

A High-Fat and High-Carbohydrate Diet Promotes Reminiscent Hallmarks of an Aging Ovary in the Rabbit Model
 , , and
, , and
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Type and Experimental Design | Main Findings | Reference |
|---|---|---|
| New Zealand rabbits from 10 weeks of age were fed high-fat, high-cholesterol (HH) diet | They analyzed follicular populations in the ovary. A higher number of atretic follicles and a reduced number of antral follicles were observed in HH group. | [23] |
| Ossabaw mini pigs were fed a high fat/cholesterol/fructose for eight months | Obese pigs showed more cystic, medium, and ovulatory-size follicles. | [24] |
| Sprague Dawley rats were fed a high-fat diet (HFD) for 18 weeks | Depletion of the ovarian reserve was analyzed. They observed a decrease in primordial follicles whereas the number of growing-to-primordial follicles was higher. | [16] |
| C57BL/6J mice strain were fed an HFD for 10 weeks | Depletion of the ovarian reserve was analyzed. They noticed decreased primordial follicles. | [25] |
| C57BL/6J mice strain were fed a HFD for 10 weeks | They analyzed depletion of the ovarian reserve. Decreased primordial follicles. | [26] |
| Mice from the Swiss albino strain were fed an HFD for 9 weeks | They evaluated the number and developmental morphology of follicles.They observed a reduced number of primordial, while the number of primary, secondary, and tertiary follicles increased with HFD. | [27] |
| Wistar rats were fed a high carbohydrate diet (HCD) for 15 days | A reduced number of primordial, primary, preantral, and antral follicles were observed in the HCD group. | [28] |
| Wistar rats were fed a high-sucrose diet (HSD) for 4 months | The HSD group showed a greater number of atretic antral follicles and cystic follicles. However, the number of follicles between groups did not manifest differences. | [29] |
2. Materials and Methods
2.1. Masson’s Trichrome Stain and Ovarian Follicle Counting
2.2. Lipofuscin Detection and Analysis
2.3. Immunohistochemistry and Immunofluorescence
2.4. Statistical Analysis
3. Results
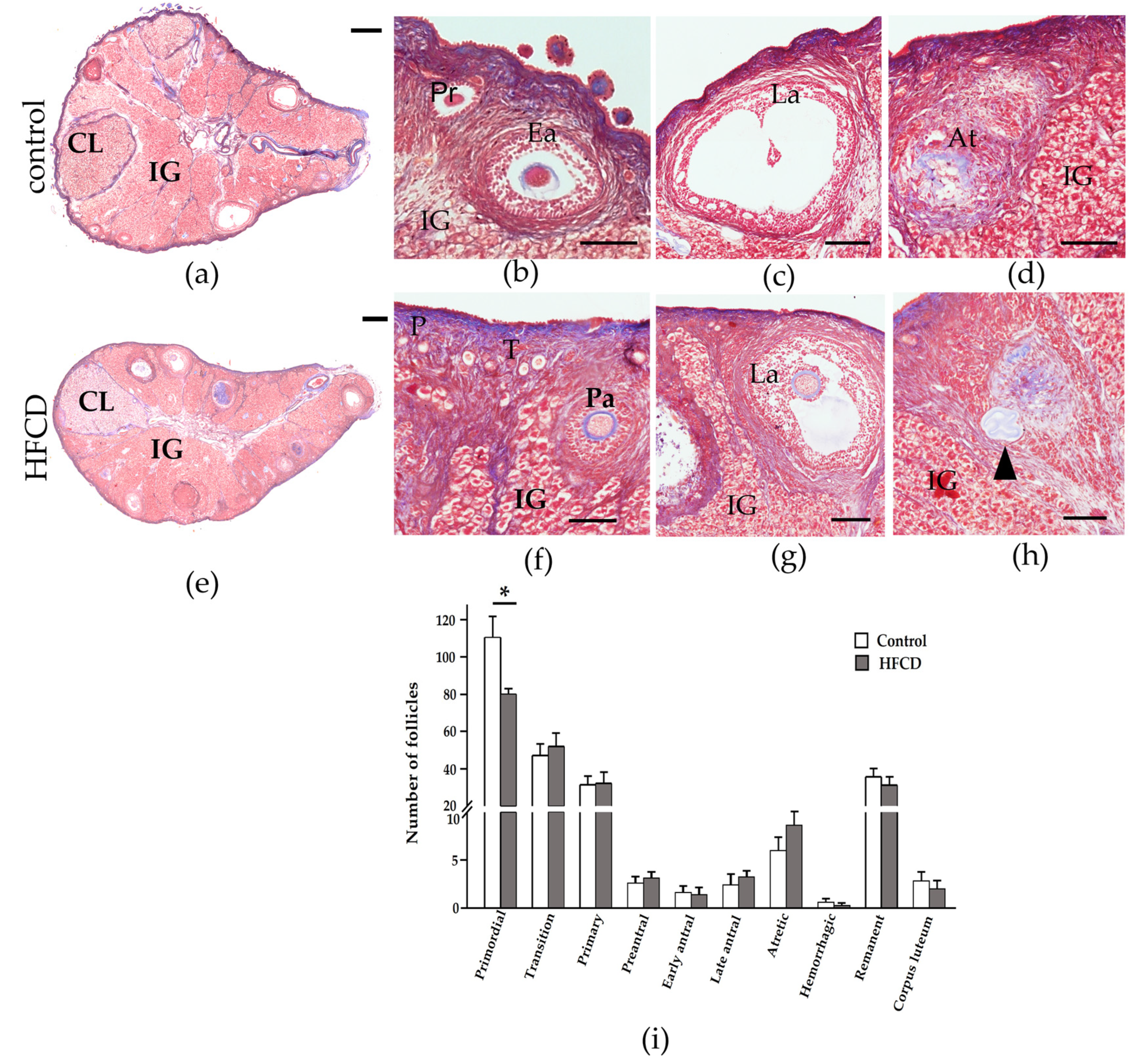
3.1. HFCD Caused a Decreased in the Ovary’s Follicular Reserve
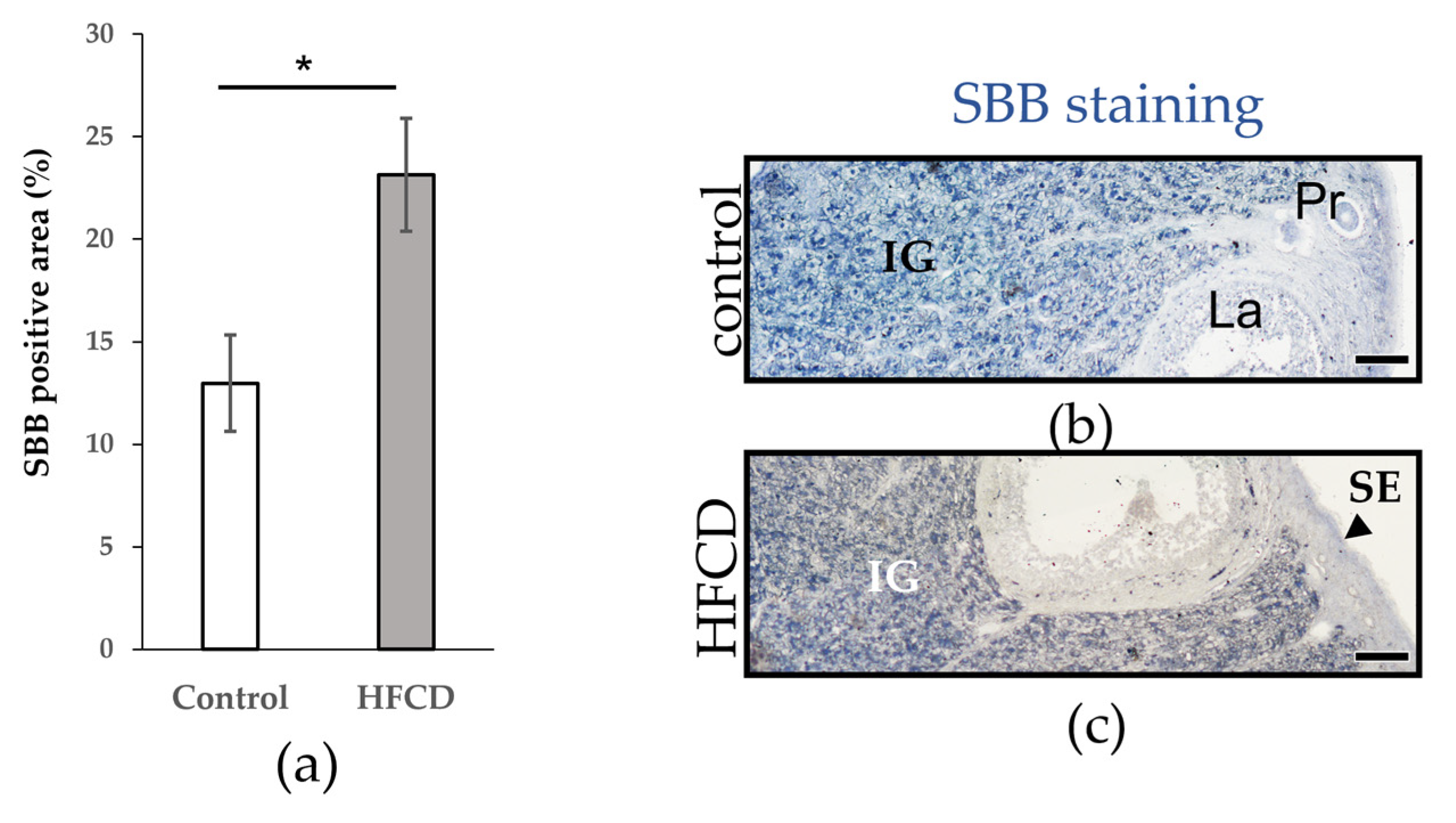
3.2. HFCD Increased the Presence of Lipofuscin in the Interstitial Gland
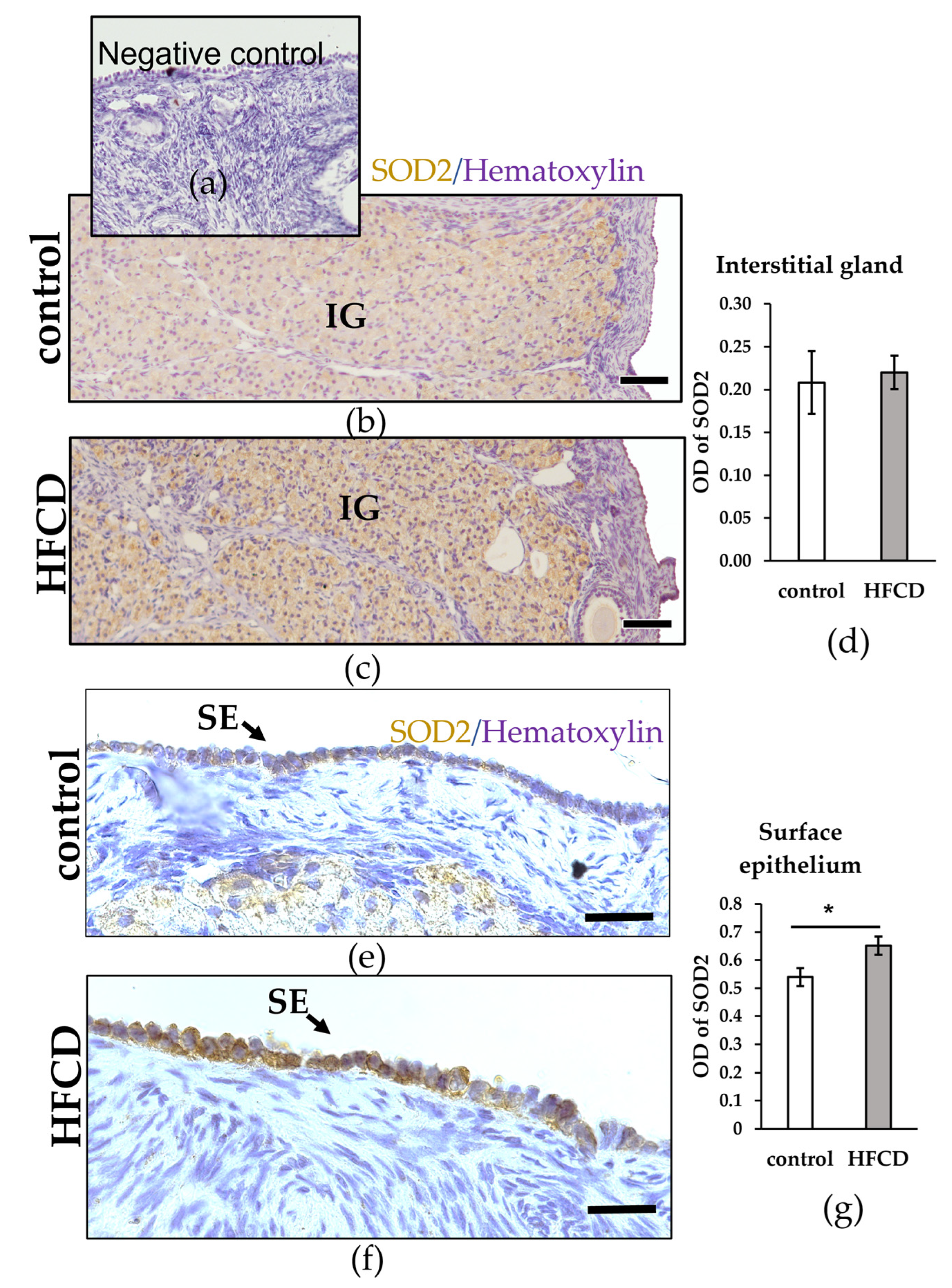
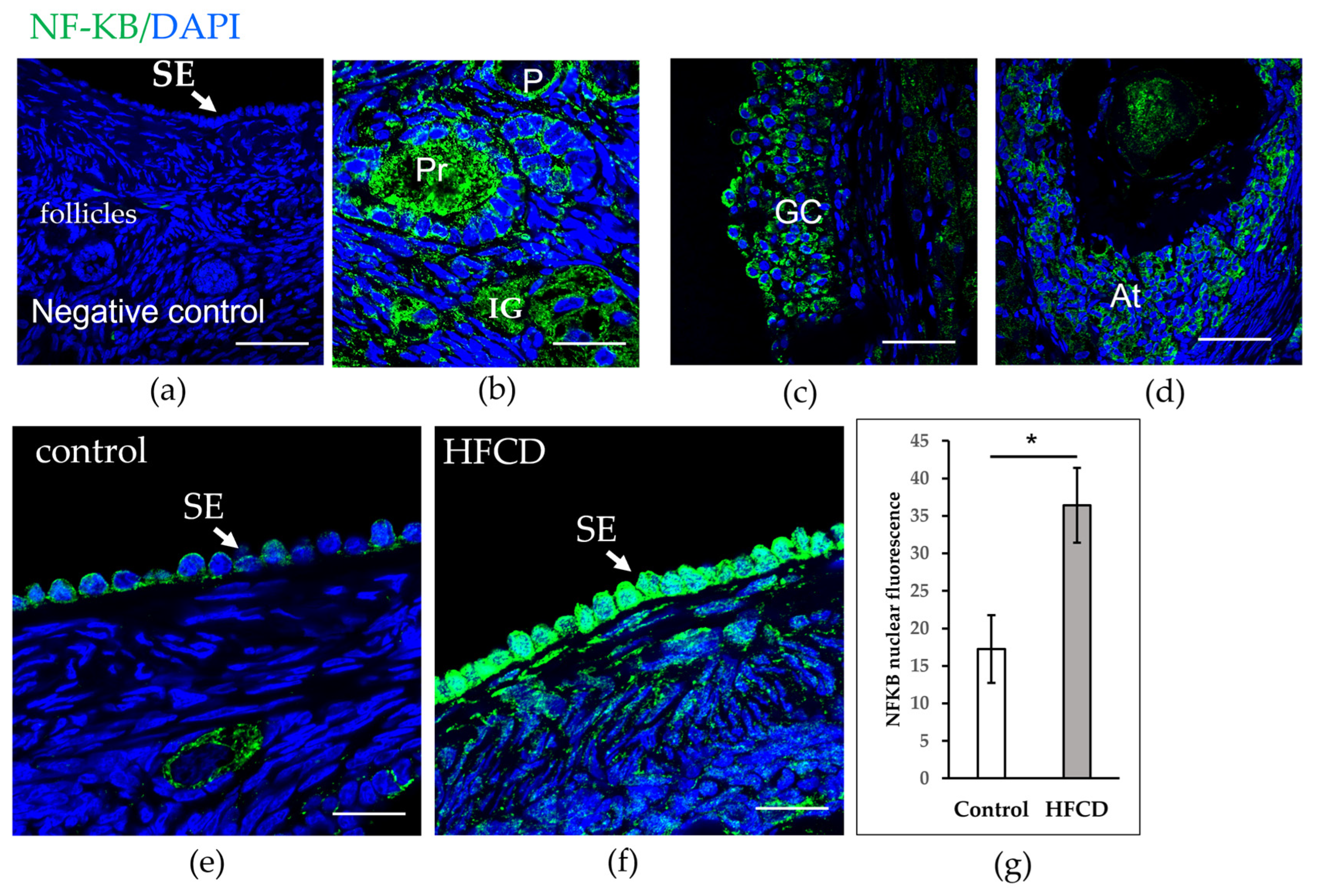
3.3. HFCD Induces NF-ΚΒ p65 Nuclear Translocation in Cells of Surface Epithelium
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ansere, V.A.; Ali-Mondal, S.; Sathiaseelan, R.; Garcia, D.N.; Isola, J.V.V.; Henseb, J.D.; Saccon, T.D.; Ocañas, S.R.; Tooley, K.B.; Stout, M.B.; et al. Cellular hallmarks of aging emerge in the ovary prior to primordial follicle depletion. Mech. Ageing Dev. 2021, 194, 111425. [Google Scholar] [CrossRef]
- Broekmans, F.J.; Knauff, E.A.; te Velde, E.R.; Macklon, N.S.; Fauser, B.C. Female reproductive ageing: Current knowledge and future trends. Trends Endocrinol. Metab. 2007, 18, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Tatone, C.; Amicarelli, F. The aging ovary--the poor granulosa cells. Fertil. Steril. 2013, 99, 12–17. [Google Scholar] [CrossRef]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef] [PubMed]
- Ilie, O.D.; Ciobica, A.; Riga, S.; Dhunna, N.; McKenna, J.; Mavroudis, I.; Doroftei, B.; Ciobanu, A.M.; Riga, D. Mini-Review on Lipofuscin and Aging: Focusing on The Molecular Interface, The Biological Recycling Mechanism, Oxidative Stress, and The Gut-Brain Axis Functionality. Medicina 2020, 56, 626. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Luderer, U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biol. Reprod. 2011, 84, 775–782. [Google Scholar] [CrossRef] [Green Version]
- Höhn, A.; Grune, T. Lipofuscin: Formation, effects and role of macroautophagy. Redox Biol. 2013, 1, 140–144. [Google Scholar] [CrossRef] [Green Version]
- Helenius, M.; Hänninen, M.; Lehtinen, S.K.; Salminen, A. Changes associated with aging and replicative senescence in the regulation of transcription factor nuclear factor-kappa B. Biochem. J. 1996, 318, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.S.; Aggarwal, B.B. Transcription Factor NF-κB: A Sensor for Smoke and Stress Signals. Ann. N. Y. Acad. Sci. 2005, 1056, 218–233. [Google Scholar] [CrossRef] [PubMed]
- Irshad, M.; Chaudhuri, P.S. Oxidant-antioxidant system: Role and significance in human body. Indian J. Exp. Biol. 2002, 40, 1233–1239. [Google Scholar] [PubMed]
- White, K.L.; Rider, D.N.; Kalli, K.R.; Knutson, K.L.; Jarvik, G.P.; Goode, E.L. Genomics of the NF-κB signaling pathway: Hypothesized role in ovarian cancer. Cancer Causes Control. 2011, 22, 785–801. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Liu, L.; Li, C.; Luo, N.; Chen, R.; Li, L.; Yu, F.; Cheng, Z. TRIM52 plays an oncogenic role in ovarian cancer associated with NF-kB pathway. Cell Death Dis. 2018, 9, 908. [Google Scholar] [CrossRef] [Green Version]
- Silvestris, E.; Lovero, D.; Palmirotta, R. Nutrition and Female Fertility: An Interdependent Correlation. Front. Endocrinol. 2019, 10, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogrodnik, M.; Zhu, Y.; Langhi, L.G.P.; Tchkonia, T.; Krüger, P.; Fielder, E.; Victorelli, S.; Ruswhandi, R.A.; Giorgadze, N.; Pirtskhalava, T.; et al. Obesity-Induced Cellular Senescence Drives Anxiety and Impairs Neurogenesis. Cell Metab. 2019, 29, 1061–1077.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snider, A.P.; Wood, J.R. Obesity induces ovarian inflammation and reduces oocyte quality. Reproduction 2019, 158, R79–R90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Luo, L.L.; Xu, J.J.; Xu, M.Y.; Zhang, X.M.; Zhou, X.L.; Liu, W.J.; Fu, Y.C. Obesity accelerates ovarian follicle development and follicle loss in rats. Metab. Clin. Exp. 2014, 63, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Chinkes, D.L.; Aarsland, A.; Herndon, D.N.; Wolfe, R.R. Lipid metabolism in diet-induced obese rabbits is similar to that of obese humans. J. Nutr. 2008, 138, 515–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Zhang, C.; Yang, W.; Wang, Y.; Lin, Y.; Yang, P.; Yu, Q.; Fan, J.; Liu, E. Fat and cholesterol diet induced lipid metabolic disorders and insulin resistance in rabbit. Exp. Clin. Endocrinol. Diabetes 2009, 117, 400–405. [Google Scholar] [CrossRef]
- Zhao, S.; Chu, Y.; Zhang, C.; Lin, Y.; Xu, K.; Yang, P.; Fan, J.; Liu, E. Diet-induced central obesity and insulin resistance in rabbits. J. Anim. Physiol. Anim. Nutr. 2008, 92, 105–111. [Google Scholar] [CrossRef]
- Díaz-Hernández, V.; Caldelas, I.; Merchant-Larios, H. Gene Expression in the Supporting Cells at the Onset of Meiosis in Rabbit Gonads. Sex. Dev. 2019, 13, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Banco, B.; Grilli, G.; Giudice, C.; Marques, A.T.; Cotti Cometti, S.; Visigalli, G.; Grieco, V. Immunophenotyping of Rabbit Testicular Germ and Sertoli Cells Across Maturational Stages. J. Histochem. Cytochem. 2016, 64, 715–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Hernández, V.; Caldelas, I.; Montaño, L.M.; Merchant-Larios, H. Morphological rearrangement of the cortical region, in aging ovaries. Histol. Histopathol. 2019, 34, 775–789. [Google Scholar] [PubMed]
- Cordier, A.G.; Léveillé, P.; Dupont, C.; Tarrade, A.; Picone, O.; Larcher, T.; Dahirel, M.; Poumerol, E.; Mandon-Pepin, B.; Lévy, R.; et al. Dietary lipid and cholesterol induce ovarian dysfunction and abnormal LH response to stimulation in rabbits. PLoS ONE 2013, 8, e63101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newell-Fugate, A.E.; Taibl, J.N.; Alloosh, M.; Sturek, M.; Bahr, J.M.; Nowak, R.A.; Krisher, R.L. Effects of Obesity and Metabolic Syndrome on Steroidogenesis and Folliculogenesis in the Female Ossabaw Mini-Pig. PLoS ONE 2015, 10, e0128749. [Google Scholar] [CrossRef] [Green Version]
- Skaznik-Wikiel, M.E.; Swindle, D.C.; Allshouse, A.A.; Polotsky, A.J.; McManaman, J.L. High-Fat Diet Causes Subfertility and Compromised Ovarian Function Independent of Obesity in Mice. Biol. Reprod. 2016, 94, 108. [Google Scholar] [CrossRef]
- Hohos, N.M.; Cho, K.J.; Swindle, D.C.; Skaznik-Wikiel, M.E. High-fat diet exposure, regardless of induction of obesity, is associated with altered expression of genes critical to normal ovulatory function. Mol. Cell. Endocrinol. 2018, 470, 199–207. [Google Scholar] [CrossRef]
- Hilal, G.; Fatma, T.; Ferruh, Y.; Sabire, G.; Yüksel, A. Effect of high-fat diet on the various morphological parameters of the ovary. Anat. Cell Biol. 2020, 53, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Niño, O.M.S.; da Costa, C.S.; Torres, K.M.; Zanol, J.F.; Freitas-Lima, L.C.; Miranda-Alves, L.; Graceli, J.B. High-refined carbohydrate diet leads to polycystic ovary syndrome-like features and reduced ovarian reserve in female rats. Toxicol. Lett. 2020, 332, 42–55. [Google Scholar] [CrossRef]
- de Melo, G.B.; Soares, J.F.; Costa, T.C.L.; Benevides, R.O.A.; Vale, C.C.; Paes, A.M.A.; Gaspar, R.S. Early Exposure to High-Sucrose Diet Leads to Deteriorated Ovarian Health. Front. Endocrinol. 2021, 12, 656831. [Google Scholar] [CrossRef]
- Navarrete, E.; Díaz, G.; Salazar, A.M.; Montúfar-Chaveznava, R.; Caldelas, I. Long-term changes in the diurnal temporal regulation and set points of metabolic parameters associated with chronic maternal overnutrition in rabbits. Am. J. Physiol. Endocrinol. Metab. 2022, in press. [Google Scholar] [CrossRef]
- Navarrete, E.; Díaz-Villaseñor, A.; Díaz, G.; Salazar, A.M.; Montúfar-Chaveznava, R.; Ostrosky-Wegman, P.; Caldelas, I. Misadjustment of diurnal expression of core temperature and locomotor activity in lactating rabbits associated with maternal over-nutrition before and during pregnancy. PLoS ONE 2020, 15, e0232400. [Google Scholar] [CrossRef] [PubMed]
- Salazar, A.M.; Sordo, M.; Navarrete-Monroy, E.; Pánico, P.; Díaz-Villaseñor, A.; Montúfar-Chaveznava, R.; Caldelas, I.; Ostrosky-Wegman, P. Maternal overnutrition before and during pregnancy induces DNA damage in male offspring: A rabbit model. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2021, 865, 503324. [Google Scholar] [CrossRef] [PubMed]
- Exbrayat, J.M. Microscopy: Light Microscopy and Histochemical Methods. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; pp. 715–723. [Google Scholar]
- Rodríguez-Castelán, J.; Méndez-Tepepa, M.; Rodríguez-Antolín, J.; Castelán, F.; Cuevas-Romero, E. Hypothyroidism affects lipid and glycogen content and peroxisome proliferator-activated receptor δ expression in the ovary of the rabbit. Reprod. Fertil. Dev. 2018, 30, 1380–1387. [Google Scholar] [CrossRef] [PubMed]
- Hense, J.D.; Garcia, D.N.; Isola, J.V.; Alvarado-Rincón, J.A.; Zanini, B.M.; Prosczek, J.B.; Stout, M.B.; Mason, J.B.; Walsh, P.T.; Brieño-Enríquez, M.A.; et al. Senolytic treatment reverses obesity-mediated senescent cell accumulation in the ovary. GeroScience 2022, 44, 1747–1759. [Google Scholar] [CrossRef]
- Urzua, U.; Chacon, C.; Espinoza, R.; Martínez, S.; Hernandez, N. Parity-Dependent Hemosiderin and Lipofuscin Accumulation in the Reproductively Aged Mouse Ovary. Anal. Cell. Pathol. 2018, 2018, 1289103. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Pirtskhalava, T.; Farr, J.N.; Weigand, B.M.; Palmer, A.K.; Weivoda, M.M.; Inman, C.L.; Ogrodnik, M.B.; Hachfeld, C.M.; Fraser, D.G.; et al. Senolytics improve physical function and increase lifespan in old age. Nat. Med. 2018, 24, 1246–1256. [Google Scholar] [CrossRef]
- Limon, M. Étude histologique et histogénique de la glande interstitielle de l’ovaire. Arch. D’ant. Micro 1902, 5, 155–190. [Google Scholar]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Auersperg, N.; Wong, A.S.; Choi, K.C.; Kang, S.K.; Leung, P.C. Ovarian surface epithelium: Biology, endocrinology, and pathology. Endocr. Rev. 2001, 22, 255–288. [Google Scholar]
- Agarwal, A.; Gupta, S.; Sharma, R.K. Role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2005, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Murdoch, W.J.; Martinchick, J.F. Oxidative damage to DNA of ovarian surface epithelial cells affected by ovulation: Carcinogenic implication and chemoprevention. Exp. Biol. Med. 2004, 229, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Terman, A. Catabolic insufficiency and aging. Ann. N. Y. Acad. Sci. 2006, 1067, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Li, Y.; Ma, Z.; Jing, J.; Zhang, Z.; Liu, Y.; Ding, Z. Obesity induces morphological and functional changes in female reproductive system through increases in NF-κB and MAPK signaling in mice. Reprod. Biol. Endocrinol. 2021, 19, 148. [Google Scholar] [CrossRef]




| Composition | Control Diet (%) | HFCD (%) |
|---|---|---|
| Digestive carbohydrates (Nitrogen-free extract) | 47.8 | 52.6 |
| Fat (ether extract) | 3.8 | 5.6 |
| Crude protein | 15.7 | 11.2 |
| Crude fiber | 15.3 | 10.2 |
| Mineral (ashes) | 11.5 | 7.6 |
| Humidity | 5.6 | 12.8 |
| Primary Antibodies | Host | Dilution | Cat. Number | Supplier |
|---|---|---|---|---|
| Superoxide dismutase 2 (SOD 2) | Mouse | 1:50 | GTX630558 | GeneTex |
| NF-ΚΒ p65 (G-8) | Mouse | 1:60 | sc-398442 | Santa Cruz Biotechnology |
| Secondary antibodies | ||||
| Biotinylated anti-Mouse IgG | --- | 1:500 | BMK-2202 | Vector Laboratories |
| Alexa Fluor 488 anti-mouse IgG | Donkey | 1:500 | A21202 | Thermo Fisher Scientific |
| HRP-conjugated streptavidin | --- | 1:500 | SA-5004 | Vector Laboratories |
| Control | HFCD | |
|---|---|---|
| Stroma | + | + |
| Surface epithelium | + | + |
| Primordial follicles | + | + |
| Primary follicles | + | + |
| Secondary follicles (granulosa cells) | ++ | ++ |
| Late secondary follicles (granulosa cells) | ++ | ++ |
| Oocytes | ++ | ++ |
| Cumulus cells | +++ | +++ |
| Interstitial gland | ++ | +++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Hernández, V.; Montaño, L.M.; Caldelas, I.; Marmolejo-Valencia, A. A High-Fat and High-Carbohydrate Diet Promotes Reminiscent Hallmarks of an Aging Ovary in the Rabbit Model. Biomedicines 2022, 10, 3068. https://doi.org/10.3390/biomedicines10123068
Díaz-Hernández V, Montaño LM, Caldelas I, Marmolejo-Valencia A. A High-Fat and High-Carbohydrate Diet Promotes Reminiscent Hallmarks of an Aging Ovary in the Rabbit Model. Biomedicines. 2022; 10(12):3068. https://doi.org/10.3390/biomedicines10123068
Chicago/Turabian StyleDíaz-Hernández, Verónica, Luis M. Montaño, Ivette Caldelas, and Alejandro Marmolejo-Valencia. 2022. "A High-Fat and High-Carbohydrate Diet Promotes Reminiscent Hallmarks of an Aging Ovary in the Rabbit Model" Biomedicines 10, no. 12: 3068. https://doi.org/10.3390/biomedicines10123068