
Ethanol Effects on Early Developmental Stages Studied Using the Zebrafish

{kind=link}
Abstract
:1. Fetal Alcohol Spectrum Disorder
2. Use of Zebrafish as an FASD Model
3. Early Zebrafish Development
3.1. Blastula Stage
3.2. Gastrulation
3.3. Pluripotency Circuit
4. Consequences of Ethanol Exposure during Early Zebrafish Development
4.1. Pluripotency Gene Expression Defects
4.1.1. sox2
4.1.2. elf3
4.1.3. pou5f1
4.2. Epiboly Defects
4.3. Sonic Hedgehog
4.4. Cdon
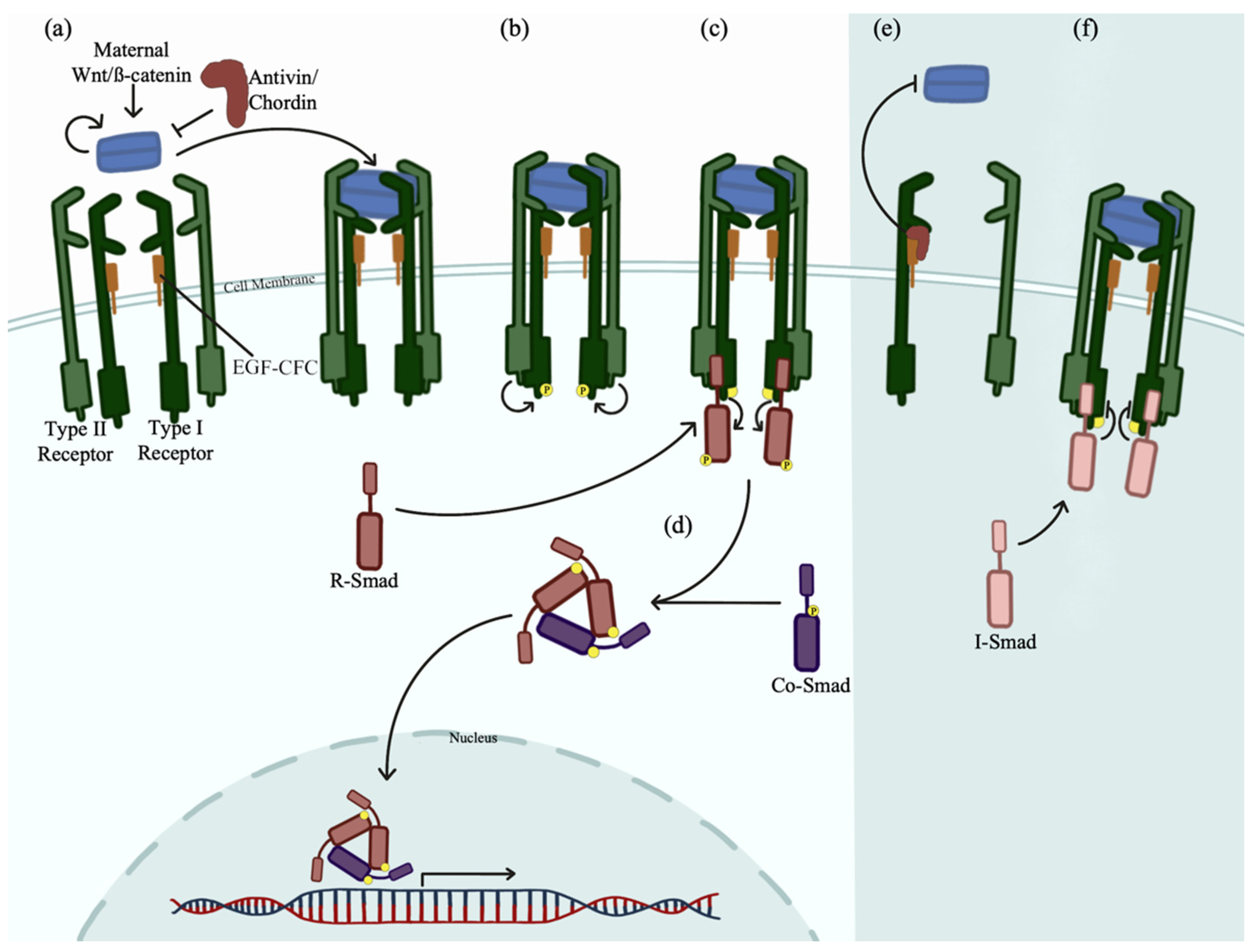
5. Nodal and Bmp Gradients
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wallen, E.; Auvinen, P.; Kaminen-Ahola, N. The Effects of Early Prenatal Alcohol Exposure on Epigenome and Embryonic Development. Genes 2021, 12, 1095. [Google Scholar] [CrossRef]
- Bilotta, J.; Barnett, J.A.; Hancock, L.; Saszik, S. Ethanol exposure alters zebrafish development: A novel model of fetal alcohol syndrome. Neurotoxicol. Teratol. 2004, 26, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Probst, C.; Gmel, G.; Rehm, J.; Burd, L.; Popova, S. Global Prevalence of Fetal Alcohol Spectrum Disorder among Children and Youth: A Systematic Review and Meta-analysis. JAMA Pediatr. 2017, 171, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Denny, C.H.; Acero, C.S.; Naimi, T.S.; Kim, S.Y. Consumption of Alcohol Beverages and Binge Drinking among Pregnant Women Aged 18–44 Years-United States, 2015–2017. Morbid. Mortal. Wkly. Rep. 2019, 68, 365–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidharan, P.; Sarmah, S.; Marrs, J.A. Zebrafish retinal defects induced by ethanol exposure are rescued by retinoic acid and folic acid supplement. Alcohol 2015, 49, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Sarmah, S.; Muralidharan, P.; Curtis, C.L.; McClintick, J.N.; Buente, B.B.; Holdgrafer, D.J.; Ogbeifun, O.; Olorungbounmi, O.C.; Patino, L.; Lucas, R.; et al. Ethanol exposure disrupts extraembryonic microtubule cytoskeleton and embryonic blastomere cell adhesion, producing epiboly and gastrulation defects. Biol. Open 2013, 2, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Serrano, M.; Han, M.; Brinez, P.; Linask, K.K. Fetal alcohol syndrome: Cardiac birth defects in mice and prevention with folate. Am. J. Obstet. Gynecol. 2010, 203, 75.e7–75.e15. [Google Scholar] [CrossRef]
- da Silva, J.P.; Luchiari, A.C. Embryonic ethanol exposure on zebrafish early development. Brain Behav. 2021, 11, e02062. [Google Scholar] [CrossRef]
- Fernandes, Y.; Lovely, C.B. Zebrafish models of fetal alcohol spectrum disorders. Genesis 2021, 59, e23460. [Google Scholar] [CrossRef]
- Lovely, C.B.; Fernandes, Y.; Eberhart, J.K. Fishing for Fetal Alcohol Spectrum Disorders: Zebrafish as a Model for Ethanol Teratogenesis. Zebrafish 2016, 13, 391–398. [Google Scholar] [CrossRef]
- Paranjpe, S.S.; Veenstra, G.J. Establishing pluripotency in early development. Biochim. Biophys. Acta 2015, 1849, 626–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akdogan-Ozdilek, B.; Duval, K.L.; Goll, M.G. Chromatin dynamics at the maternal to zygotic transition: Recent advances from the zebrafish model. F1000Res 2020, 9, 299. [Google Scholar] [CrossRef] [PubMed]
- Schauer, A.; Pinheiro, D.; Hauschild, R.; Heisenberg, C.P. Zebrafish embryonic explants undergo genetically encoded self-assembly. Elife 2020, 9, e55190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leichsenring, M.; Maes, J.; Mossner, R.; Driever, W.; Onichtchouk, D. Pou5f1 transcription factor controls zygotic gene activation in vertebrates. Science 2013, 341, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Bruce, A.E.E.; Heisenberg, C.P. Mechanisms of zebrafish epiboly: A current view. Curr. Top. Dev. Biol. 2020, 136, 319–341. [Google Scholar] [CrossRef]
- Gilbert, S.F.; Barresi, M.J.F. Developmental Biology 11th Edition; Sinauer Associates Inc.: Sunderland, MA, USA, 2016. [Google Scholar]
- Chen, S.; Kimelman, D. The role of the yolk syncytial layer in germ layer patterning in zebrafish. Development 2000, 127, 4681–4689. [Google Scholar] [CrossRef]
- Chambers, I.; Tomlinson, S.R. The transcriptional foundation of pluripotency. Development 2009, 136, 2311–2322. [Google Scholar] [CrossRef] [Green Version]
- Ogony, J.W.; Malahias, E.; Vadigepalli, R.; Anni, H. Ethanol alters the balance of Sox2, Oct4, and Nanog expression in distinct subpopulations during differentiation of embryonic stem cells. Stem Cells Dev. 2013, 22, 2196–2210. [Google Scholar] [CrossRef] [Green Version]
- Robles, V.; Marti, M.; Belmonte, J.C.I. Study of pluripotency markers in zebrafish embryos and transient embryonic stem cell cultures. Zebrafish 2011, 8, 57–63. [Google Scholar] [CrossRef]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodda, D.J.; Chew, J.L.; Lim, L.H.; Loh, Y.H.; Wang, B.; Ng, H.H.; Robson, P. Transcriptional regulation of Nanog by Oct4 and Sox2. J. Biol. Chem. 2005, 280, 24731–24737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Cui, W. Sox2, a key factor in the regulation of pluripotency and neural differentiation. World J. Stem Cells 2014, 6, 305–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmah, S.; Srivastava, R.; McClintick, J.N.; Janga, S.C.; Edenberg, H.J.; Marrs, J.A. Embryonic ethanol exposure alters expression of sox2 and other early transcripts in zebrafish, producing gastrulation defects. Sci. Rep. 2020, 10, 3951. [Google Scholar] [CrossRef] [Green Version]
- Okuda, Y.; Yoda, H.; Uchikawa, M.; Furutani-Seiki, M.; Takeda, H.; Kondoh, H.; Kamachi, Y. Comparative genomic and expression analysis of group B1 sox genes in zebrafish indicates their diversification during vertebrate evolution. Dev. Dyn. 2006, 235, 811–825. [Google Scholar] [CrossRef]
- Lachnit, M.; Kur, E.; Driever, W. Alterations of the cytoskeleton in all three embryonic lineages contribute to the epiboly defect of Pou5f1/Oct4 deficient MZspg zebrafish embryos. Dev. Biol. 2008, 315, 1–17. [Google Scholar] [CrossRef] [Green Version]
- E!Ensembl Gene: ELF3 ENSG00000163435. Available online: https://useast.ensembl.org/Homo_sapiens/Gene/ExpressionAtlas?db=core;g=ENSG00000163435;r=1:202007945-202017183 (accessed on 27 July 2022).
- Kageyama, S.; Liu, H.; Nagata, M.; Aoki, F. The role of ETS transcription factors in transcription and development of mouse preimplantation embryos. Biochem. Biophys. Res. Commun. 2006, 344, 675–679. [Google Scholar] [CrossRef]
- Oliver, J.R.; Kushwah, R.; Hu, J. Multiple roles of the epithelium-specific ETS transcription factor, ESE-1, in development and disease. Lab. Investig. 2012, 92, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Sarmah, S.; Hawkins, M.R.; Manikandan, P.; Farrell, M.; James, A. Marrs. Elf3 deficiency during zebrafish development alters extracellular matrix organization and disrupts tissue morphogenesis. PLoS ONE, 2022; In Press. [Google Scholar]
- Wang, Z.; Yin, P.; Sun, Y.; Na, L.; Gao, J.; Wang, W.; Zhao, C. LGR4 maintains HGSOC cell epithelial phenotype and stem-like traits. Gynecol. Oncol. 2020, 159, 839–849. [Google Scholar] [CrossRef]
- Park, S.W.; Do, H.J.; Choi, W.; Lim, D.S.; Park, K.H.; Kim, J.H. Epithelium-specific ETS transcription factor-1 regulates NANOG expression and inhibits NANOG-induced proliferation of human embryonic carcinoma cells. Biochimie 2021, 186, 33–42. [Google Scholar] [CrossRef]
- Onichtchouk, D.; Geier, F.; Polok, B.; Messerschmidt, D.M.; Mossner, R.; Wendik, B.; Song, S.; Taylor, V.; Timmer, J.; Driever, W. Zebrafish Pou5f1-dependent transcriptional networks in temporal control of early development. Mol. Syst. Biol. 2010, 6, 354. [Google Scholar] [CrossRef] [PubMed]
- Onichtchouk, D. Pou5f1/oct4 in pluripotency control: Insights from zebrafish. Genesis 2012, 50, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Eckerle, S.; Onichtchouk, D.; Marrs, J.A.; Nitschke, R.; Driever, W. Pou5f1-dependent EGF expression controls E-cadherin endocytosis, cell adhesion, and zebrafish epiboly movements. Dev. Cell 2013, 24, 486–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babb, S.G.; Marrs, J.A. E-cadherin regulates cell movements and tissue formation in early zebrafish embryos. Dev. Dyn. 2004, 230, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Kane, D.A.; McFarland, K.N.; Warga, R.M. Mutations in half baked/E-cadherin block cell behaviors that are necessary for teleost epiboly. Development 2005, 132, 1105–1116. [Google Scholar] [CrossRef] [Green Version]
- Emond, M.R.; Biswas, S.; Blevins, C.J.; Jontes, J.D. A complex of Protocadherin-19 and N-cadherin mediates a novel mechanism of cell adhesion. J. Cell Biol. 2011, 195, 1115–1121. [Google Scholar] [CrossRef] [Green Version]
- Mao, H.; Diehl, A.M.; Li, Y.X. Sonic hedgehog ligand partners with caveolin-1 for intracellular transport. Lab. Investig. 2009, 89, 290–300. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.X.; Yang, H.T.; Zdanowicz, M.; Sicklick, J.K.; Qi, Y.; Camp, T.J.; Diehl, A.M. Fetal alcohol exposure impairs Hedgehog cholesterol modification and signaling. Lab. Investig. 2007, 87, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Hong, M.; Krauss, R.S. Ethanol itself is a holoprosencephaly-inducing teratogen. PLoS ONE 2017, 12, e0176440. [Google Scholar] [CrossRef] [Green Version]
- Sidik, A.; Dixon, G.B.; Kirby, H.G.; Eberhart, J.K. Gene-environment interactions characterized by single embryo transcriptomics. BioRxiv 2020, 805556. [Google Scholar] [CrossRef]
- Slavotinek, A.M. Chapter 128 Dysmorphology-Nelson Textbook of Pediatrics; Elsevier Inc.: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Hong, M.; Christ, A.; Christa, A.; Willnow, T.E.; Krauss, R.S. Cdon mutation and fetal alcohol converge on Nodal signaling in a mouse model of holoprosencephaly. Elife 2020, 9, e60351. [Google Scholar] [CrossRef] [PubMed]
- Eberhart, J.K.; Parnell, S.E. The Genetics of Fetal Alcohol Spectrum Disorders. Alcohol Clin. Exp. Res. 2016, 40, 1154–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loucks, E.J.; Ahlgren, S.C. Deciphering the role of Shh signaling in axial defects produced by ethanol exposure. Birth Defects Res. A Clin. Mol. Teratol. 2009, 85, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Echevarria-Andino, M.L.; Allen, B.L. The hedgehog co-receptor BOC differentially regulates SHH signaling during craniofacial development. Development 2020, 147, dev189076. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.I.; Krauss, R.S. Cdon Mutation and Fetal Ethanol Exposure Synergize to Produce Midline Signaling Defects and Holoprosencephaly Spectrum Disorders in Mice. PLoS Genet. 2012, 8, e1002999. [Google Scholar] [CrossRef] [Green Version]
- Powell, D.R.; Williams, J.S.; Hernandez-Lagunas, L.; Salcedo, E.; O’Brien, J.H.; Artinger, K.B. Cdon promotes neural crest migration by regulating N-cadherin localization. Dev. Biol. 2015, 407, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Cardozo, M.J.; Sanchez-Arrones, L.; Sandonis, A.; Sanchez-Camacho, C.; Gestri, G.; Wilson, S.W.; Guerrero, I.; Bovolenta, P. Cdon acts as a Hedgehog decoy receptor during proximal-distal patterning of the optic vesicle. Nat. Commun. 2014, 5, 4272. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Lin, X.; Cai, Z.; Zhang, Z.; Han, C.; Jia, S.; Meng, A.; Wang, Q. Global identification of SMAD2 target genes reveals a role for multiple co-regulatory factors in zebrafish early gastrulas. J. Biol. Chem. 2011, 286, 28520–28532. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.Y.; Wang, H.P.; Zhu, Z.Y.; Sun, Y.H. Transcriptional factors smad1 and smad9 act redundantly to mediate zebrafish ventral specification downstream of smad5. J. Biol. Chem. 2014, 289, 6604–6618. [Google Scholar] [CrossRef] [Green Version]
- Rogers, K.W.; Muller, P. Nodal and BMP dispersal during early zebrafish development. Dev. Biol. 2019, 447, 14–23. [Google Scholar] [CrossRef]
- Crick, F. Diffusion in embryogenesis. Nature 1970, 225, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, D.; Heisenberg, C.P. Zebrafish gastrulation: Putting fate in motion. Curr. Top. Dev. Biol. 2020, 136, 343–375. [Google Scholar] [CrossRef] [PubMed]
- Troilo, H.; Zuk, A.V.; Tunnicliffe, R.B.; Wohl, A.P.; Berry, R.; Collins, R.F.; Jowitt, T.A.; Sengle, G.; Baldock, C. Nanoscale structure of the BMP antagonist chordin supports cooperative BMP binding. Proc. Natl. Acad. Sci. USA 2014, 111, 13063–13068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marlow, F.L. Setting up for gastrulation in zebrafish. Curr. Top. Dev. Biol. 2020, 136, 33–83. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Gilligan, P.C.; Lim, S.; Tran, L.D.; Winkler, S.; Philp, R.; Sampath, K. An essential role for maternal control of Nodal signaling. Elife 2013, 2, e00683. [Google Scholar] [CrossRef] [PubMed]
- Massague, J. TGFbeta signalling in context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massague, J.; Chen, Y.G. Controlling TGF-beta signaling. Genes. Dev. 2000, 14, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.T.; Joubin, K.; Cheng, S.; Aanstad, P.; Herwig, R.; Clark, M.; Lehrach, H.; Schier, A.F. Nodal signaling activates differentiation genes during zebrafish gastrulation. Dev. Biol. 2007, 304, 525–540. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Novoselov, V.; Celeste, A.J.; Wolfman, N.M.; ten Dijke, P.; Kuehn, M.R. Nodal signaling uses activin and transforming growth factor-beta receptor-regulated Smads. J. Biol. Chem. 2001, 276, 656–661. [Google Scholar] [CrossRef] [Green Version]
- Guglielmo, G.M.D.; Roy, C.L.; Goodfellow, A.F.; Wrana, J.L. Distinct endocytic pathways regulate TGF-beta receptor signalling and turnover. Nat. Cell Biol. 2003, 5, 410–421. [Google Scholar] [CrossRef]
- Dick, A.; Mayr, T.; Bauer, H.; Meier, A.; Hammerschmidt, M. Cloning and characterization of zebrafish smad2, smad3 and smad4. Gene 2000, 246, 69–80. [Google Scholar] [CrossRef]
- Schier, A.F. Nodal morphogens. Cold Spring Harb. Perspect. Biol. 2009, 1, a003459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meno, C.; Gritsman, K.; Ohishi, S.; Ohfuji, Y.; Heckscher, E.; Mochida, K.; Shimono, A.; Kondoh, H.; Talbot, W.S.; Robertson, E.J.; et al. Mouse Lefty2 and zebrafish antivin are feedback inhibitors of nodal signaling during vertebrate gastrulation. Mol. Cell 1999, 4, 287–298. [Google Scholar] [CrossRef]
- Miyazawa, K.; Miyazono, K. Regulation of TGF-beta Family Signaling by Inhibitory Smads. Cold Spring Harb. Perspect. Biol. 2017, 9, a022095. [Google Scholar] [CrossRef] [Green Version]
- Conidi, A.; Cazzola, S.; Beets, K.; Coddens, K.; Collart, C.; Cornelis, F.; Cox, L.; Joke, D.; Dobreva, M.P.; Dries, R.; et al. Few Smad proteins and many Smad-interacting proteins yield multiple functions and action modes in TGFbeta/BMP signaling in vivo. Cytokine Growth Factor Rev. 2011, 22, 287–300. [Google Scholar] [CrossRef]
- Park, D.S.; Yoon, G.H.; Kim, E.Y.; Lee, T.; Kim, K.; Lee, P.C.; Chang, E.J.; Choi, S.C. Wip1 regulates Smad4 phosphorylation and inhibits TGF-beta signaling. EMBO Rep. 2020, 21, e48693. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shao, M.; Wang, L.; Liu, Z.; Gao, M.; Liu, C.; Zhang, H. Ethanol exposure affects cell movement during gastrulation and induces split axes in zebrafish embryos. Int. J. Dev. Neurosci. 2010, 28, 283–288. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manikandan, P.; Sarmah, S.; Marrs, J.A. Ethanol Effects on Early Developmental Stages Studied Using the Zebrafish. Biomedicines 2022, 10, 2555. https://doi.org/10.3390/biomedicines10102555
Manikandan P, Sarmah S, Marrs JA. Ethanol Effects on Early Developmental Stages Studied Using the Zebrafish. Biomedicines. 2022; 10(10):2555. https://doi.org/10.3390/biomedicines10102555
Chicago/Turabian StyleManikandan, Priyadharshini, Swapnalee Sarmah, and James A. Marrs. 2022. "Ethanol Effects on Early Developmental Stages Studied Using the Zebrafish" Biomedicines 10, no. 10: 2555. https://doi.org/10.3390/biomedicines10102555