
Microbiome Shapes the T Cell Receptor Repertoire among CD4+CD8+ Thymocytes

 and
and 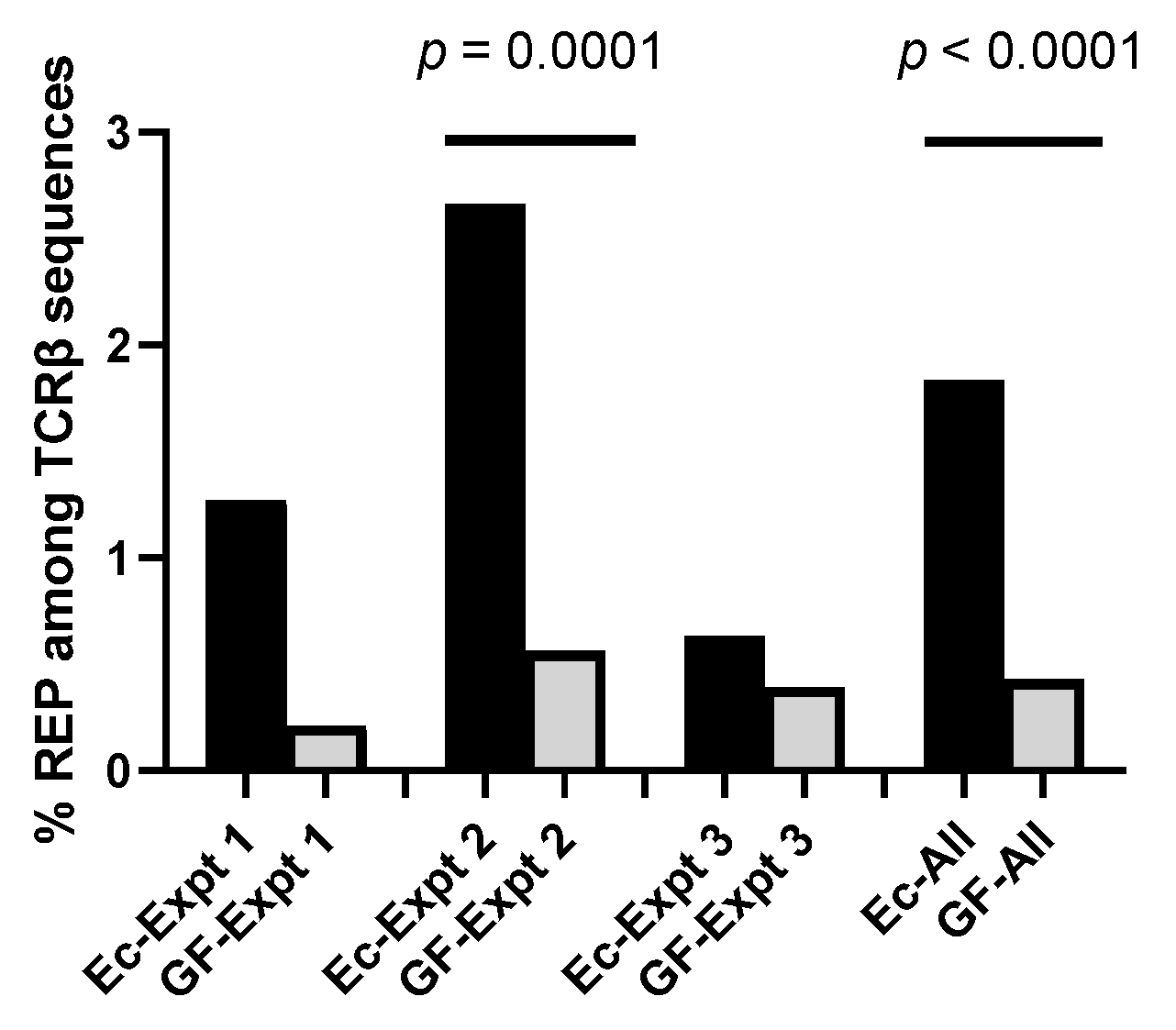{kind=link}
{kind=link}
{kind=link}
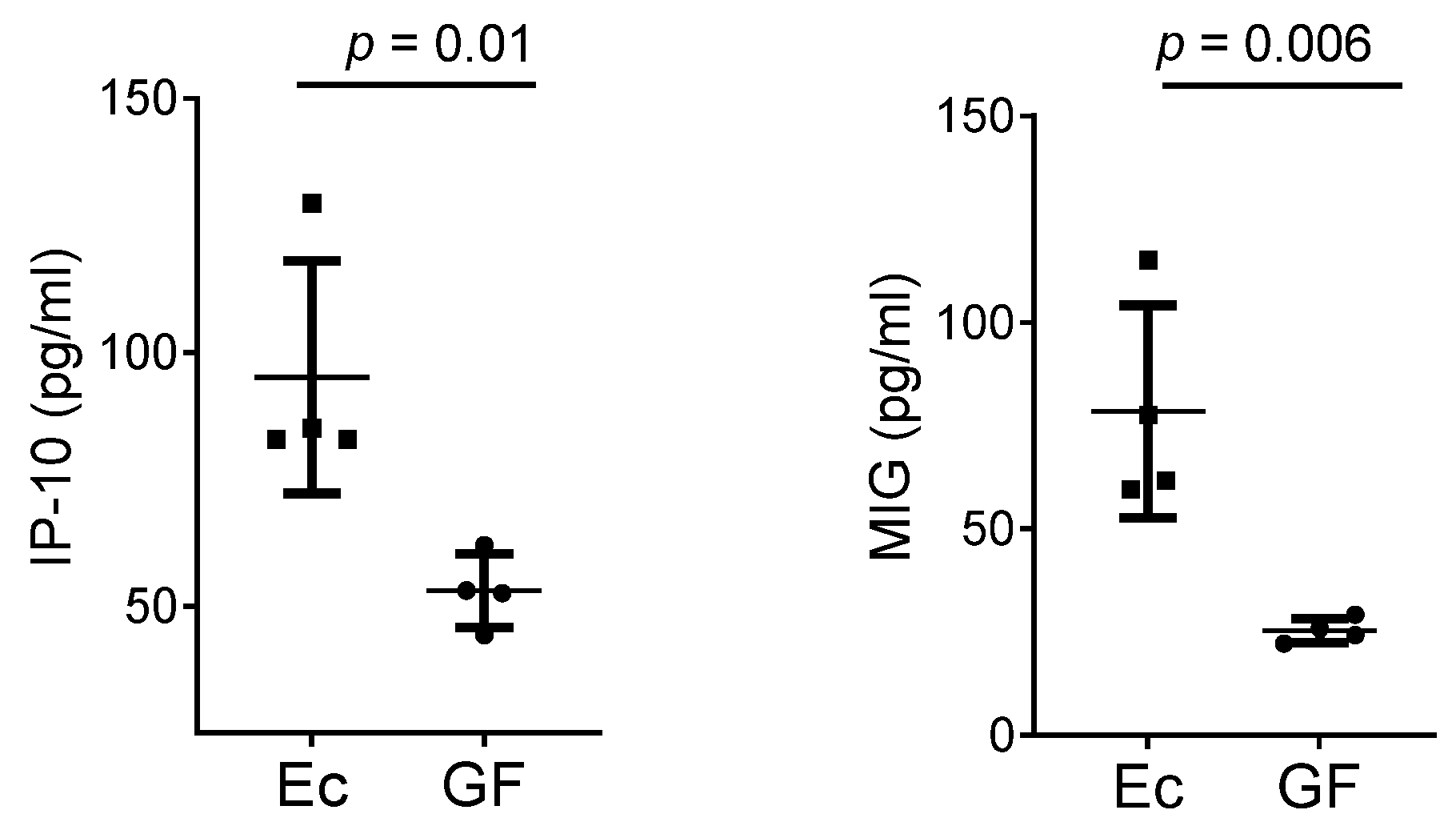{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Single-Cell Sorting
2.3. Single-Cell Sequencing of TCR
2.4. Cytokine Analyses
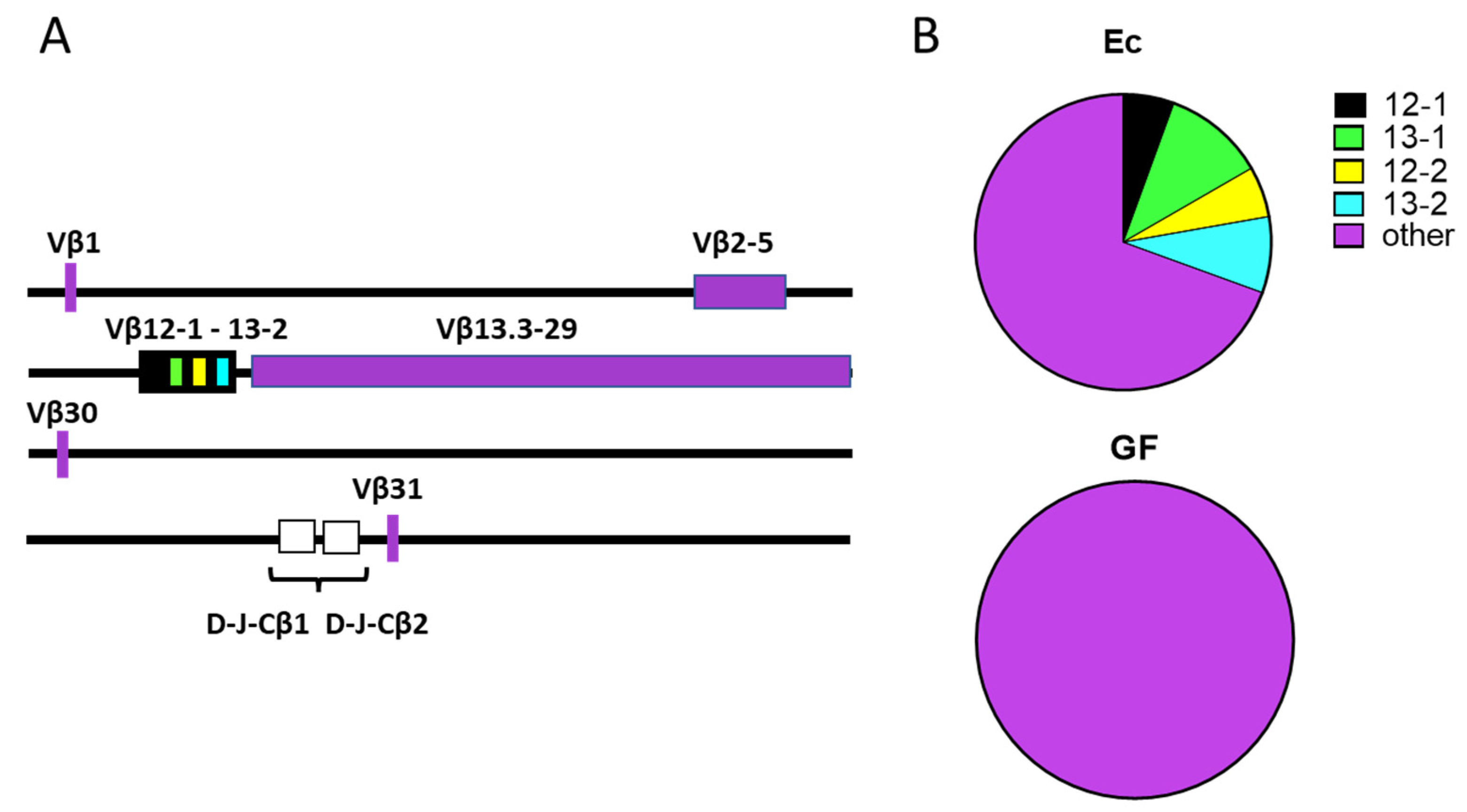
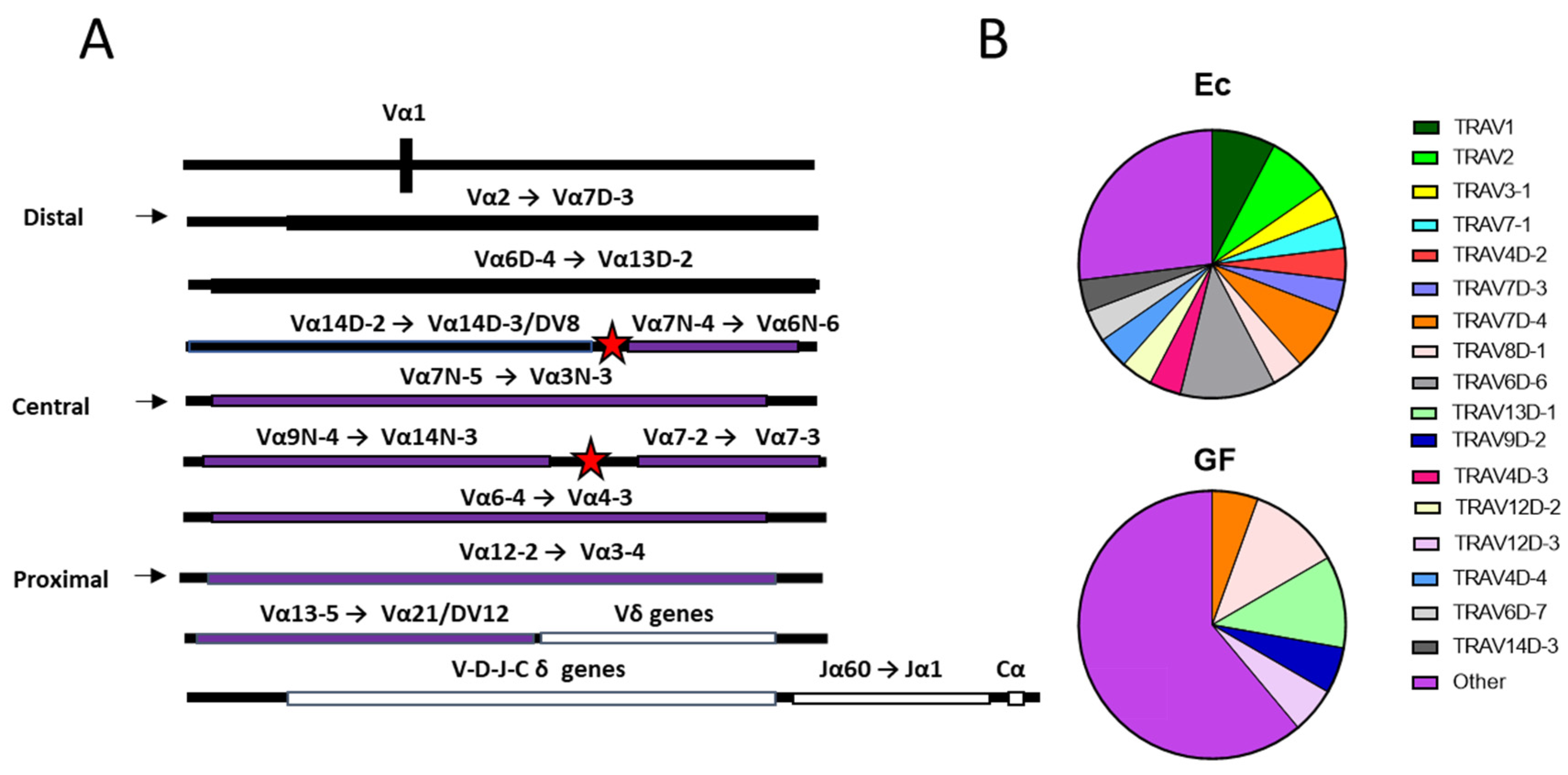
3. Results
4. Discussion
4.1. The Microbiome Influences TCR Repertoires among Developing CD4+CD8+ Thymocytes
4.2. Mechanisms Pertinent to TCR Repertoire Manipulation
4.3. Cytokines and Chemokines
4.4. Clinical Implications of a TCR Bias
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TCR | T cell receptor |
| V | Variable |
| D | Diversity |
| J | Joining |
| C | Constant |
| GF | germ-free |
| Ec | E. coli mono-associated |
| REP | repeat |
References
- Born, W.; Yagüe, J.; Palmer, E.; Kappler, J.; Marrack, P. Rearrangement of T-cell receptor beta-chain genes during T-cell development. Proc. Natl. Acad. Sci. USA 1985, 82, 2925–2929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, S.D.; Pelkonen, J.; Hurwitz, J.L. First T cell receptor alpha gene rearrangements during T cell ontogeny skew to the 5’ region of the J alpha locus. J. Immunol. 1990, 145, 2347–2352. [Google Scholar] [PubMed]
- The international ImMunoGeneTics Information System. Available online: www.imgt.org. (accessed on 11 February 2022).
- Wilson, A.; Marechal, C.; MacDonald, H.R. Biased V beta usage in immature thymocytes is independent of DJ beta proximity and pT alpha pairing. J. Immunol. 2001, 166, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Olson, D.; Shoukry, N.H.; Brady, K.W.; Kim, H.; Olson, D.P.; Hartman, K.; Shintani, A.K.; Walker, C.M.; Kalams, S.A. Limited T Cell Receptor Diversity of HCV-specific T Cell Responses Is Associated with CTL Escape. J. Exp. Med. 2004, 200, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Su, J.; Zhang, X.-B.; Yang, Z.; Zhou, M.-Q.; Jiang, Z.-M.; Hao, P.-P.; Liu, S.-D.; Wen, Q.; Jin, Q.; et al. Limited T Cell Receptor Repertoire Diversity in Tuberculosis Patients Correlates with Clinical Severity. PLoS ONE 2012, 7, e48117. [Google Scholar] [CrossRef] [Green Version]
- Porritt, R.A.; Paschold, L.; Rivas, M.N.; Cheng, M.H.; Yonker, L.M.; Chandnani, H.; Lopez, M.; Siminica, D.; Schultheiß, C.; Santiskulvong, C.; et al. Identification of a unique TCR repertoire, consistent with a superantigen selection process in Children with Multi-system Inflammatory Syndrome. bioRxiv 2020, 372169. [Google Scholar]
- Lu, C.; Pi, X.; Xu, W.; Qing, P.; Tang, H.; Li, Y.; Zhao, Y.; Liu, X.; Tang, H.; Liu, Y. Clinical significance of T cell receptor repertoire in primary Sjogren’s syndrome. EBioMedicine 2022, 84, 104252. [Google Scholar] [CrossRef]
- Rodríguez-Caparrós, A.; Álvarez-Santiago, J.; Del Valle-Pastor, M.J.; Suñé, C.; López-Ros, J.; Hernández-Munain, C. Regulation of T-cell Receptor Gene Expression by Three-Dimensional Locus Conformation and Enhancer Function. Int. J. Mol. Sci. 2020, 21, 8478. [Google Scholar] [CrossRef]
- Thomas-Claudepierre, A.-S.; Robert, I.; Rocha, P.P.; Raviram, R.; Schiavo, E.; Heyer, V.; Bonneau, R.; Luo, V.M.; Reddy, J.K.; Borggrefe, T.; et al. Mediator facilitates transcriptional activation and dynamic long-range contacts at the IgH locus during class switch recombination. J. Exp. Med. 2016, 213, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.G.; Sealy, R.E.; Penkert, R.R.; Surman, S.L.; Maul, R.W.; Neale, G.; Xu, B.; Gearhart, P.J.; Hurwitz, J.L. Complex sex-biased antibody responses: Estrogen receptors bind estrogen response elements centered within immunoglobulin heavy chain gene enhancers. Int. Immunol. 2019, 31, 141–156. [Google Scholar] [CrossRef]
- Hosokawa, H.; Rothenberg, E.V. Cytokines, Transcription Factors, and the Initiation of T-Cell Development. Cold Spring Harb. Perspect. Biol. 2018, 10, a028621. [Google Scholar] [CrossRef]
- Larche, M.; Rencher, S.D.; Hurwitz, J.L. Environmental influence on T cell receptor alpha gene rearrangement and expression in vitro. Eur. J. Immunol. 1992, 22, 2733–2736. [Google Scholar] [CrossRef] [PubMed]
- Zegarra-Ruiz, D.F.; Kim, D.V.; Norwood, K.; Kim, M.; Wu, W.-J.H.; Saldana-Morales, F.B.; Hill, A.A.; Majumdar, S.; Orozco, S.; Bell, R.; et al. Thymic development of gut-microbiota-specific T cells. Nature 2021, 594, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Malchiodi, E.; Eisenstein, E.; Fields, A.B.; Ohlendorf, D.H.; Schlievert, P.; Karjalainen, K.; Mariuzza, A.R. Superantigen binding to a T cell receptor beta chain of known three-dimensional structure. J. Exp. Med. 1995, 182, 1833–1845. [Google Scholar] [CrossRef] [Green Version]
- Schirmer, M.; Smeekens, S.P.; Vlamakis, H.; Jaeger, M.; Oosting, M.; Franzosa, E.A.; Horst, T.B.; Jansen, T.; Jacobs, L.; Bonder, M.C.; et al. Linking the Human Gut Microbiome to Inflammatory Cytokine Production Capacity. Cell 2016, 167, 1125e8–1136e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, K.; Zhu, L.; Bin Li, B.; Zong, D.; Cai, P.; Jiang, C.; Du, P.; Lin, J.; Qu, K. Development of double-positive thymocytes at single-cell resolution. Genome Med. 2021, 13, 49. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Tonkonogy, S.L.; Albright, C.A.; Tsang, J.; Balish, E.J.; Braun, J.; Huycke, M.M.; Sartor, R.B. Variable phenotypes of enterocolitis in interleukin 10-deficient mice monoassociated with two different commensal bacteria. Gastroenterology 2005, 128, 891–906. [Google Scholar] [CrossRef] [PubMed]
- Dash, P.; Fiore-Gartland, A.J.; Hertz, T.; Wang, G.C.; Sharma, S.; Souquette, A.; Crawford, J.C.; Clemens, E.B.; Nguyen, T.H.O.; Kedzierska, K.; et al. Quantifiable predictive features define epitope-specific T cell receptor repertoires. Nature 2017, 547, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Dash, P.; McClaren, J.L.; Oguin, T.H.; Rothwell, W.; Todd, B.; Morris, M.Y.; Becksfort, J.; Reynolds, C.; Brown, S.A.; Doherty, P.C.; et al. Paired analysis of TCRalpha and TCRbeta chains at the single-cell level in mice. J. Clin. Investig. 2011, 121, 288–295. [Google Scholar] [CrossRef]
- Larche, M.; Manzo, A.R.; Hurwitz, J.L. Environmental and allele-specific influences on T cell receptor gene rearrangement: Skewed alpha, delta, and gamma gene rearrangement patterns in chimeric mice. Eur. J. Immunol. 1991, 21, 2943–2949. [Google Scholar] [CrossRef]
- Amedei, A.; Barcelo-Coblijn, G. Editorial of Special Issue “The Interplay of Microbiome and Immune Response in Health and Diseases”. Int. J. Mol. Sci. 2019, 20, 3708. [Google Scholar] [CrossRef]
- Deacy, A.M.; Gan, S.K.-E.; Derrick, J.P. Superantigen Recognition and Interactions: Functions, Mechanisms and Applications. Front. Immunol. 2021, 12, 731845. [Google Scholar] [CrossRef]
- Oh, P.H.Y.; Visvalingam, V.; Wahli, W. The PPAR-microbiota-metabolic organ trilogy to fine-tune physiology. FASEB J. 2019, 33, 9706–9730. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, A.; Negishi, N.; Tsurui, H.; Kadowaki-Ohtsuji, N.; Maeda, K.; Nanno, M.; Yamaguchi, Y.; Shimizu, N.; Yagita, H.; Okumura, K.; et al. Commensal Bacteria Regulate Thymic Aire Expression. PLoS ONE 2014, 9, e105904. [Google Scholar] [CrossRef]
- Ennamorati, M.; Vasudevan, C.; Clerkin, K.; Halvorsen, S.; Verma, S.; Ibrahim, S.; Prosper, S.; Porter, C.; Yeliseyev, V.; Kim, M.; et al. Intestinal microbes influence development of thymic lymphocytes in early life. Proc. Natl. Acad. Sci. USA 2020, 117, 2570–2578. [Google Scholar] [CrossRef]
- Murphy, M.K.; Heimberger, A.B.; Loh, D.Y. Induction by antigen of intrathymic apoptosis of CD4+CD8+TCRlo thymocytes in vivo. Science 1990, 250, 1720–1723. [Google Scholar] [CrossRef] [PubMed]
- Lazzeri, E.; Romagnani, P. CXCR3-binding chemokines: Novel multifunctional therapeutic targets. Curr. Drug Targets Immune Endocr. Metab. Disord 2005, 5, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, P.; Annunziato, F.; Lazzeri, E.; Cosmi, L.; Beltrame, C.; Lasagni, L.; Gali, G.; Francalanci, M.; Manetti, R.; Marra, F. Interferon-inducible protein 10, monokine induced by interferon gamma, and interferon-inducible T-cell alpha chemoattractant are produced by thymic epithelial cells and attract T-cell receptor (TCR) alphabeta+ CD8+ single-positive T cells, TCRgammadelta+ T cells, and natural killer-type cells in human thymus. Blood 2001, 97, 601–607. [Google Scholar] [PubMed] [Green Version]
- Brosnahan, A.J.; Schlievert, P. Gram-positive bacterial superantigen outside-in signaling causes toxic shock syndrome. FEBS J. 2011, 278, 4649–4667. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surman, S.L.; Crawford, J.; Dash, P.; Tonkonogy, S.L.; Thomas, P.G.; Hurwitz, J.L. Microbiome Shapes the T Cell Receptor Repertoire among CD4+CD8+ Thymocytes. Biomedicines 2022, 10, 3015. https://doi.org/10.3390/biomedicines10123015
Surman SL, Crawford J, Dash P, Tonkonogy SL, Thomas PG, Hurwitz JL. Microbiome Shapes the T Cell Receptor Repertoire among CD4+CD8+ Thymocytes. Biomedicines. 2022; 10(12):3015. https://doi.org/10.3390/biomedicines10123015
Chicago/Turabian StyleSurman, Sherri L., Jeremy Crawford, Pradyot Dash, Susan L. Tonkonogy, Paul G. Thomas, and Julia L. Hurwitz. 2022. "Microbiome Shapes the T Cell Receptor Repertoire among CD4+CD8+ Thymocytes" Biomedicines 10, no. 12: 3015. https://doi.org/10.3390/biomedicines10123015