
Investigating the Antibacterial Properties of Prospective Scabicides

Abstract
:1. Introduction
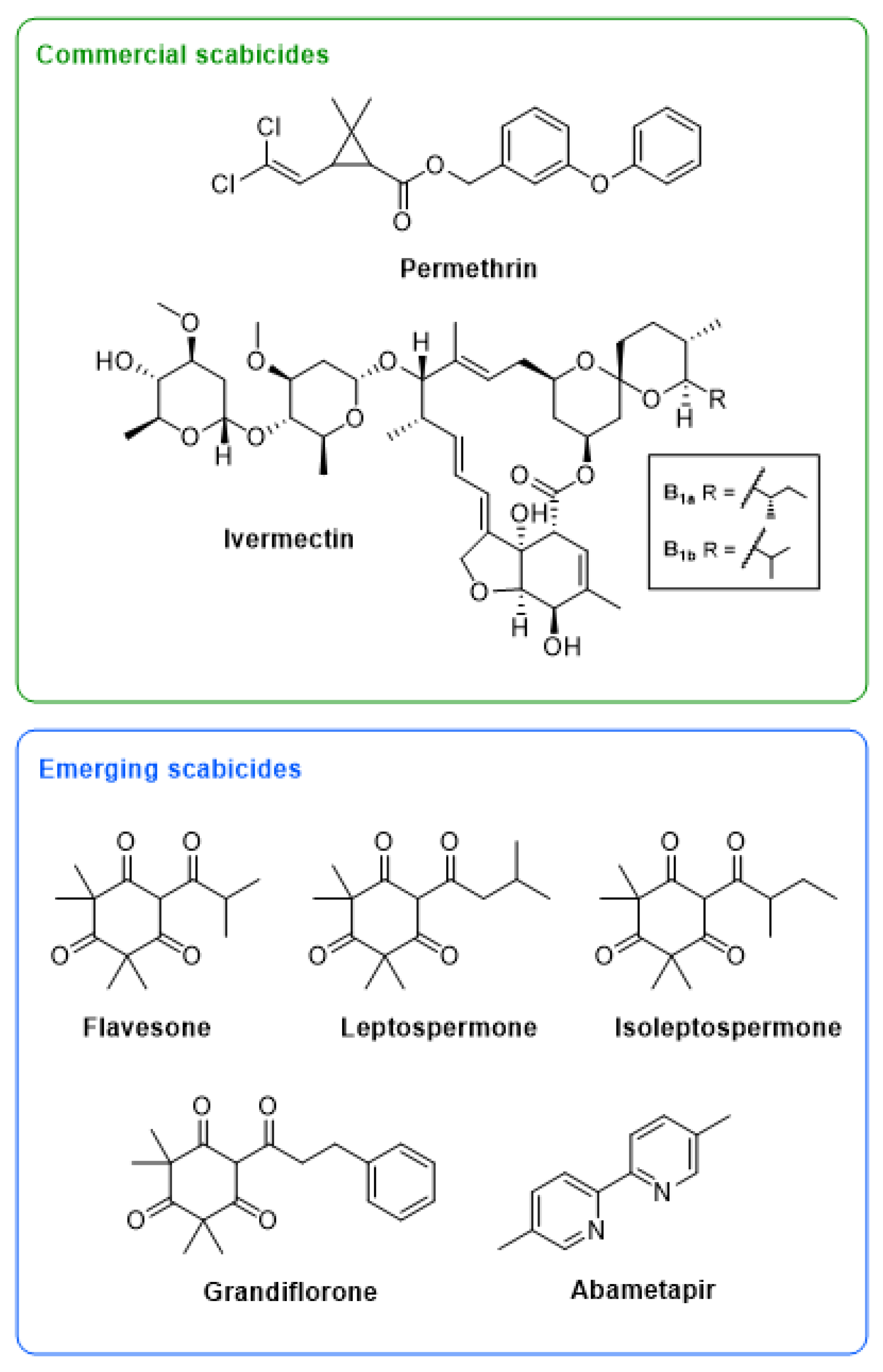
1.1. Emerging Scabicides
1.2. The Importance of Anti-Bacterial Activity in Addition to Acaricidal Properties in a Scabies Treatment
2. Materials and Methods
2.1. Synthesis
2.1.1. General Synthesis of Acylphloroglucinols
2.1.2. General Synthesis of β-Triketones
2.2. Drug Dilution Details for Antimicrobial Testing
2.3. Bacterial Strains and Growth Conditions
2.4. Statistical Analysis
3. Results
3.1. Antibacterial Activity of Mānuka Oil and Its β-Triketones
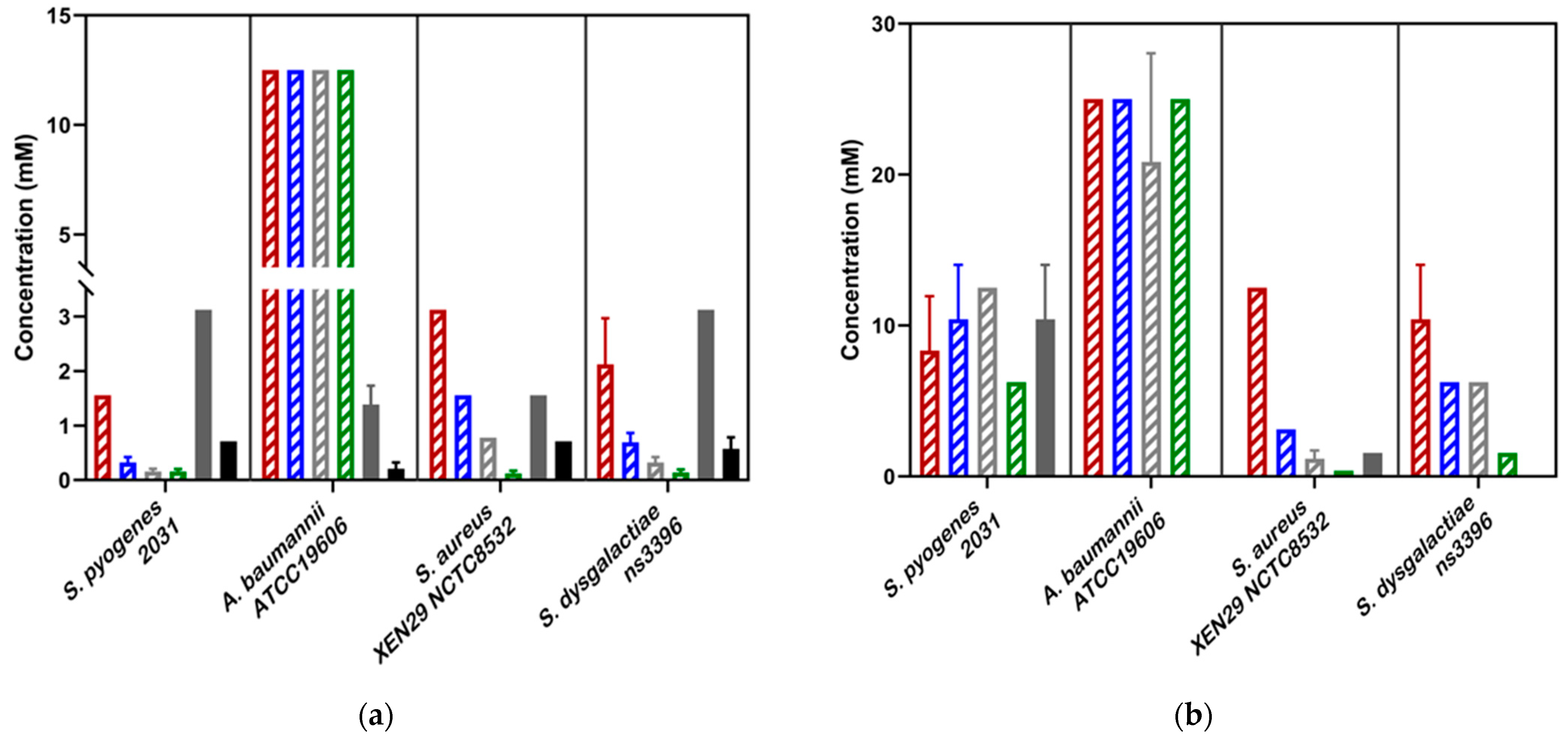
3.2. Antibacterial Activity of Abametapir and Ivermectin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, S.L.; Collinson, S.; Timothy, J.; Zayzay, S.K.; Kollie, K.K.; Candy, N.; Lebas, E.; Halliday, K.; Pullan, R.; Fallah, M.; et al. A community-based validation of the International Alliance for the Control of Scabies Consensus Criteria by expert and non-expert examiners in Liberia. PLoS Neglected Trop. Dis. 2020, 14, e0008717. [Google Scholar] [CrossRef] [PubMed]
- Stephens, J.M.C.; Molan, P.C.; Clarkson, B.D. A review of Leptospermum scoparium (Myrtaceae) in New Zealand. New Zealand J. Bot. 2005, 43, 431–449. [Google Scholar] [CrossRef]
- Bernigaud, C.; Samarawickrama, G.R.; Jones, M.K.; Gasser, R.B.; Fischer, K. The Challenge of Developing a Single-Dose Treatment for Scabies. Trends Parasitol. 2019, 35, 931–943. [Google Scholar] [CrossRef]
- Bernigaud, C.; Fernando, D.D.; Lu, H.; Taylor, S.; Hartel, G.; Guillot, J.; Chosidow, O.; Fischer, K. In Vitro ovicidal activity of current and under-development scabicides: Which treatments kill scabies eggs? Br. J. Dermatol. 2020, 182, 511–513. [Google Scholar] [CrossRef]
- Mounsey, K.; Ho, M.-F.; Kelly, A.; Willis, C.; Pasay, C.; Kemp, D.J.; McCarthy, J.; Fischer, K. A Tractable Experimental Model for Study of Human and Animal Scabies. PLoS Neglected Trop. Dis. 2010, 4, e756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balestri, R.; Magnano, M.; Infusino, S.D.; Rizzoli, L.; Girardelli, C.R.; Rech, G. Scabies is becoming less sensitive to permethrin therapy. J. Eur. Acad. Dermatol. Venereol. 2021, 35, e889–e891. [Google Scholar] [CrossRef]
- Lynar, S.; Currie, B.J.; Baird, R. Scabies and mortality. Lancet Infect. Dis. 2017, 17, 1234. [Google Scholar] [CrossRef] [Green Version]
- Bernigaud, C.; Zakrzewski, M.; Taylor, S.; Swe, P.M.; Papenfuss, A.T.; Sriprakash, K.S.; Holt, D.; Chosidow, O.; Currie, B.J.; Fischer, K. First Description of the Composition and the Functional Capabilities of the Skin Microbial Community Accompanying Severe Scabies Infestation in Humans. Microorganisms 2021, 9, 907. [Google Scholar] [CrossRef]
- Asha Bowen, B.C.; Katzenellenbogen, J.; Marangou, J.; Noonan, S.; Ralph, A.; Roberts, K.; Steer, A.; Vaughan, G.; Wade, V.; Wyber, R. The 2020 Australian Guideline for Prevention, Diagnosis and Management of Acute Rheumatic Fever and Rheumatic Heart Disease; Bart Currie, A.R., Ed.; Menzies School of Health Research: Darwin, NT, Australia, 2022; p. 314. [Google Scholar]
- Thornley, S.; Marshall, R.; Jarrett, P.; Sundborn, G.; Reynolds, E.; Schofield, G. Scabies is strongly associated with acute rheumatic fever in a cohort study of Auckland children. J. Paediatr. Child Health 2018, 54, 625–632. [Google Scholar] [CrossRef]
- Bowles, V.M.; VanLuvanee, L.J.; Alsop, H.; Hazan, L.; Bs, K.S.; Sidgiddi, S.; Allenby, K.; Bn, T.A.; Hanegraaf, S. Clinical studies evaluating abametapir lotion, 0.74%, for the treatment of head louse infestation. Pediatr. Dermatol. 2018, 35, 616–621. [Google Scholar] [CrossRef]
- Bowles, V.M.; Young, A.R.; Barker, S.C. Metalloproteases and egg-hatching in Pediculus humanus, the body (clothes) louse of humans (Phthiraptera: Insecta). Parasitology 2007, 135, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.D.; Porter, C.L.; Feldman, S.R. Abametapir for the Treatment of Head Lice: A Drug Review. Ann. Pharmacother. 2021, 56, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Bowles, V.M.; Hanegraaf, S.; Ahveninen, T.; Sidgiddi, S.; Allenby, K.; Alsop, H. Effect of a New Head Lice Treatment, Abametapir Lotion, 0.74%, on Louse Eggs: A Randomized, Double-Blind Study. Glob. Pediatr. Health 2019, 6, 2333794x19831295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downs, A.M. Managing head lice in an era of increasing resistance to insecticides. Am. J. Clin. Dermatol. 2004, 5, 169–177. [Google Scholar] [CrossRef] [PubMed]
- McNair, C.M. Ectoparasites of medical and veterinary importance: Drug resistance and the need for alternative control methods. J. Pharm. Pharmacol. 2015, 67, 351–363. [Google Scholar] [CrossRef]
- Korhonen, P.K.; Gasser, R.B.; Ma, G.; Wang, T.; Stroehlein, A.J.; Young, N.D.; Ang, C.-S.; Fernando, D.D.; Lu, H.C.; Taylor, S.; et al. High-quality nuclear genome for Sarcoptes scabiei—A critical resource for a neglected parasite. PLoS Neglected Trop. Dis. 2020, 14, e0008720. [Google Scholar] [CrossRef]
- George, D.; Smith, T.; Shiel, R.; Sparagano, O.; Guy, J. Mode of action and variability in efficacy of plant essential oils showing toxicity against the poultry red mite, Dermanyssus gallinae. Vet. Parasitol. 2009, 161, 276–282. [Google Scholar] [CrossRef]
- Jeong, E.Y.; Kim, M.G.; Lee, H.S. Acaricidal activity of triketone analogues derived from Leptospermum scoparium oil against house-dust and stored-food mites. Pest Manag. Sci. 2009, 65, 327–331. [Google Scholar] [CrossRef]
- Muturi, E.J.; Ramirez, J.L.; Doll, K.; Bowman, M.J. Combined Toxicity of Three Essential Oils against Aedes aegypti (Diptera: Culicidae) Larvae. J. Med. Èntomol. 2017, 54, 1684–1691. [Google Scholar] [CrossRef]
- Park, C.; Jang, M.; Shin, E.; Kim, J. Myrtaceae Plant Essential Oils and their β-Triketone Components as Insecticides against Drosophila suzukii. Molecules 2017, 22, 1050. [Google Scholar] [CrossRef]
- Fang, F.; Candy, K.; Melloul, E.; Bernigaud, C.; Chai, L.; Darmon, C.; Durand, R.; Botterel, F.; Chosidow, O.; Izri, A.; et al. In Vitro activity of ten essential oils against Sarcoptes scabiei. Parasites Vectors 2016, 9, 594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, E.-Y.; Lee, M.-J.; Lee, H.-S. Antimicrobial activities of leptospermone isolated from Leptospermum scoparium seeds and structure–activity relationships of its derivatives against foodborne bacteria. Food Sci. Biotechnol. 2018, 27, 1541–1547. [Google Scholar] [CrossRef] [PubMed]
- Christoph, F.; Kaulfers, P.-M.; Stahl-Biskup, E. A Comparative Study of the in vitro Antimicrobial Activity of Tea Tree Oils s.l. with Special Reference to the Activity of β-Triketones. Planta Medica 2000, 66, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Harkenthal, M.; Reichling, J.; Geiss, H.K.; Saller, R. Comparative study on the in vitro antibacterial activity of Australian tea tree oil, cajuput oil, niaouli oil, manuka oil, kanuka oil, and eucalyptus oil. Die Pharm. 1999, 54, 460–463. [Google Scholar]
- Chen, C.-C.; Yan, S.-H.; Yen, M.-Y.; Wu, P.-F.; Liao, W.-T.; Huang, T.-S.; Wen, Z.-H.; Wang, H.-M.D. Investigations of kanuka and manuka essential oils for in vitro treatment of disease and cellular inflammation caused by infectious microorganisms. J. Microbiol. Immunol. Infect. 2016, 49, 104–111. [Google Scholar] [CrossRef] [Green Version]
- Mathew, C.; Tesfaye, W.; Rasmussen, P.; Peterson, G.M.; Bartholomaeus, A.; Sharma, M.; Thomas, J. Mānuka Oil—A Review of Antimicrobial and Other Medicinal Properties. Pharmaceuticals 2020, 13, 343. [Google Scholar] [CrossRef]
- Van Vuuren, S.F.; Docrat, Y.; Kamatou, G.P.P.; Viljoen, A.M. Essential oil composition and antimicrobial interactions of understudied tea tree species. S. Afr. J. Bot. 2014, 92, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Killeen, D.P.; Larsen, L.; Dayan, F.E.; Gordon, K.C.; Perry, N.B.; van Klink, J.W. Nortriketones: Antimicrobial Trimethylated Acylphloroglucinols from Mānuka (Leptospermum scoparium). J. Nat. Prod. 2016, 79, 564–569. [Google Scholar] [CrossRef]
- van Klink, J.W.; Larsen, L.; Perry, N.B.; Weavers, R.T.; Cook, G.M.; Bremer, P.J.; MacKenzie, A.D.; Kirikae, T. Triketones active against antibiotic-resistant bacteria: Synthesis, structure–activity relationships, and mode of action. Bioorganic Med. Chem. 2005, 13, 6651–6662. [Google Scholar] [CrossRef]
- Mika, A.; Reynolds, S.L.; Mohlin, F.C.; Willis, C.; Swe, P.M.; Pickering, D.A.; Halilovic, V.; Wijeyewickrema, L.C.; Pike, R.N.; Blom, A.M.; et al. Novel Scabies Mite Serpins Inhibit the Three Pathways of the Human Complement System. PLoS ONE 2012, 7, e40489. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, S.L.; Pike, R.N.; Mika, A.; Blom, A.M.; Hofmann, A.; Wijeyewickrema, L.C.; Kemp, D.; Fischer, K. Scabies Mite Inactive Serine Proteases Are Potent Inhibitors of the Human Complement Lectin Pathway. PLoS Neglected Trop. Dis. 2014, 8, e2872. [Google Scholar] [CrossRef] [PubMed]
- Swe, P.M.; Fischer, K. A Scabies Mite Serpin Interferes with Complement-Mediated Neutrophil Functions and Promotes Staphylococcal Growth. PLOS Negl. Trop. Dis. 2014, 8, e2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, L.; Veraldi, S. Skin bacterial colonizations and superinfections in immunocompetent patients with scabies. Int. J. Dermatol. 2018, 57, 1218–1220. [Google Scholar] [CrossRef] [PubMed]
- Swe, P.M.; Reynolds, S.L.; Fischer, K. Parasitic scabies mites and associated bacteria joining forces against host complement defence. Parasite Immunol. 2014, 36, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Sara Taylor, K.F. Understanding the Role of the Scabies Mite Microbiota in the Development of Novel Control Strategies. J. Cell Immunol. 2021, 3, 6. [Google Scholar]
- George, J.H.; Hesse, M.D.; Baldwin, J.E.; Adlington, R.M. Biomimetic Synthesis of Polycyclic Polyprenylated Acylphloroglucinol Natural Products Isolated from Hypericum papuanum. Org. Lett. 2010, 12, 3532–3535. [Google Scholar] [CrossRef]
- van Klink, J.W.; Brophy, J.J.; Perry, N.B.; Weavers, R.T. β-Triketones from Myrtaceae: Isoleptospermone from Leptospermum scoparium and Papuanone from Corymbia dallachiana. J. Nat. Prod. 1999, 62, 487–489. [Google Scholar] [CrossRef]
- Bharate, J.B.; Batarseh, Y.S.; Wani, A.; Sharma, S.; Vishwakarma, R.A.; Kaddoumi, A.; Kumar, A.; Bharate, S.B. Synthesis and P-glycoprotein induction activity of colupulone analogs. Org. Biomol. Chem. 2015, 13, 5488–5496. [Google Scholar] [CrossRef]
- Hellyer, R.O.; Pinhey, J.T. The structure of grandiflorone, a new β-triketone. J. Chem. Soc. C Org. 1966, 19, 1496–1498. [Google Scholar] [CrossRef]
- McDonald, M.; Dougall, A.; Holt, D.; Huygens, F.; Oppedisano, F.; Giffard, P.M.; Inman-Bamber, J.; Stephens, A.J.; Towers, R.; Carapetis, J.R.; et al. Use of a single-nucleotide polymorphism genotyping system to demonstrate the unique epidemiology of methicillin-resistant Staphylococcus aureus in remote aboriginal communities. J. Clin. Microbiol. 2006, 44, 3720–3727. [Google Scholar] [CrossRef] [Green Version]
- Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard—Tenth Edition 2015; Clinical and Laboratory Standards Institute: Malvern, PA, USA, 2015; p. 15.
- Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria; Clinical and Laboratory Standards Institute: Malvern, PA, USA, 2018; p. 10.
- Methods for Determining Bacterial Activity of Antimicrobial Agents; Approved Guidelines; Clinical and Laboratory Standards Institute (CLSI): Malvern, PA, USA, 1999; p. 50.
- Lee, L.Y.L.; Höök, M.; Haviland, D.; Wetsel, R.A.; Yonter, E.O.; Syribeys, P.; Vernachio, J.; Brown, E.L. Inhibition of Complement Activation by a Secreted Staphylococcus aureus Protein. J. Infect. Dis. 2004, 190, 571–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooijakkers, S.H.M.; Milder, F.J.; Bardoel, B.W.; Ruyken, M.; Van Strijp, J.A.G.; Gros, P. Staphylococcal Complement Inhibitor: Structure and Active Sites. J. Immunol. 2007, 179, 2989–2998. [Google Scholar] [CrossRef] [PubMed]
- Laabei, M.; Ermert, D. Catch Me if You Can: Streptococcus pyogenes Complement Evasion Strategies. J. Innate Immun. 2019, 11, 3–12. [Google Scholar] [CrossRef]
- Intorasoot, A.; Chornchoem, P.; Sookkhee, S.; Intorasoot, S. Bactericidal activity of herbal volatile oil extracts against multidrug-resistant Acinetobacter baumannii. J. Intercult. Ethnopharmacol. 2017, 6, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Wicaksono, W.A.; Jones, E.E.; Sansom, C.E.; Perry, N.B.; Monk, J.; Black, A.; Ridgway, H.J. Indigenous bacteria enhance growth and modify essential oil content in Leptospermum scoparium (mānuka). New Zealand J. Bot. 2017, 55, 306–317. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial properties of plant essential oils against human pathogens and their mode of action: An updated review. Evid. Based Complement. Altern. Med. 2016, 2016, 3012462. [Google Scholar] [CrossRef] [Green Version]
- Culp, E.; Wright, G.D. Bacterial proteases, untapped antimicrobial drug targets. J. Antibiot. 2016, 70, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.-W.; Chen, X.-L. Extracellular metalloproteases from bacteria. Appl. Microbiol. Biotechnol. 2011, 92, 253–262. [Google Scholar] [CrossRef]
- Lehman, M.K.; Nuxoll, A.S.; Yamada, K.J.; Kielian, T.; Carson, S.D.; Fey, P.D. Protease-Mediated Growth of Staphylococcus aureus on Host Proteins Is opp3 Dependent. mBio 2019, 10, e02553-18. [Google Scholar] [CrossRef] [Green Version]
- Kolar, S.L.; Ibarra, J.A.; Rivera, F.E.; Mootz, J.M.; Davenport, J.E.; Stevens, S.M.; Horswill, A.R.; Shaw, L.N. Extracellular proteases are key mediators of Staphylococcus aureus virulence via the global modulation of virulence-determinant stability. Microbiologyopen 2013, 2, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Sumitomo, T.; Nakata, M.; Higashino, M.; Terao, Y.; Kawabata, S. Group A Streptococcal Cysteine Protease Cleaves Epithelial Junctions and Contributes to Bacterial Translocation. J. Biol. Chem. 2013, 288, 13317–13324. [Google Scholar] [CrossRef] [PubMed]
- Shelver, D.; Rajagopal, L.; Harris, T.O.; Rubens, C.E. MtaR, a Regulator of Methionine Transport, Is Critical for Survival of Group B Streptococcus In Vivo. J. Bacteriol. 2003, 185, 6592–6599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinsella, R.L.; Lopez, J.; Palmer, L.D.; Salinas, N.; Skaar, E.P.; Tolia, N.; Feldman, M.F. Defining the interaction of the protease CpaA with its type II secretion chaperone CpaB and its contribution to virulence in Acinetobacter species. J. Biol. Chem. 2017, 292, 19628–19638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayasekera, M.M.; Foltin, S.; Olson, E.R.; Holler, T.P. Escherichia coli Requires the Protease Activity of FtsH for Growth. Arch. Biochem. Biophys. 2000, 380, 103–107. [Google Scholar] [CrossRef]
- Bieniossek, C.; Schalch, T.; Bumann, M.; Meister, M.; Meier, R.; Baumann, U. The molecular architecture of the metalloprotease FtsH. Proc. Natl. Acad. Sci. USA 2006, 103, 3066–3071. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, S.; Chaudhry, U.; Raza, A.; Ghosh, D.; Zhao, X. In Vitro activity of ivermectin against Staphylococcus aureus clinical isolates. Antimicrob. Resist. Infect. Control 2018, 7, 27. [Google Scholar] [CrossRef]
- Omansen, T.F.; Porter, J.L.; Johnson, P.D.R.; van der Werf, T.; Stienstra, Y.; Stinear, T.P. In-Vitro Activity of Avermectins against Mycobacterium ulcerans. PLOS Neglected Trop. Dis. 2015, 9, e0003549. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, S.; Parrine, D.; Bilal, M.; Chaudhry, U.; Lefsrud, M.; Zhao, X. Proteomic Comparison of Ivermectin Sensitive and Resistant Staphylococcus aureus Clinical Isolates Reveals Key Efflux Pumps as Possible Resistance Determinants. Antibiotics 2022, 11, 759. [Google Scholar] [CrossRef]
- Romani, L.; Whitfeld, M.J.; Koroivueta, J.; Kama, M.; Wand, H.; Tikoduadua, L.; Tuicakau, M.; Koroi, A.; Andrews, R.; Kaldor, J.M.; et al. Mass Drug Administration for Scabies Control in a Population with Endemic Disease. New Engl. J. Med. 2015, 373, 2305–2313. [Google Scholar] [CrossRef]
- Dinos, G.P. The macrolide antibiotic renaissance. Br. J. Pharmacol. 2017, 174, 2967–2983. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Bacteria (Gram Positive) | Characteristics |
| S. aureus XEN29 | NCTC8532, KanR |
| S. aureus CC75, M34 | Pyoderma clinical isolate, MRSA, lacks staphyloxanthin |
| S. aureus CC75, M5 | Pyoderma clinical isolate, MSSA, lacks staphyloxanthin |
| S. pyogenes 2967 | emm1, M1, PSGN |
| S. pyogenes 2031 | emm1, M1, type strain |
| S. pyogenes 8830 | Pyoderma clinical isolate, emm97 |
| S. dysgalactiae subs. equisimilis MD10 | STG 6 |
| S. dysgalactiae subs. equisimilis NS3396 | Clinical isolate, Acute Rheumatic Fever patient, STG 480 |
| Bacteria (Gram negative) | Characteristics |
| A. baumannii ATCC19606 | Biofilm forming strain, type strain (urine) |
| A. baumannii ATCC17987 | Drug sensitive (fatal meningitis) |
| A. baumannii BAA-1605 | Clinical isolate, sputum, multi-drug resistant |
| A.baumannii ATCC19606 | A.baumannii ATCC17978 | A.baumannii BAA1605 | S. aureus XEN29 NCTC8532 | S. aureus MSSA CC75 M5 | S. aureus MRSA CC75 M34 | S. pyogenes 2031 | S. pyogenes 2967 | S. pyogenes 8830 | S. dysgalactiae MD10 | S. dysgalactiae ns3396 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC (mM) | MBC (mM) | MIC (mM) | MBC (mM) | MIC (mM) | MBC (mM) | MIC (mM) | MBC (mM) | MIC (mM) | MBC (mM) | MIC (mM) | MBC (mM) | MIC (mM) | MBC (mM) | MIC (mM) | MBC (mM) | MIC (mM) | MBC (mM) | MIC (mM) | MBC (mM) | MIC (mM) | MBC (mM) | |
| Flavesone | 12.5 ± 0 | 25 ± 0 | 10 ± 3 | 25 ± 0 | 6.25 ± 0 | 25 ± 0 | 3.125 ± 0 | 12.5 ± 0 | 1.56 ± 0 | 12.5 ± 0 | 1.9 ± 0.7 | 12.5 ± 0 | 1.56 ± 0 | 8.3 ± 3.1 | 1.0 ± 0.4 | 1.91 ± 0.7 | 0.78 ± 0 | 4.7 ± 2.3 | 1.56 ± 0 | 10.4 ± 3.6 | 2.08 ± 0.78 | 12.5 ± 0 |
| Isoleptospermone | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 21 ± 7 | 12.5 ± 0 | 25 ± 0 | 1.56 ± 0 | 3.1 ± 0 | 0.78 ± 0 | 5.2 ± 1.6 | 1.1 ± 0.4 | 6.25 ± 0 | 0.32 ± 0.10 | 10.4 ± 3.1 | 0.78 ± 0 | 1.17 ± 0.58 | 0.39 ± 0 | 3.125 ± 0 | 0.78 ± 0 | 4.17 ± 1.8 | 0.70 ± 0.17 | 8.3 ± 3.6 |
| Leptospermone | 12.5 ± 0 | 21 ± 7 | 12.5 ± 0 | 21 ± 7 | 12.5 ± 0 | 25 ± 0 | 0.78 ± 0 | 1.2 ± 0.4 | 0.35 ± 0.09 | 2.6 ± 0.8 | 0.65 ± 0.19 | 3.1 ± 0 | 0.16 ± 0.05 | 12.5 ± 0 | 0.37 ± 0.06 | 1.17 ± 0.43 | 0.24 ± 0.09 | 6.25 ± 0 | 0.43 ± 0.13 | 6.25 ± 0 | 0.32 ± 0.10 | 6.25 ± 0 |
| Grandiflorone | 12.5 ± 0 | 25 ± 0 | 10 ± 0.3 | 25 ± 0 | 12.5 ± 0 | 12.5 ± 0 | 0.12 ± 0.05 | 0.39 ± 0 | 0.15 ± 0.05 | 1.3 ± 0.4 | 0.16 ± 0.05 | 0.52 ± 0.19 | 0.16 ± 0.04 | 6.25 ± 0 | 0.13 ± 0.04 | 0.195 ± 0 | 0.086 ± 0.060 | 0.780 ± 0 | 0.17 ± 0.04 | 1.17 ± 0.78 | 0.14 ± 0.05 | 1.56 ± 0.0 |
| Mānuka Oil [a] | 3.1 ± 0 | 3.1 ± 0 | 3.1 ± 0 | 3.1 ± 0 | 2.6 ± 0.8 | 3.1 ± 0 | 0.30 ± 0.101 | 0.39 ± 0 | 0.32 ± 0.10 | 0.39 ± 0 | 0.39 ± 0 | 0.39 ± 0 | 0.41 ± 0.16 | 0.91 ± 0.60 | 0.28 ± 0.13 | 0.45 ± 0.30 | 0.20 ± 0.11 | 0.78 ± 0.68 | 0.13 ± 0.05 | 0.52 ± 0.23 | 0.32 ± 0.10 | 0.78 ± 0.68 |
| Abametapir | 1.4 ± 0.3 | - | 0.74 ± 0.13 | - | 1.56 ± 0 | - | 1.56 ± 0 | 1.56 ± 0 | 1.3 ± 0.4 | 5.2 ± 1.6 | 1.3 ± 0.4 | 5.2 ± 1.6 | 3.12 ± 0 | 10.42 ± 3.1 | 3.12 ± 0 | 10.42 ± 3.1 | 2.8 ± 0.5 | 25 ± 0 | 3.12 ± 0 | - | 3.12 ± 0 | - |
| Ivomec (formulation) | 0.21 ± 0.12 | - | 0.23 ± 0.13 | - | 0.39 ± 0.28 | - | 0.71 ± 0 | - | 0.71 ± 0 | - | 0.71 ± 0 | - | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| Ivermectin | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | 0.71 ± 0 | - | 0.15 ± 0.04 | - | 0.71 ± 0 | - | 0.71 ± 0 | - | 0.58 ± 0.21 | - |
| DMSO [a] | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 |
| Isopropanol [a] | 6.25 ± 0 | 12.5 ± 0 | 6.25 ± 0 | 12.5 ± 0 | 6.25 ± 0 | 12.5 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | 12.5 ± 0 | 25 ± 0 | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taylor, S.; Walther, D.; Fernando, D.D.; Swe-Kay, P.; Fischer, K. Investigating the Antibacterial Properties of Prospective Scabicides. Biomedicines 2022, 10, 3287. https://doi.org/10.3390/biomedicines10123287
Taylor S, Walther D, Fernando DD, Swe-Kay P, Fischer K. Investigating the Antibacterial Properties of Prospective Scabicides. Biomedicines. 2022; 10(12):3287. https://doi.org/10.3390/biomedicines10123287
Chicago/Turabian StyleTaylor, Sara, Deonne Walther, Deepani D. Fernando, Pearl Swe-Kay, and Katja Fischer. 2022. "Investigating the Antibacterial Properties of Prospective Scabicides" Biomedicines 10, no. 12: 3287. https://doi.org/10.3390/biomedicines10123287