
Temporal Evolution of Multiday, Epileptic Functional Networks Prior to Seizure Occurrence
 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
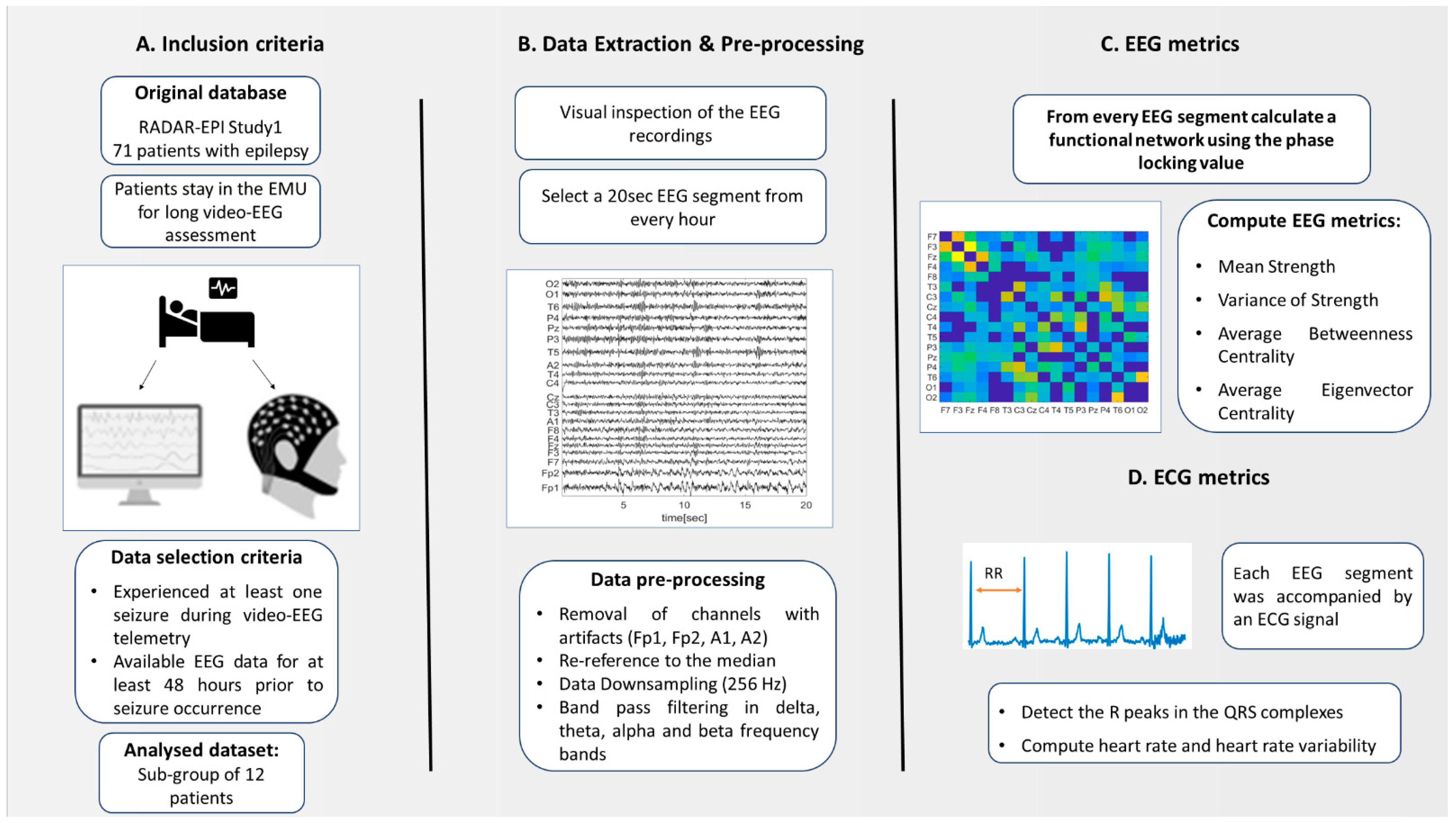
2. Materials and Methods
2.1. Participants Recruitment
2.2. Inclusion and Exlusion Criteria
2.3. Ethics Statement
2.4. EEG Measurements
2.4.1. Data Preprosessing
2.4.2. Functional Network
2.4.3. EEG Metrics
2.5. ECG Metrics
HR and HRV Metrics
2.6. Statistical Analysis
3. Results
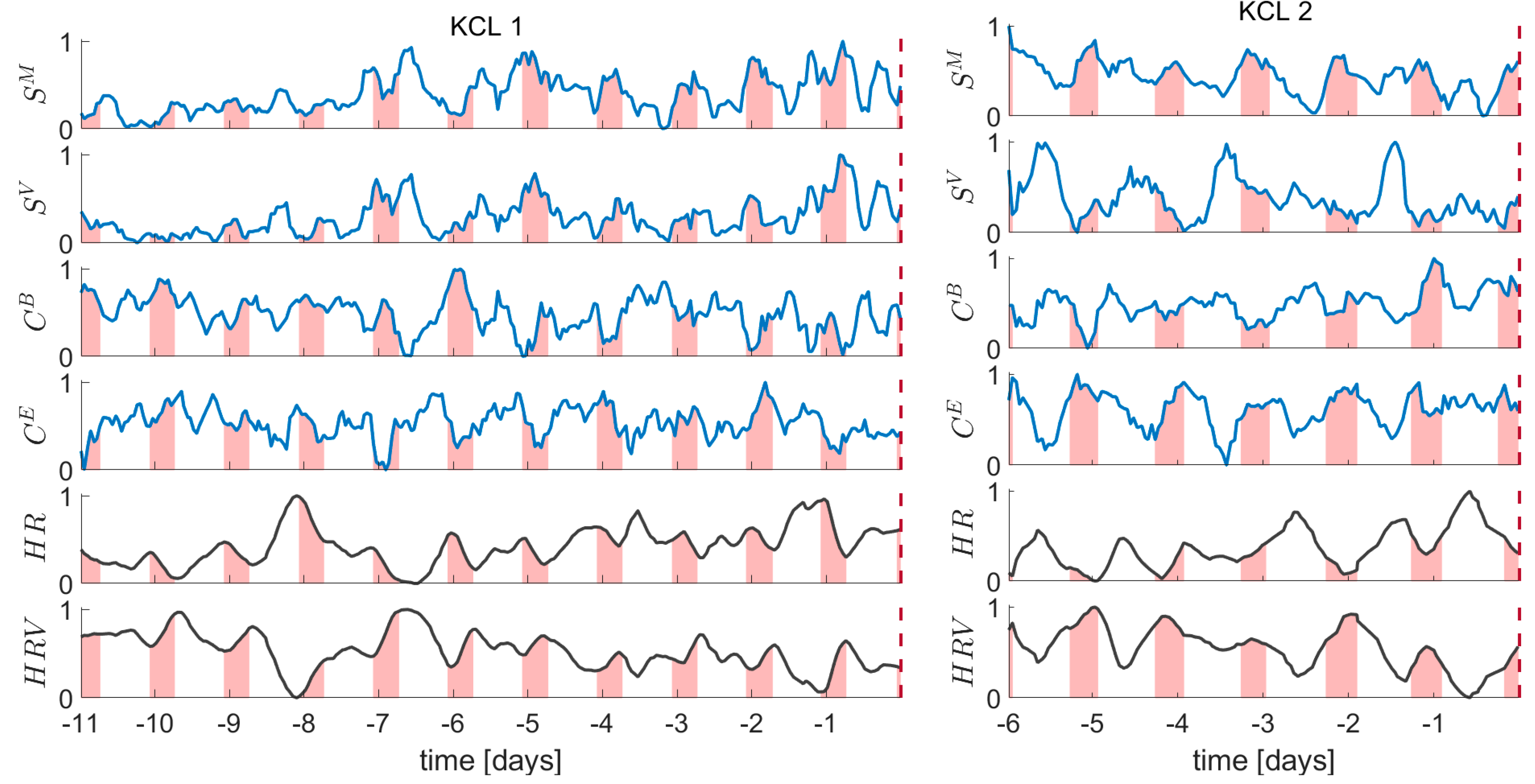
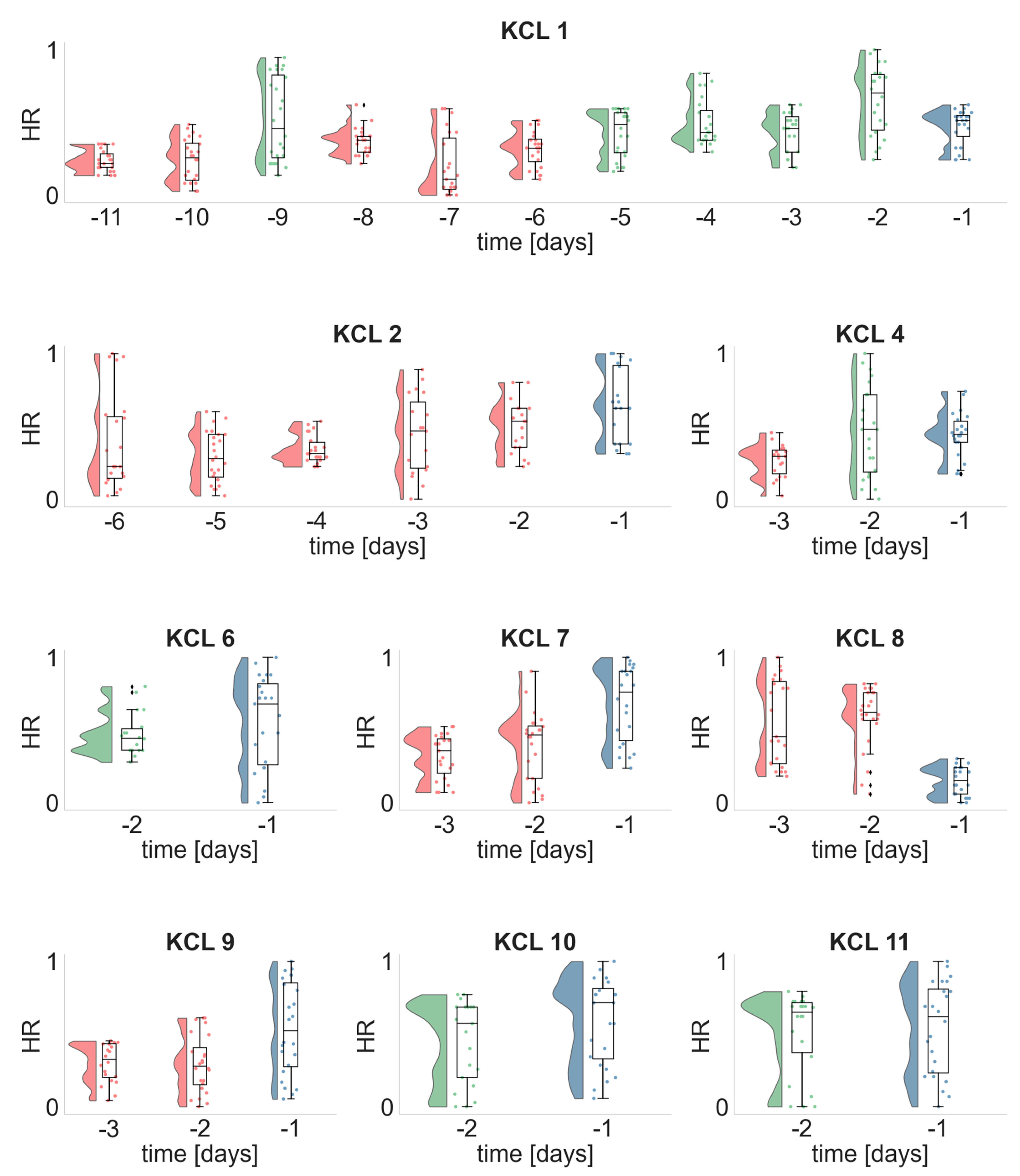
3.1. Temporal Evolution of the EEG and ECG Metrics
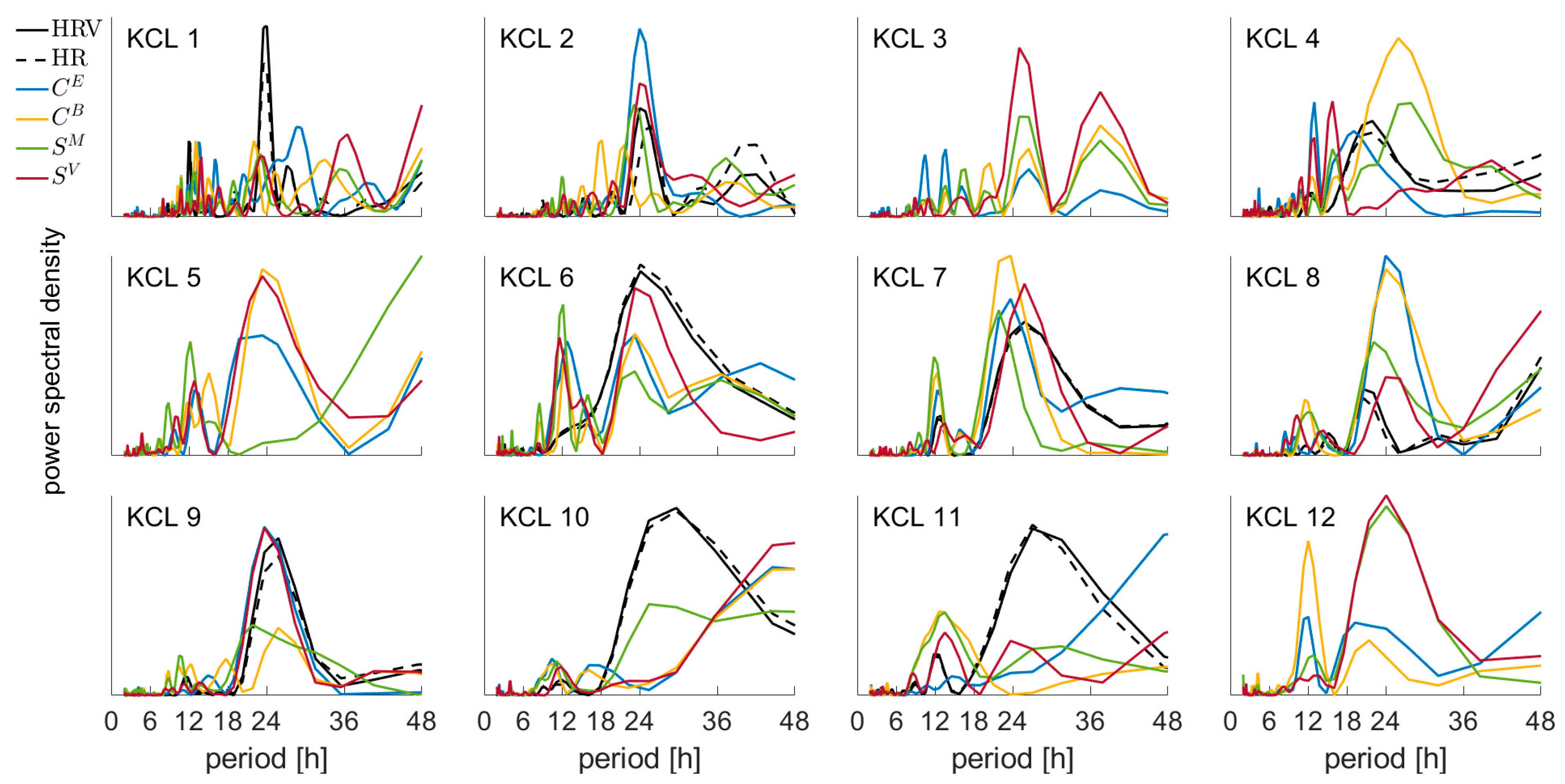
3.2. Periodicity of the EEG and ECG Metrics
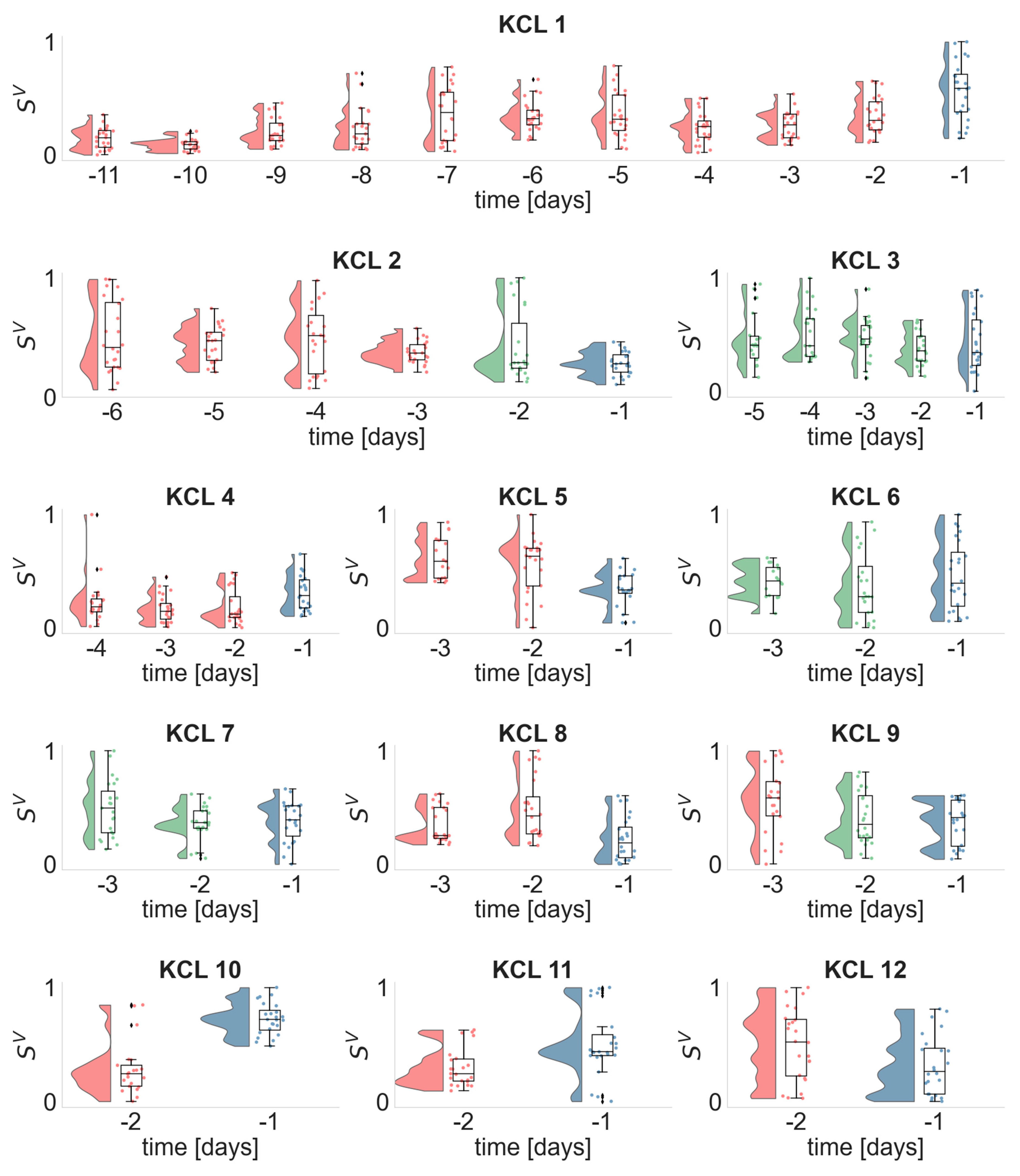
3.3. Daily Distributions of the EEG and ECG Metrics
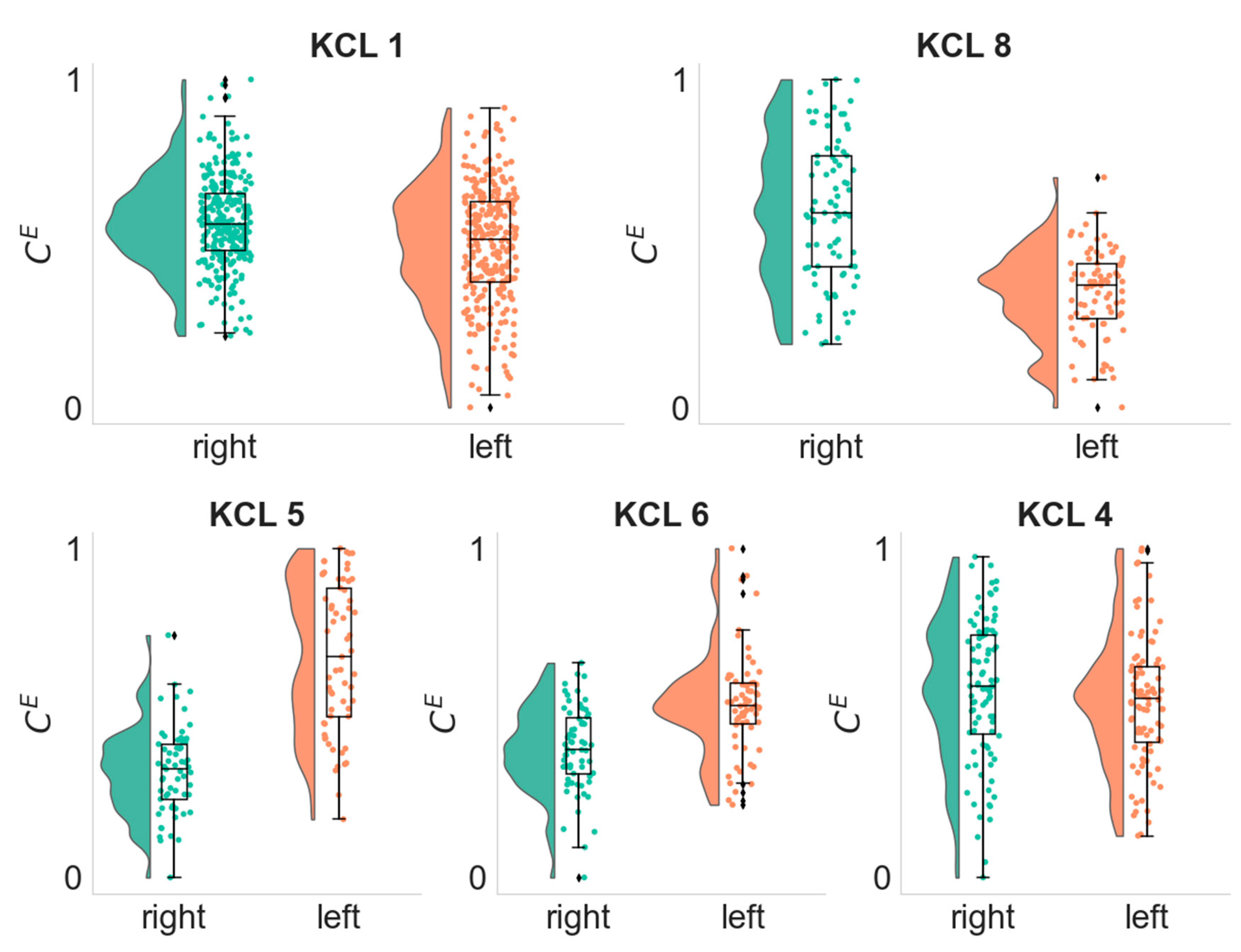
3.4. Lateralization of the Seizure Focus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McKenna, T.M.; McMullen, T.A.; Shlesinger, M.F. The brain as a dynamic physical system. Neuroscience 1994, 60, 587–605. [Google Scholar] [CrossRef]
- Da Silva, F.L.; Blanes, W.; Kalitzin, S.N.; Parra, J.; Suffczynski, P.; Velis, D.N. Epilepsies as dynamical diseases of brain systems: Basic models of the transition between normal and epileptic activity. Epilepsia 2003, 44, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.S.; Vickrey, B.G.; Gibson, P.; Hermann, B.; Penovich, P.; Scherer, A.; Walker, S. The impact of epilepsy from the patient’s perspective I. Descriptions and subjective perceptions. Epilepsy Res. 2000, 41, 39–51. [Google Scholar] [CrossRef]
- Dumanis, S.B.; French, J.A.; Bernard, C.; Worrell, G.A.; Fureman, B.E. Seizure forecasting from idea to reality. Outcomes of the my seizure gauge epilepsy innovation institute workshop. Eneuro 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, S.S. Neural networks in human epilepsy: Evidence of and implications for treatment. Epilepsia 2002, 43, 219–227. [Google Scholar] [CrossRef]
- Richardson, M.P. Large scale brain models of epilepsy: Dynamics meets connectomics. J. Neurol. Neurosurg. Psychiatry 2012, 83, 1238–1248. [Google Scholar] [CrossRef] [Green Version]
- Besson, P.; Bandt, S.K.; Proix, T.; Lagarde, S.; Jirsa, V.K.; Ranjeva, J.P.; Bartolomei, F.; Guye, M. Anatomic consistencies across epilepsies: A stereotactic-EEG informed high-resolution structural connectivity study. Brain 2017, 140, 2639–2652. [Google Scholar] [CrossRef] [Green Version]
- Bartolomei, F.; Lagarde, S.; Wendling, F.; McGonigal, A.; Jirsa, V.; Guye, M.; Bénar, C. Defining epileptogenic networks: Contribution of SEEG and signal analysis. Epilepsia 2017, 58, 1131–1147. [Google Scholar] [CrossRef] [Green Version]
- Sporns, O. Graph theory methods: Applications in brain networks. Dialogues Clin. Neurosci. 2018, 20, 111–121. [Google Scholar] [CrossRef]
- Bassett, D.S.; Zurn, P.; Gold, J.I. On the nature and use of models in network neuroscience. Nat. Rev. Neurosci. 2018, 19, 566–578. [Google Scholar] [CrossRef]
- Kramer, M.A.; Kolaczyk, E.D.; Kirsch, H.E. Emergent network topology at seizure onset in humans. Epilepsy Res. 2008, 79, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Schindler, K.A.; Bialonski, S.; Horstmann, M.T.; Elger, C.E.; Lehnertz, K. Evolving functional network properties and synchronizability during human epileptic seizures. Chaos Interdiscip. J. Nonlinear Sci. 2008, 18, 033119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhnert, M.T.; Elger, C.E.; Lehnertz, K. Long-term variability of global statistical properties of epileptic brain networks. Chaos Interdiscip. J. Nonlinear Sci. 2010, 20, 043126. [Google Scholar] [CrossRef]
- Lehnertz, K.; Ansmann, G.; Bialonski, S.; Dickten, H.; Geier, C.; Porz, S. Evolving networks in the human epileptic brain. Phys. D Nonlinear Phenom. 2014, 267, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Geier, C.; Lehnertz, K. Long-term variability of importance of brain regions in evolving epileptic brain networks. Chaos Interdiscip. J. Nonlinear Sci. 2017, 27, 043112. [Google Scholar] [CrossRef]
- Newman, M.E. The structure and function of complex networks. SIAM Rev. 2003, 45, 167–256. [Google Scholar] [CrossRef] [Green Version]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Bruno, E.; Böttcher, S.; Viana, P.F.; Amengual-Gual, M.; Joseph, B.; Epitashvili, N.; Dümpelmann, M.; Glasstetter, M.; Biondi, A.; Van Laerhoven, K.; et al. Wearable devices for seizure detection: Practical experiences and recommendations from the Wearables for Epilepsy and Research (WEAR) International Study Group. Epilepsia 2021, 62, 2307–2321. [Google Scholar] [CrossRef]
- Tass, P.; Rosenblum, M.G.; Weule, J.; Kurths, J.; Pikovsky, A.; Volkmann, J.; Schnitzler, A.; Freund, H.-J. Detection of n:m Phase Locking from Noisy Data: Application to Magnetoencephalography. Phys. Rev. Lett. 1998, 81, 3291. [Google Scholar] [CrossRef]
- Lachaux, J.P.; Rodriguez, E.; Martinerie, J.; Varela, F.J. Measuring phase synchrony in brain signals. Hum. Brain Mapp. 1999, 8, 194–208. [Google Scholar] [CrossRef]
- Mormann, F.; Lehnertz, K.; David, P.; Elger, C.E. Mean phase coherence as a measure for phase synchronization and its application to the EEG of epilepsy patients. Phys. D Nonlinear Phenom. 2000, 144, 358–369. [Google Scholar] [CrossRef]
- Bastos, A.M.; Schoffelen, J.M. A tutorial review of functional connectivity analysis methods and their interpretational pitfalls. Front. Syst. Neurosci. 2016, 8, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, T.; Schmitz, A. Improved surrogate data for nonlinearity tests. Phys. Rev. Lett. 1996, 77, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, T.; Schmitz, A. Surrogate time series. Physica D: Nonlinear Phenomena 2000, 142, 346–382. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Tompkins, W.J. A real-time QRS detection algorithm. IEEE Trans. Biomed. Eng. 1985, 3, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, F.; Ginsberg, J.P. An overview of heart rate variability metrics and norms. Front. Public Health 2017, 5, 258. [Google Scholar] [CrossRef] [Green Version]
- Shaffer, F.; Meehan, Z.M.; Zerr, C.L. A critical review of ultra-short-term heart rate variability norms research. Front. Neurosci. 2020, 14, 594880. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Allen, M.; Poggiali, D.; Whitaker, K.; Marshall, T.R.; Kievit, R.A. Raincloud plots: A multi-platform tool for robust data visualization. Wellcome Open Res. 2019, 4, 63. [Google Scholar] [CrossRef] [Green Version]
- Khambhati, A.N.; Shafi, A.; Rao, V.R.; Chang, E.F. Long-term brain network reorganization predicts responsive neurostimulation outcomes for focal epilepsy. Sci. Transl. Med. 2021, 13, eabf6588. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, F.A.; Woldman, W.; FitzGerald, T.H.; Elwes, R.D.; Nashef, L.; Terry, J.R.; Richardson, M.P. Revealing a brain network endophenotype in families with idiopathic generalised epilepsy. PLoS ONE 2014, 9, e110136. [Google Scholar] [CrossRef] [PubMed]
- Pegg, E.J.; Taylor, J.R.; Laiou, P.; Richardson, M.; Mohanraj, R. Interictal electroencephalographic functional network topology in drug-resistant and well-controlled idiopathic generalized epilepsy. Epilepsia 2021, 62, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.A.; Richardson, M.P.; Abela, E.; Rummel, C.; Schindler, K.; Goodfellow, M.; Terry, J.R. An optimal strategy for epilepsy surgery: Disruption of the rich-club? PLoS Comput. Biol. 2017, 13, e1005637. [Google Scholar] [CrossRef] [Green Version]
- Laiou, P.; Avramidis, E.; Lopes, M.A.; Abela, E.; Müller, M.; Akman, O.E.; Richardson, M.P.; Rummel, C.; Schindler, K.; Goodfellow, M. Quantification and selection of ictogenic zones in epilepsy surgery. Front. Neurol. 2019, 10, 1045. [Google Scholar] [CrossRef]
- Geier, C.; Bialonski, S.; Elger, C.E.; Lehnertz, K. How important is the seizure onset zone for seizure dynamics? Seizure 2015, 25, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Jiruska, P.; De Curtis, M.; Jefferys, J.G.; Schevon, C.A.; Schiff, S.J.; Schindler, K. Synchronization and desynchronization in epilepsy: Controversies and hypotheses. J. Physiol. 2013, 591, 787–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Van Quyen, M.; Soss, J.; Navarro, V.; Robertson, R.; Chavez, M.; Baulac, M.; Martinerie, J. Preictal state identification by synchronization changes in long-term intracranial EEG recordings. Clin. Neurophysiol. 2005, 116, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Mormann, F.; Kreuz, T.; Andrzejak, R.G.; David, P.; Lehnertz, K.; Elger, C.E. Epileptic seizures are preceded by a decrease in synchronization. Epilepsy Res. 2003, 53, 173–185. [Google Scholar] [CrossRef]
- Coito, A.; Plomp, G.; Genetti, M.; Abela, E.; Wiest, R.; Seeck, M.; Michel, C.M.; Vulliemoz, S. Dynamic directed interictal connectivity in left and right temporal lobe epilepsy. Epilepsia 2015, 56, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Karoly, P.J.; Stirling, R.E.; Freestone, D.R.; Nurse, E.S.; Maturana, M.I.; Halliday, A.J.; Neal, A.; Gregg, N.M.; Brinkmann, B.H.; Richardson, M.P.; et al. Multiday cycles of heart rate are associated with seizure likelihood: An observational cohort study. EBioMedicine 2021, 72, 103619. [Google Scholar] [CrossRef]
- Gregg, N.M.; Attia, T.P.; Nasseri, M.; Joseph, B.; Karoly, P.J.; Cui, J.; Stirling, R.E.; Viana, P.F.; Richner, T.J.; Nurse, E.S.; et al. Multimodal wearable sensors inform cycles of seizure risk. medRxiv 2022. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient ID | Age | Sex | Seizure Focus | Epilepsy Type | Number of Analyzed Days Prior to Seizure Occurrence | Number of Analyzed EEG Epochs | Number of Analyzed ECG Epochs |
|---|---|---|---|---|---|---|---|
| KCL 1 | 65 | male | right | RTLE | 11 | 262 | 258 |
| KCL 2 | 32 | female | unclear | TLE | 6 | 142 | 128 |
| KCL 3 | 38 | female | unclear | FLE | 5 | 110 | 0 |
| KCL 4 | 28 | male | left | LTLE | 4 | 91 | 64 |
| KCL 5 | 37 | female | left | LTLE | 3 | 61 | 0 |
| KCL 6 | 35 | male | left | LTLE | 3 | 64 | 41 |
| KCL 7 | 52 | male | unclear | TLE | 3 | 70 | 69 |
| KCL 8 | 47 | female | right | RTLE | 3 | 71 | 71 |
| KCL 9 | 43 | male | unclear | FLE | 3 | 70 | 68 |
| KCL 10 | 39 | male | unclear | IGE | 2 | 45 | 45 |
| KCL 11 | 22 | male | unclear | TLE | 2 | 47 | 46 |
| KCL 12 | 55 | male | unclear | TLE | 2 | 48 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laiou, P.; Biondi, A.; Bruno, E.; Viana, P.F.; Winston, J.S.; Rashid, Z.; Ranjan, Y.; Conde, P.; Stewart, C.; Sun, S.; et al. Temporal Evolution of Multiday, Epileptic Functional Networks Prior to Seizure Occurrence. Biomedicines 2022, 10, 2662. https://doi.org/10.3390/biomedicines10102662
Laiou P, Biondi A, Bruno E, Viana PF, Winston JS, Rashid Z, Ranjan Y, Conde P, Stewart C, Sun S, et al. Temporal Evolution of Multiday, Epileptic Functional Networks Prior to Seizure Occurrence. Biomedicines. 2022; 10(10):2662. https://doi.org/10.3390/biomedicines10102662
Chicago/Turabian StyleLaiou, Petroula, Andrea Biondi, Elisa Bruno, Pedro F. Viana, Joel S. Winston, Zulqarnain Rashid, Yatharth Ranjan, Pauline Conde, Callum Stewart, Shaoxiong Sun, and et al. 2022. "Temporal Evolution of Multiday, Epileptic Functional Networks Prior to Seizure Occurrence" Biomedicines 10, no. 10: 2662. https://doi.org/10.3390/biomedicines10102662