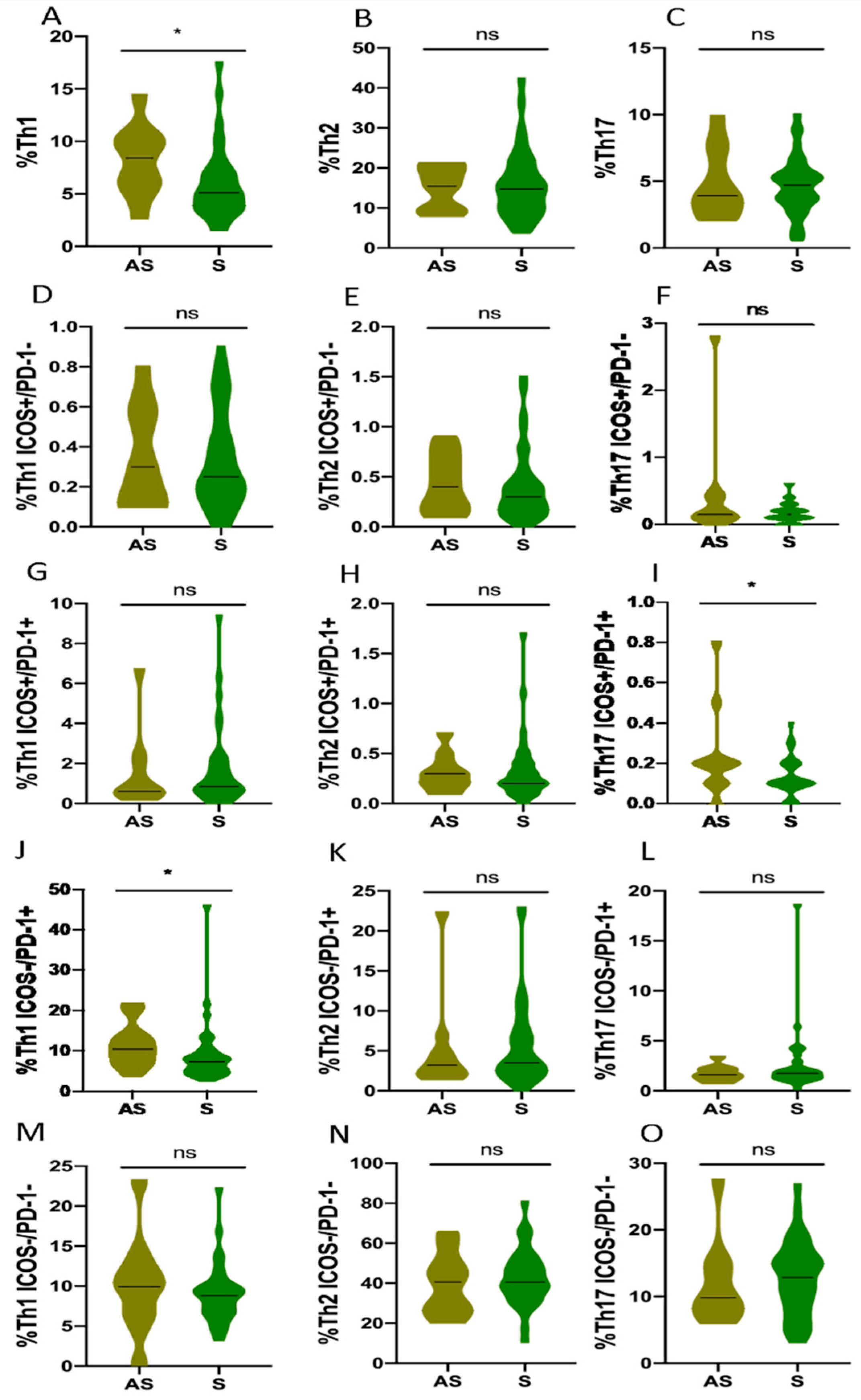
3.5. Th1, Th2 and Th17 Subsets in Non-Hospitalized and Hospitalized COVID-19 Patients
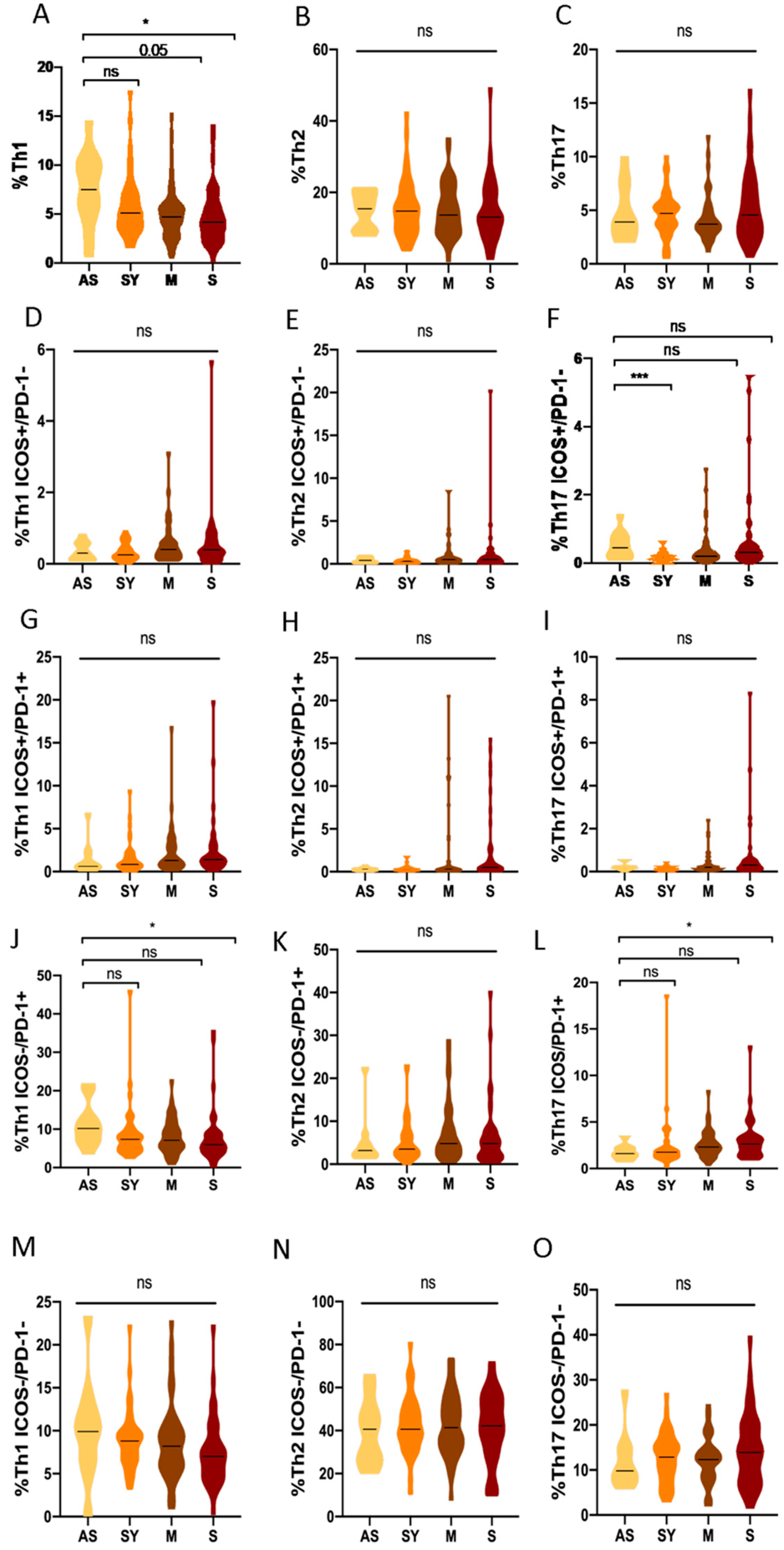
3.6. Th1, Th2 and Th17 Subsets in Asymptomatic COVID-19 Patients Compared to Different Clinical Profiles
3.7. Analysis of Comorbidities in COVID-19 Patients
3.8. Multivariate Analysis in COVID-19 Disease
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bohn, M.K.; Hall, A.; Sepiashvili, L.; Jung, B.; Steele, S.; Adeli, K. Pathophysiology of COVID-19: Mechanisms Underlying Disease Severity and Progression. Physiology 2020, 35, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.-H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Auto-antibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.; Espinosa, G.; Lalueza, A.; Bravo-Gallego, L.Y.; Diaz-Simon, R.; Garcinuno, S.; Gil-Etayo, J.; Moises, J.; Naranjo, L.; Prieto-Gonzalez, S.; et al. Beta-2-glycoprotein-i deficiency could precipitate an antiphospholipid syndrome-like prothrombotic situation in patients with coronavirus disease 2019. ACR Open Rheumatol. 2021, 3, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Hadid, T.; Kafri, Z.; Al-Katib, A. Coagulation and anticoagulation in COVID-19. Blood Rev. 2021, 47, 100761. [Google Scholar] [CrossRef] [PubMed]
- Ellinghaus, D.; Degenhardt, F.; Bujanda, L.; Buti, M.; Albillos, A.; Invernizzi, P.; Fernandez, J.; Prati, D.; Baselli, G.; Asselta, R.; et al. Genomewide association study of severe COVID-19 with respiratory failure. N. Engl. J. Med. 2020, 383, 1522–1534. [Google Scholar]
- Zhang, Q.; Bastard, P.; Liu, Z.; Le Pen, J.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.D.; Hodeib, S.; Korol, C.; et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 2020, 370, eabd4570. [Google Scholar] [CrossRef]
- Cabrera-Marante, O.; Rodriguez de Frias, E.; Pleguezuelo, D.E.; Allende, L.M.; Serrano, A.; Laguna-Goya, R.; Mancebo, M.E.; Talayero, P.; Alvarez-Vallina, L.; Morales, P.; et al. Perforin gene variant a91v in young patients with severe COVID-19. Haematologica 2020, 105, 2844–2846. [Google Scholar] [CrossRef]
- Clerici, M.; Shearer, G.M. The Th1-Th2 hypothesis of HIV infection: New insights. Immunol. Today 1994, 15, 575–581. [Google Scholar] [CrossRef]
- Mosmann, T.R.; Sad, S. The expanding universe of T-cell subsets: Th1, Th2 and more. Immunol. Today 1996, 17, 138–146. [Google Scholar] [CrossRef]
- Saini, C.; Tarique, M.; Rai, R.; Siddiqui, A.; Khanna, N.; Sharma, A. T helper cells in leprosy: An update. Immunol. Lett. 2017, 184, 61–66. [Google Scholar] [CrossRef]
- Sadhu, S.; Mitra, D.K. Emerging Concepts of Adaptive Immunity in Leprosy. Front. Immunol. 2018, 9, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clerici, M.; Shearer, G.M. A Th1→Th2 switch is a critical step in the etiology of HIV infection. Immunol. Today 1993, 14, 107–111. [Google Scholar] [CrossRef]
- Calarota, S.A.; Weiner, D.B. Enhancement of human immunodeficiency virus type 1-DNA vaccine potency through incorporation of T-helper 1 molecular adjuvants. Immunol. Rev. 2004, 199, 84–99. [Google Scholar] [CrossRef] [PubMed]
- Saravia, J.; Chapman, N.M.; Chi, H. Helper T cell differentiation. Cell. Mol. Immunol. 2019, 16, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.Y. Multi-tasking of helper T cells. Immunology 2010, 130, 166–171. [Google Scholar] [CrossRef]
- Hsieh, C.S.; Macatonia, S.E.; Tripp, C.S.; Wolf, S.F.; O’Garra, A.; Murphy, K.M. Development of Th1 CD4+ t cells through IL-12 produced by listeria-induced macrophages. Science 1993, 260, 547–549. [Google Scholar] [CrossRef]
- Seder, R.A.; Gazzinelli, R.; Sher, A.; Paul, W.E. Interleukin 12 acts directly on CD4+ t cells to enhance priming for interferon gamma production and diminishes interleukin 4 inhibition of such priming. Proc. Natl. Acad. Sci. USA 1993, 90, 10188–10192. [Google Scholar] [CrossRef] [Green Version]
- Wenner, C.A.; Guler, M.L.; Macatonia, S.E.; O’Garra, A.; Murphy, K.M. Roles of ifn-gamma and ifn-alpha in IL-12-induced t helper cell-1 development. J. Immunol. 1996, 156, 1442–1447. [Google Scholar]
- Szabo, S.J.; Kim, S.T.; Costa, G.L.; Zhang, X.; Fathman, C.G.; Glimcher, L.H. A novel transcription factor, t-bet, directs Th1 lineage commitment. Cell 2000, 100, 655–669. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.L.; Weinberg, A.D.; English, M.; Huston, G. IL-4 directs the development of Th2-like helper effectors. J. Immunol. 1990, 145, 3796–3806. [Google Scholar]
- Le Gros, G.; Ben-Sasson, S.Z.; Seder, R.; Finkelman, F.D.; Paul, W.E. Generation of interleukin 4 (IL-4)-producing cells in vivo and in vitro: IL-2 and IL-4 are required for in vitro generation of IL-4-producing cells. J. Exp. Med. 1990, 172, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Parronchi, P.; Macchia, D.; Piccinni, M.P.; Biswas, P.; Simonelli, C.; Maggi, E.; Ricci, M.; Ansari, A.A.; Romagnani, S. Allergen- and bacterial antigen-specific t-cell clones established from atopic donors show a different profile of cytokine production. Proc. Natl. Acad. Sci. USA 1991, 88, 4538–4542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, S.Y.; Truitt, M.L.; Ho, I.C. Gata-3 deficiency abrogates the development and maintenance of t helper type 2 cells. Proc. Natl. Acad. Sci. USA 2004, 101, 1993–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Min, B.; Hu-Li, J.; Watson, C.J.; Grinberg, A.; Wang, Q.; Killeen, N.; Urban, J.F., Jr.; Guo, L.; Paul, W.E. Conditional deletion of gata3 shows its essential function in t(h)1-t(h)2 responses. Nat. Immunol. 2004, 5, 1157–1165. [Google Scholar] [CrossRef]
- Abers, M.S.; Delmonte, O.M.; Ricotta, E.E.; Fintzi, J.; Fink, D.L.; de Jesus, A.A.A.; Zarember, K.A.; Alehashemi, S.; Oikonomou, V.; Desai, J.V.; et al. An immune-based biomarker signature is associated with mortality in COVID-19 patients. JCI Insight 2021, 6, e144455. [Google Scholar] [CrossRef]
- Vabret, N.; Britton, G.J.; Gruber, C.; Hegde, S.; Kim, J.; Kuksin, M.; Levantovsky, R.; Malle, L.; Moreira, A.; Park, M.D.; et al. Immunology of COVID-19: Current state of the science. Immunity 2020, 52, 910–941. [Google Scholar] [CrossRef]
- Lucas, C.; Wong, P.; Klein, J.; Castro, T.B.R.; Silva, J.; Sundaram, M.; Ellingson, M.K.; Mao, T.; Oh, J.E.; Israelow, B.; et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 2020, 584, 463–469. [Google Scholar] [CrossRef]
- Martin-Sanchez, E.; Garces, J.J.; Maia, C.; Inoges, S.; Lopez-Diaz de Cerio, A.; Carmona-Torre, F.; Marin-Oto, M.; Alegre, F.; Molano, E.; Fernandez-Alonso, M.; et al. Immunological biomarkers of fatal COVID-19: A study of 868 patients. Front. Immunol. 2021, 12, 659018. [Google Scholar] [CrossRef]
- Weiskopf, D.; Schmitz, K.S.; Raadsen, M.P.; Grifoni, A.; Okba, N.M.A.; Endeman, H.; van den Akker, J.P.C.; Molenkamp, R.; Koopmans, M.P.G.; van Gorp, E.C.M.; et al. Phenotype and kinetics of SARS-CoV-2-specific t cells in COVID-19 patients with acute respiratory distress syndrome. Sci. Immunol. 2020, 5, eabd2071. [Google Scholar] [CrossRef]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Stralin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust t cell immunity in convalescent individuals with asymptomatic or mild COVID-19. Cell 2020, 183, 158–168.e14. [Google Scholar] [CrossRef]
- Roncati, L.; Nasillo, V.; Lusenti, B.; Riva, G. Signals of Th2 immune response from COVID-19 patients requiring intensive care. Ann. Hematol. 2020, 99, 1419–1420. [Google Scholar] [CrossRef] [PubMed]
- Gil-Etayo, F.J.; Suarez-Fernandez, P.; Cabrera-Marante, O.; Arroyo, D.; Garcinuno, S.; Naranjo, L.; Pleguezuelo, D.E.; Allende, L.M.; Mancebo, E.; Lalueza, A.; et al. T-helper cell subset response is a determining factor in COVID-19 progression. Front. Cell. Infect. Microbiol. 2021, 11, 624483. [Google Scholar] [CrossRef] [PubMed]
- Parrot, T.; Gorin, J.B.; Ponzetta, A.; Maleki, K.T.; Kammann, T.; Emgard, J.; Perez-Potti, A.; Sekine, T.; Rivera-Ballesteros, O.; Gredmark-Russ, S.; et al. Mait cell activation and dynamics associated with COVID-19 disease severity. Sci. Immunol. 2020, 5, eabe1670. [Google Scholar] [CrossRef] [PubMed]
- Jouan, Y.; Guillon, A.; Gonzalez, L.; Perez, Y.; Boisseau, C.; Ehrmann, S.; Ferreira, M.; Daix, T.; Jeannet, R.; Francois, B.; et al. Phenotypical and functional alteration of unconventional t cells in severe COVID-19 patients. J. Exp. Med. 2020, 217, e20200872. [Google Scholar] [CrossRef]
- Lombardi, A.; Trombetta, E.; Cattaneo, A.; Castelli, V.; Palomba, E.; Tirone, M.; Mangioni, D.; Lamorte, G.; Manunta, M.; Prati, D.; et al. Early phases of COVID-19 are characterized by a reduction in lymphocyte populations and the presence of atypical monocytes. Front. Immunol. 2020, 11, 560330. [Google Scholar] [CrossRef]
- Osman, M.; Faridi, R.M.; Sligl, W.; Shabani-Rad, M.T.; Dharmani-Khan, P.; Parker, A.; Kalra, A.; Tripathi, M.B.; Storek, J.; Cohen Tervaert, J.W.; et al. Impaired natural killer cell counts and cytolytic activity in patients with severe COVID-19. Blood Adv. 2020, 4, 5035–5039. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine storm in COVID-19: The current evidence and treatment strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef]
- Fajgenbaum, D.C.; June, C.H. Cytokine storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef]
- Petrey, A.C.; Qeadan, F.; Middleton, E.A.; Pinchuk, I.V.; Campbell, R.A.; Beswick, E.J. Cytokine release syndrome in COVID-19: Innate immune, vascular, and platelet pathogenic factors differ in severity of disease and sex. J. Leukoc. Biol. 2021, 109, 55–66. [Google Scholar] [CrossRef]
- Martinez-Feito, A.; Melero, J.; Mora-Diaz, S.; Rodriguez-Vigil, C.; Elduayen, R.; Gonzalez-Granado, L.I.; Perez-Mendez, D.; Sanchez-Zapardiel, E.; Ruiz-Garcia, R.; Menchen, M.; et al. Autoimmune lymphoproliferative syndrome due to somatic fas mutation (alps-sfas) combined with a germline caspase-10 (casp10) variation. Immunobiology 2016, 221, 40–47. [Google Scholar] [CrossRef]
- Zeng, H.; Ma, Y.; Zhou, Z.; Liu, W.; Huang, P.; Jiang, M.; Liu, Q.; Chen, P.; Luo, H.; Chen, Y. Spectrum and clinical characteristics of symptomatic and asymptomatic coronavirus disease 2019 (COVID-19) with and without pneumonia. Front. Med. 2021, 8, 645651. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Wang, Q.; Zhang, D.; Ding, J.; Huang, Q.; Tang, Y.Q.; Miao, H. Lymphopenia predicts disease severity of COVID-19: A descriptive and predictive study. Signal Transduct. Target. Ther. 2020, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Reusch, N.; De Domenico, E.; Bonaguro, L.; Schulte-Schrepping, J.; Bassler, K.; Schultze, J.L.; Aschenbrenner, A.C. Neutrophils in COVID-19. Front. Immunol. 2021, 12, 652470. [Google Scholar] [CrossRef] [PubMed]
- Lindsley, A.W.; Schwartz, J.T.; Rothenberg, M.E. Eosinophil responses during COVID-19 infections and coronavirus vaccination. J. Allergy Clin. Immunol 2020, 146, 1–7. [Google Scholar] [CrossRef] [PubMed]
- San Segundo, D.; Arnaiz de Las Revillas, F.; Lamadrid-Perojo, P.; Comins-Boo, A.; Gonzalez-Rico, C.; Alonso-Pena, M.; Irure-Ventura, J.; Olmos, J.M.; Farinas, M.C.; Lopez-Hoyos, M. Innate and adaptive immune assessment at admission to predict clinical outcome in COVID-19 patients. Biomedicines 2021, 9, 917. [Google Scholar] [CrossRef] [PubMed]
- De Biasi, S.; Meschiari, M.; Gibellini, L.; Bellinazzi, C.; Borella, R.; Fidanza, L.; Gozzi, L.; Iannone, A.; Lo Tartaro, D.; Mattioli, M.; et al. Marked t cell activation, senescence, exhaustion and skewing towards Th17 in patients with COVID-19 pneumonia. Nat. Commun. 2020, 11, 3434. [Google Scholar] [CrossRef]
- Infante-Duarte, C.; Kamradt, T. Th1/Th2 balance in infection. Springer Semin. Immunopathol. 1999, 21, 317–338. [Google Scholar] [CrossRef]
- Rowland-Jones, S.; Tan, R.; McMichael, A. Role of cellular immunity in protection against hiv infection. Adv. Immunol. 1997, 65, 277–346. [Google Scholar]
- Kaul, R.; Rowland-Jones, S.L.; Kimani, J.; Fowke, K.; Dong, T.; Kiama, P.; Rutherford, J.; Njagi, E.; Mwangi, F.; Rostron, T.; et al. New insights into hiv-1 specific cytotoxic t-lymphocyte responses in exposed, persistently seronegative kenyan sex workers. Immunol. Lett. 2001, 79, 3–13. [Google Scholar] [CrossRef]
- Unim, B.; Palmieri, L.; Lo Noce, C.; Brusaferro, S.; Onder, G. Prevalence of COVID-19-related symptoms by age group. Aging Clin. Exp. Res. 2021, 33, 1145–1147. [Google Scholar] [CrossRef]
- Davies, N.G.; Klepac, P.; Liu, Y.; Prem, K.; Jit, M.; Eggo, R.M. Age-dependent effects in the transmission and control of COVID-19 epidemics. Nat. Med. 2020, 26, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Parcha, V.; Booker, K.S.; Kalra, R.; Kuranz, S.; Berra, L.; Arora, G.; Arora, P. A retrospective cohort study of 12,306 pediatric COVID-19 patients in the united states. Sci. Rep. 2021, 11, 10231. [Google Scholar] [CrossRef] [PubMed]
- Josa-Laorden, C.; Crestelo-Vieitez, A.; Garcia Andreu, M.D.M.; Rubio-Rivas, M.; Sanchez, M.; Toledo Samaniego, N.; Arnalich Fernandez, F.; Iguaran Bermudez, R.; Fonseca Aizpuru, E.M.; Vargas Nunez, J.A.; et al. Gender-based differences by age range in patients hospitalized with COVID-19: A spanish observational cohort study. J. Clin. Med. 2021, 10, 899. [Google Scholar] [CrossRef] [PubMed]
- Laguna-Goya, R.; Utrero-Rico, A.; Talayero, P.; Lasa-Lazaro, M.; Ramirez-Fernandez, A.; Naranjo, L.; Segura-Tudela, A.; Cabrera-Marante, O.; Rodriguez de Frias, E.; Garcia-Garcia, R.; et al. IL-6-based mortality risk model for hospitalized patients with COVID-19. J. Allergy Clin. Immunol. 2020, 146, 799–807.e9. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Park, S.S.; Kim, T.Y.; Lee, D.G.; Kim, D.W. Lymphopenia as a biological predictor of outcomes in COVID-19 patients: A nationwide cohort study. Cancers 2021, 13, 471. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Bautista, J.F.; Rodriguez-Nicolas, A.; Rosales-Castillo, A.; Jimenez, P.; Garrido, F.; Anderson, P.; Ruiz-Cabello, F.; Lopez-Ruz, M.A. Negative clinical evolution in COVID-19 patients is frequently accompanied with an increased proportion of undifferentiated th cells and a strong underrepresentation of the Th1 subset. Front. Immunol. 2020, 11, 596553. [Google Scholar] [CrossRef]
- Mahmoud Salehi Khesht, A.; Karpisheh, V.; Qubais Saeed, B.; Olegovna Zekiy, A.; Yapanto, L.M.; Nabi Afjadi, M.; Aksoun, M.; Nasr Esfahani, M.; Aghakhani, F.; Movahed, M.; et al. Different t cell related immunological profiles in COVID-19 patients compared to healthy controls. Int. Immunopharmacol. 2021, 97, 107828. [Google Scholar] [CrossRef]
- Salkowska, A.; Karwaciak, I.; Karas, K.; Dastych, J.; Ratajewski, M. SARS-CoV-2 proteins induce ifng in Th1 lymphocytes generated from CD4+ cells from healthy, unexposed polish donors. Vaccines 2020, 8, 673. [Google Scholar] [CrossRef]
- Thakur, A.; Mikkelsen, H.; Jungersen, G. Intracellular pathogens: Host immunity and microbial persistence strategies. J. Immunol. Res. 2019, 2019, 1356540. [Google Scholar] [CrossRef]
- Mortaz, E.; Tabarsi, P.; Varahram, M.; Folkerts, G.; Adcock, I.M. The immune response and immunopathology of COVID-19. Front. Immunol. 2020, 11, 2037. [Google Scholar] [CrossRef]
- Zhao, Y.; Kilian, C.; Turner, J.E.; Bosurgi, L.; Roedl, K.; Bartsch, P.; Gnirck, A.C.; Cortesi, F.; Schultheiss, C.; Hellmig, M.; et al. Clonal expansion and activation of tissue-resident memory-like Th17 cells expressing gm-csf in the lungs of severe COVID-19 patients. Sci. Immunol. 2021, 6, eabf6692. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.Y. T(h)1 and t(h)2 cells: A historical perspective. Nat. Rev. Immunol. 2002, 2, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Forthal, D.N. Functions of antibodies. Microbiol. Spectr. 2014, 2, 2–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guglani, L.; Khader, S.A. Th17 cytokines in mucosal immunity and inflammation. Curr. Opin. HIV AIDS 2010, 5, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Martonik, D.; Parfieniuk-Kowerda, A.; Rogalska, M.; Flisiak, R. The role of Th17 response in COVID-19. Cells 2021, 10, 1550. [Google Scholar] [CrossRef]
- Pasrija, R.; Naime, M. The deregulated immune reaction and cytokines release storm (crs) in COVID-19 disease. Int. Immunopharmacol. 2021, 90, 107225. [Google Scholar] [CrossRef]
- Croci, S.; Bonacini, M.; Dolci, G.; Massari, M.; Facciolongo, N.; Pignatti, E.; Pisciotta, A.; Carnevale, G.; Negro, A.; Cassone, G.; et al. Human dental pulp stem cells modulate cytokine production in vitro by peripheral blood mononuclear cells from coronavirus disease 2019 patients. Front. Cell Dev. Biol. 2020, 8, 609204. [Google Scholar] [CrossRef]
- Sarmiento-Monroy, J.C.; Parra-Medina, R.; Garavito, E.; Rojas-Villarraga, A. T helper 17 response to severe acute respiratory syndrome coronavirus 2: A type of immune response with possible therapeutic implications. Viral Immunol. 2021, 34, 190–200. [Google Scholar] [CrossRef]
- Chan, Y.H.; Fong, S.W.; Poh, C.M.; Carissimo, G.; Yeo, N.K.; Amrun, S.N.; Goh, Y.S.; Lim, J.; Xu, W.; Chee, R.S.; et al. Asymptomatic COVID-19: Disease tolerance with efficient anti-viral immunity against SARS-CoV-2. EMBO Mol. Med. 2021, 13, e14045. [Google Scholar] [CrossRef]
- Roberts, A.D.; Ely, K.H.; Woodland, D.L. Differential contributions of central and effector memory t cells to recall responses. J. Exp. Med. 2005, 202, 123–133. [Google Scholar] [CrossRef]
- Wang, F.; Hou, H.; Luo, Y.; Tang, G.; Wu, S.; Huang, M.; Liu, W.; Zhu, Y.; Lin, Q.; Mao, L.; et al. The laboratory tests and host immunity of COVID-19 patients with different severity of illness. JCI Insight 2020, 5, e137799. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Tian, F.B.; Xiang, D.D.; Yu, B. Characteristics of inflammatory factors and lymphocyte subsets in patients with severe COVID-19. J. Med. Virol. 2020, 92, 2600–2606. [Google Scholar] [CrossRef] [PubMed]
- Kusnadi, A.; Ramirez-Suastegui, C.; Fajardo, V.; Chee, S.J.; Meckiff, B.J.; Simon, H.; Pelosi, E.; Seumois, G.; Ay, F.; Vijayanand, P.; et al. Severely ill COVID-19 patients display impaired exhaustion features in SARS-CoV-2-reactive CD8(+) t cells. Sci. Immunol. 2021, 6, eabe4782. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Gao, Y.; Wang, G.; Song, G.; Liu, S.; Sun, D.; Xu, Y.; Tian, Z. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cell. Mol. Immunol. 2020, 17, 533–535. [Google Scholar] [CrossRef] [Green Version]
- Ganneru, B.; Jogdand, H.; Daram, V.K.; Das, D.; Molugu, N.R.; Prasad, S.D.; Kannappa, S.V.; Ella, K.M.; Ravikrishnan, R.; Awasthi, A.; et al. Th1 skewed immune response of whole virion inactivated SARS-CoV-2 vaccine and its safety evaluation. iScience 2021, 24, 102298. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine bnt162b1 elicits human antibody and Th1 t cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- Almendro-Vazquez, P.; Laguna-Goya, R.; Ruiz-Ruigomez, M.; Utrero-Rico, A.; Lalueza, A.; Maestro de la Calle, G.; Delgado, P.; Perez-Ordono, L.; Muro, E.; Vila, J.; et al. Longitudinal dynamics of SARS-CoV-2-specific cellular and humoral immunity after natural infection or BNT162b2 vaccination. PLoS Pathog. 2021, 17, e1010211. [Google Scholar] [CrossRef]
- Korsholm, K.S.; Petersen, R.V.; Agger, E.M.; Andersen, P. T-helper 1 and t-helper 2 adjuvants induce distinct differences in the magnitude, quality and kinetics of the early inflammatory response at the site of injection. Immunology 2010, 129, 75–86. [Google Scholar] [CrossRef]
- Firacative, C.; Gressler, A.E.; Schubert, K.; Schulze, B.; Muller, U.; Brombacher, F.; von Bergen, M.; Alber, G. Identification of t helper (th)1- and Th2-associated antigens of cryptococcus neoformans in a murine model of pulmonary infection. Sci. Rep. 2018, 8, 2681. [Google Scholar] [CrossRef]
- Maeyama, J.; Takatsuka, H.; Suzuki, F.; Kubota, A.; Horiguchi, S.; Komiya, T.; Shimada, I.; Murata, E.; Osawa, Y.; Kitagawa, H.; et al. A palindromic cpg-containing phosphodiester oligodeoxynucleotide as a mucosal adjuvant stimulates plasmacytoid dendritic cell-mediated t(h)1 immunity. PLoS ONE 2014, 9, e88846. [Google Scholar] [CrossRef]
- Safar, H.A.; Mustafa, A.S.; Amoudy, H.A.; El-Hashim, A. The effect of adjuvants and delivery systems on Th1, Th2, Th17 and treg cytokine responses in mice immunized with mycobacterium tuberculosis-specific proteins. PLoS ONE 2020, 15, e0228381. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Non-Hospitalized; n = 54 | Hospitalized; n = 92 | p-Value |
|---|---|---|---|
| n(%)/Median (IQR) | n(%)/Median (IQR) | ||
| Age | 48.5 (39–63) | 58.5 (46–72) | 0.004 |
| Sex (male) | 29 (54%) | 58 (63%) | 0.038 |
| LDH | 276.5 (222.5–310) | 336 (225.5–402.2) | 0.001 |
| CRP | 2.9 (1.3–5.2) | 6 (3.7–11) | <0.001 |
| DD | 555 (299.5–907.5) | 691 (410–1414) | 0.058 |
| Healthy Control; n = 29 | COVID-19 Cohort; n = 146 | ||||
|---|---|---|---|---|---|
| Variables | Median | IQR | Median | IQR | p-Value |
| Lymphocytes | 1846 | 1585–2129 | 1100 | 800–1500 | < 0.001 |
| CD4 count | 783 | 664–1099 | 636 | 416–708 | 0.012 |
| %CD4 Naïve | 35 | 29.32–46.65 | 30.3 | 18.8–38.1 | 0.028 |
| %CD4 CM | 41 | 30.22–46 | 36.7 | 27.97–46.2 | 0.499 |
| %CD4 EM | 18.3 | 13.6–24.9 | 25.8 | 18.6–35.2 | 0.001 |
| %CD4 TEMRA | 0.7 | 0.3–1.92 | 1.5 | 0.5–4 | 0.087 |
| %CD4 activation | 4.1 | 3.47–5.52 | 6.2 | 3.5–9.5 | 0.023 |
| %CD4 senescence | 7.4 | 4.85–12.9 | 5.5 | 2.9–10.1 | 0.112 |
| %CD4 double positive | 0.8 | 0.5–1.17 | 1 | 0.4–2.4 | 0.505 |
| CD8 count | 404 | 285–524 | 296 | 185–477 | 0.022 |
| %CD8 Naïve | 27.6 | 17.8–41.12 | 13 | 5.5–23 | <0.001 |
| %CD8 CM | 13.7 | 8.37–18.7 | 8.3 | 5.1–11.2 | <0.001 |
| %CD8 EM | 42 | 26.6–49.9 | 48.1 | 40–60.2 | <0.001 |
| %CD8 TEMRA | 10 | 7.52–24.5 | 20.5 | 11–30.5 | 0.003 |
| %CD8 activation | 9.5 | 6–12.6 | 18 | 9.1–26 | <0.001 |
| %CD8 senescence | 26.2 | 20–35.9 | 38.7 | 18.75–50 | 0.024 |
| %CD8 double positive | 4.5 | 2.4–8.32 | 6 | 2.67–12.95 | 0.08 |
| Condition (All Patients) | Hospitalized (n = 92) | Not Hospitalized (n = 54) | p Value | OR | 95% CI | ||
|---|---|---|---|---|---|---|---|
| Patients with comorbidities | 51 | (55.4%) | 22 | (40.7%) | 0.0865 | ||
| Myocardial infarction | 0 | (0%) | 1 | (1.9%) | 0.3699 | ||
| Diabetes mellitus | 18 | (19.6%) | 7 | (13%) | 0.3066 | ||
| Advanzed chronical kidney disease | 0 | (0%) | 2 | (3.7%) | 0.1352 | ||
| Active smokers | 3 | (3.3%) | 4 | (7.4%) | 0.4239 | ||
| Former smokers | 12 | (13%) | 3 | (5.6%) | 0.1718 | ||
| Obesity | 16 | (17.4%) | 4 | (7.4%) | 0.1336 | ||
| Dyslipidemia | 23 | (25%) | 9 | (16.7%) | 0.2416 | ||
| Hypertension | 27 | (29.3%) | 13 | (24.1%) | 0.4903 | ||
| Condition (Hospitalized Patients) | ICU Treated (n = 21) | Not ICU (n = 71) | |||||
| Patients with comorbidities | 16 | (76.2%) | 35 | (49.3%) | 0.0303 | 3.29 | 1.09–9.95 |
| Myocardial infarction | 0 | 0 | |||||
| Diabetes mellitus | 5 | (23.8%) | 13 | (18.3%) | 0.5768 | ||
| Advanzed chronical kidney disease | 0 | 0 | |||||
| Active smokers | 2 | (9.5%) | 1 | (1.4%) | 0.1293 | ||
| Former smokers | 3 | (14.3%) | 9 | (12.7%) | 1.0 | ||
| Obesity | 7 | (33.3%) | 9 | (12.7%) | 0.0291 | 3.44 | 1.10–10.83 |
| Dyslipidemia | 7 | (33.3%) | 16 | (22.5%) | 0.3154 | ||
| Hypertension | 9 | (42.9%) | 18 | (25.4%) | 0.1217 | ||
| Variables | Univariate | Multivariate | ||||
|---|---|---|---|---|---|---|
| OR | OR 95% CI | p-Value | OR | OR 95% CI | p-Value | |
| (A) NH vs. H | ||||||
| Sex (male) | 2.13 | 1.07–4.22 | 0.03 | 1.3 | 0.54–3.11 | 0.544 |
| Age | 1.03 | 1–1.04 | 0.006 | 1.02 | 0.9–1.04 | 0.077 |
| Lymphocytes | 0.3 | 0.13–0.65 | 0.003 | 0.47 | 0.18–1.23 | 0.126 |
| %CD4 activation | 2.87 | 1.3–5.94 | 0.005 | 2.53 | 0.94–6.82 | 0.066 |
| %CD8 activation | 2.42 | 1.14–5.12 | 0.02 | 1.41 | 0.48–4.07 | 0.526 |
| %Th1 | 0.3 | 0.11–0.78 | 0.014 | 0.18 | 0.04–0.75 | 0.018 |
| LDH | 1 | 1–1.1 | 0.006 | 1.1 | 1–1.1 | 0.016 |
| CRP | 1.1 | 1–1.14 | 0.032 | 1 | 0.93–1.07 | 0.956 |
| Area Under the ROC Curve | 0.802 | (0.723–0.867) | ||||
| (B) NS vs. S | ||||||
| Age | 1.02 | 1–1.04 | 0.023 | 1.02 | 0.99–1.05 | 0.069 |
| Comorbidity | 2.94 | 1.39–6.2 | 0.004 | 2.57 | 0.9–7.35 | 0.076 |
| Obesity | 3.44 | 1.31–9.05 | 0.012 | 2.08 | 0.67–6.42 | 0.199 |
| Lymphocytes | 0.4 | 0.19–0.86 | 0.019 | 0.49 | 0.2–1.23 | 0.131 |
| %Th1 quiescence | 0.37 | 0.17–0.80 | 0.012 | 0.34 | 0.13–0.85 | 0.022 |
| %Th2 late-activation | 3.41 | 1.24–9.38 | 0.017 | 5.71 | 1.65–19.74 | 0.005 |
| %Th17 quiescence | 2.3 | 1.01–5.24 | 0.046 | 2.9 | 1.02–8.23 | 0.045 |
| Area Under the ROC Curve | 0.801 | (0.727–0.863) | ||||
| (C) NH vs. M | ||||||
| Age | 1.01 | 0.99–1.04 | 0.072 | 1.01 | 0.99–1.04 | 0.199 |
| Lymphocytes | 0.37 | 0.15– 0.89 | 0.026 | 0.94 | 0.28–2.83 | 0.086 |
| CD4 count | 0.44 | 0.26–0.75 | 0.002 | 0.62 | 0.31–1.25 | 0.185 |
| %CD4 activation | 2.8 | 1.3–6.08 | 0.008 | 2.58 | 1.12–2.95 | 0.026 |
| %Th1 | 0.36 | 0.12–1.12 | 0.078 | 0.32 | 0.8–1.23 | 0.099 |
| Area Under the ROC Curve | 0.762 | (0.334–0.841) | ||||
| (D) NH vs. S | ||||||
| Comorbidity | 3.11 | 1.35–7.18 | 0.007 | 3.6 | 1.21–10.71 | 0.021 |
| Obesity | 4.16 | 1.22–14.19 | 0.022 | 4.48 | 0.81–25 | 0.086 |
| Lymphocytes | 0.23 | 0.089–0.63 | 0.004 | 0.24 | 0.07–0.73 | 0.012 |
| %CD8 activation | 4.14 | 1.75–9.83 | 0.001 | 5.91 | 1.93–18.03 | 0.0018 |
| %Th1 | 0.23 | 0.06–0.87 | 0.03 | 0.17 | 0.03–0.81 | 0.025 |
| %Th17 early-activation | 6.79 | 0.76–60.5 | 0.085 | 1.53 | 0.13–16.93 | 0.725 |
| Area Under the ROC Curve | 0.844 | (0.756–0.909) | ||||
| (E) M vs. S | ||||||
| Comrbidity | 2.75 | 1.17–6.46 | 0.019 | 5.08 | 1.54–16.75 | 0.007 |
| %CD4 senescence | 0.33 | 0.09–1.14 | 0.082 | 0.11 | 0.02–0.64 | 0.014 |
| %CD8 activation | 2.89 | 1.23–6.79 | 0.014 | 2.53 | 0.54–11.92 | 0.237 |
| %CD8 double positive | 1.09 | 1.03–1.16 | 0.004 | 1.12 | 0.99–1.27 | 0.052 |
| %Th1 quiescence | 0.43 | 0.18–1.03 | 0.061 | 0.27 | 0.08–0.86 | 0.027 |
| LDH | 1 | 1–1.01 | 0.008 | 1 | 1–1.01 | 0.013 |
| Area Under the ROC Curve | 0.86 | (0.770–0.925) | ||||
| (F) A vs. SY | ||||||
| %CD4 CD57+ | 0.23 | 0.03–1.33 | 0.12 | 0.31 | 0.04–2.07 | 0.231 |
| %Th1 | 0.2 | 0.049–0.8 | 0.023 | 0.23 | 0.05–0.96 | 0.045 |
| Area Under the ROC Curve | 0.697 | (0.557–0.815) | ||||
| (G) A vs. S | ||||||
| Lymphocytes | 0.18 | 0.04–0.81 | 0.025 | 0.38 | 0.07–1.95 | 0.247 |
| %CD8 activation | 6.57 | 1.28–33.61 | 0.023 | 6.67 | 1.05–45.6 | 0.044 |
| %Th1 | 0.07 | 0.01–0.37 | 0.002 | 0.09 | 0.01–0.63 | 0.015 |
| Area Under the ROC Curve | 0.837 | (0.714–0.922) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil-Etayo, F.J.; Garcinuño, S.; Utrero-Rico, A.; Cabrera-Marante, O.; Arroyo-Sanchez, D.; Mancebo, E.; Pleguezuelo, D.E.; Rodríguez-Frías, E.; Allende, L.M.; Morales-Pérez, P.; et al. An Early Th1 Response Is a Key Factor for a Favorable COVID-19 Evolution. Biomedicines 2022, 10, 296. https://doi.org/10.3390/biomedicines10020296
Gil-Etayo FJ, Garcinuño S, Utrero-Rico A, Cabrera-Marante O, Arroyo-Sanchez D, Mancebo E, Pleguezuelo DE, Rodríguez-Frías E, Allende LM, Morales-Pérez P, et al. An Early Th1 Response Is a Key Factor for a Favorable COVID-19 Evolution. Biomedicines. 2022; 10(2):296. https://doi.org/10.3390/biomedicines10020296
Chicago/Turabian StyleGil-Etayo, Francisco Javier, Sara Garcinuño, Alberto Utrero-Rico, Oscar Cabrera-Marante, Daniel Arroyo-Sanchez, Esther Mancebo, Daniel Enrique Pleguezuelo, Edgard Rodríguez-Frías, Luis M. Allende, Pablo Morales-Pérez, and et al. 2022. "An Early Th1 Response Is a Key Factor for a Favorable COVID-19 Evolution" Biomedicines 10, no. 2: 296. https://doi.org/10.3390/biomedicines10020296