
Chemerin as a Potential Marker of Resolution of Inflammation in COVID-19 Infection
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Main Clinical and Biochemical Results
3.2. Biomarkers in Severe vs. Non-Severe Illness
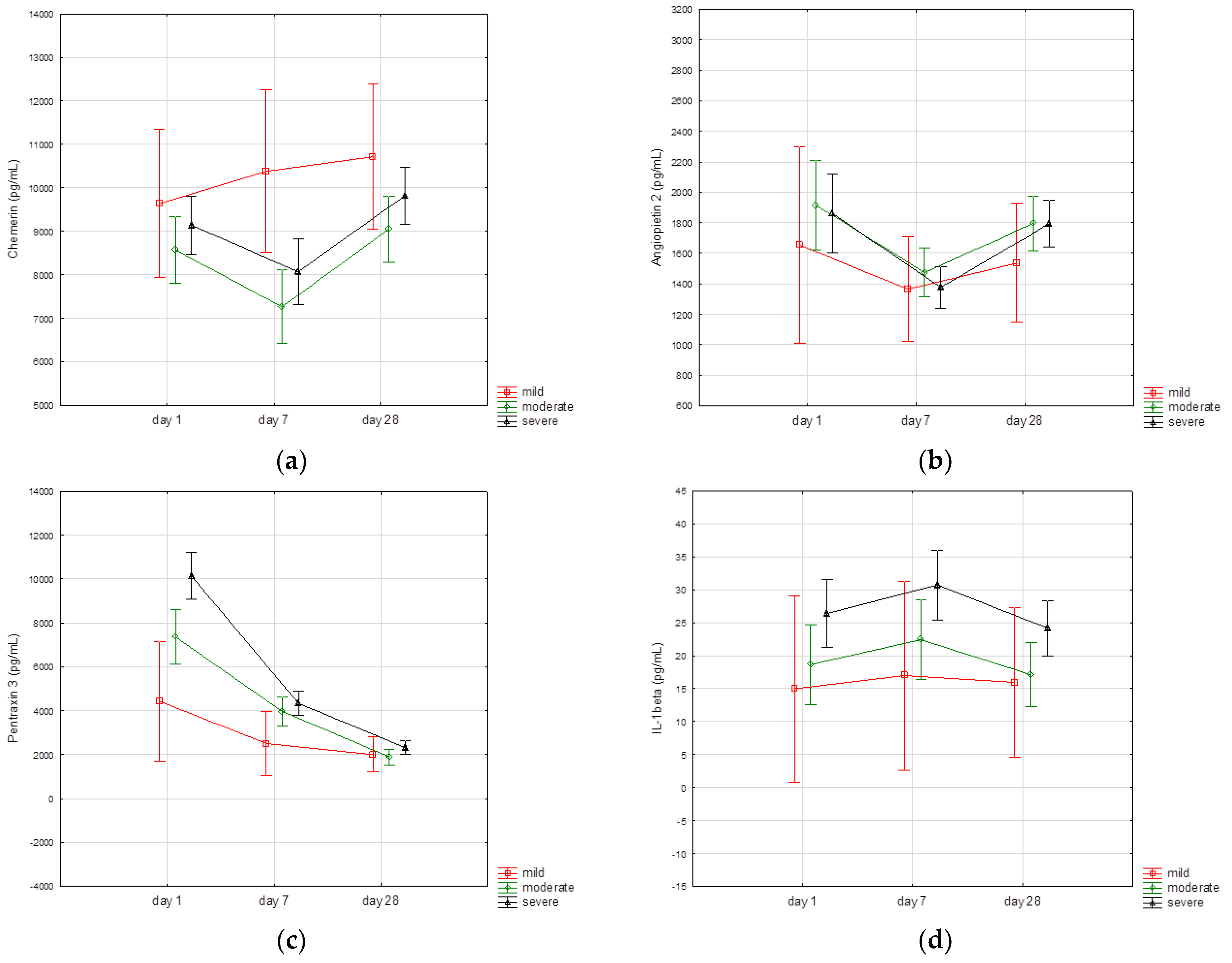
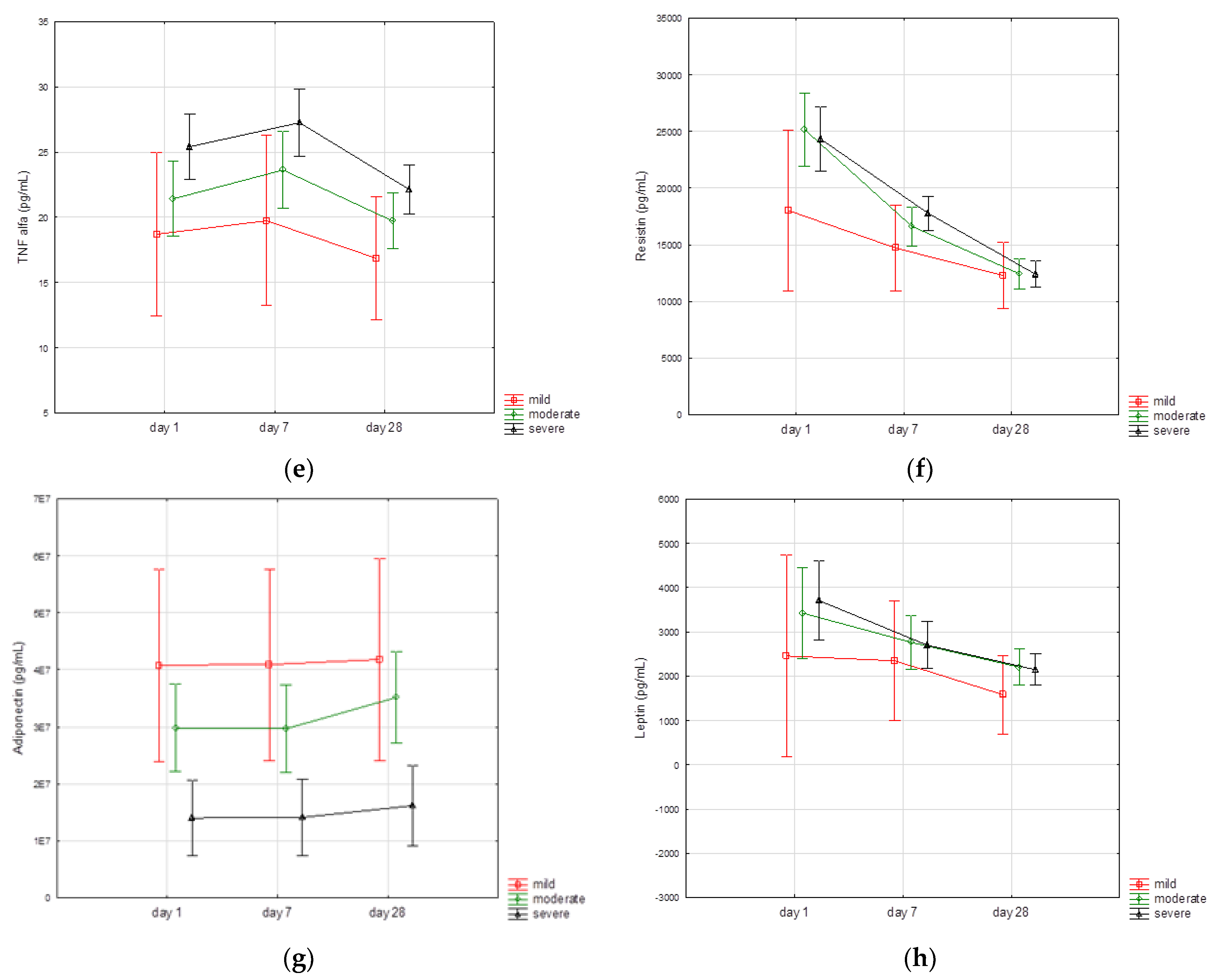
3.3. Biomarkers Kinetics
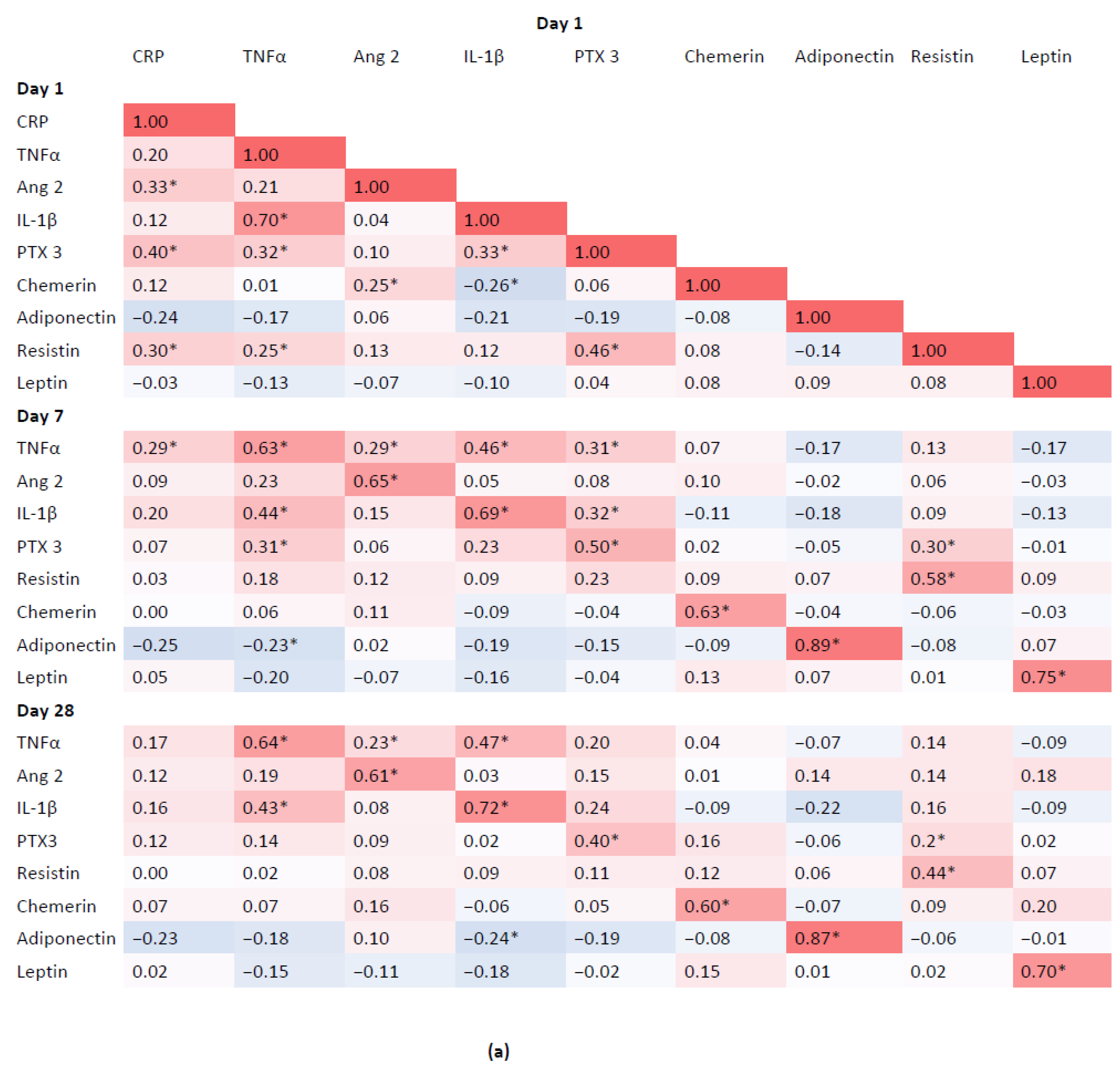
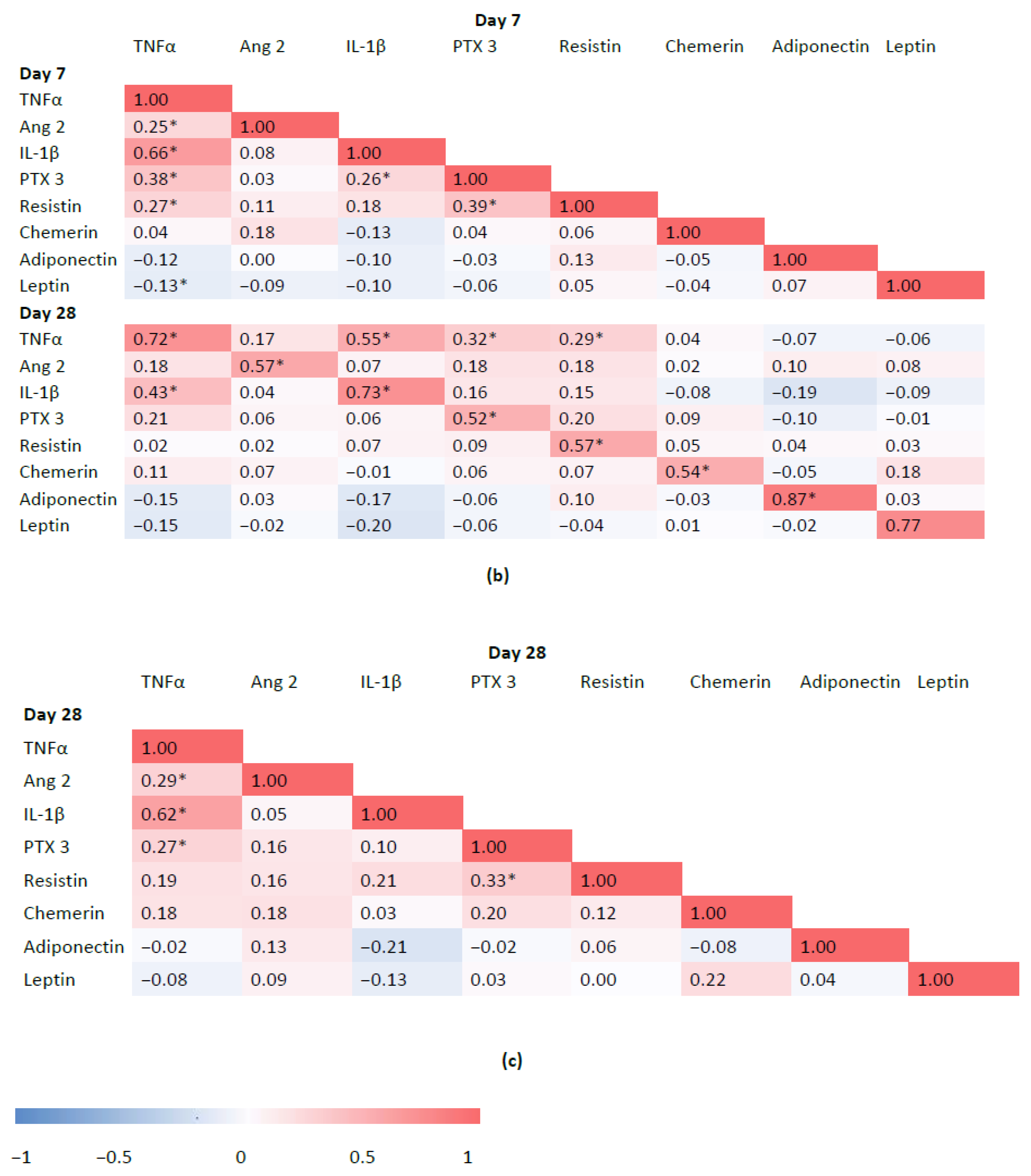
3.4. Associations between Inflammatory Biomarkers and Adipokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trombetta, A.C.; Farias, G.B.; Gomes, A.M.C.; Godinho-Santos, A.; Rosmaninho, P.; Conceição, C.M.; Laia, J.; Santos, D.F.; Almeida, A.R.M.; Mota, C.; et al. Severe COVID-19 Recovery Is Associated with Timely Acquisition of a Myeloid Cell Immune-Regulatory Phenotype. Front. Immunol. 2021, 12, 691725. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, S.; Herbert, J.A.; McNamara, P.S.; Hedrich, C.M. COVID-19: Immunology and treatment options. Clin. Immunol. 2020, 215, 108448. [Google Scholar] [CrossRef]
- Tam, V.C.; Quehenberger, O.; Oshansky, C.M.; Suen, R.; Armando, A.M.; Treuting, P.M.; Thomas, P.G.; Dennis, E.A.; Aderem, A. Lipidomic profiling of influenza infection identifies mediators that induce and resolve inflammation. Cell 2013, 154, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. HLH Across Speciality Collaboration, UK. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Merad, M.; Martin, J.C. Pathological inflammation in patients with COVID-19: A key role for monocytes and macrophages. Nat. Rev. Immunol. 2020, 20, 355–362. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N.; Dalli, J. The resolution code of acute inflammation: Novel pro-resolving lipid mediators in resolution. Semin. Immunol. 2015, 27, 200–215. [Google Scholar] [CrossRef] [Green Version]
- de Moraes, C.R.P.; de Borba Junior, I.; de Lima, F.; Barbosa, M.S.; Huber, S.C.; Palma, A.C.; Nunes, T.A.; Ulaf, R.G.; Ribeiro, L.C.; Bernandes, A.F.; et al. Circulating levels of Ang/Tie2 and VEGF-a pathway mediators are associated with clinical severity, endothelial barrier disruption and coagulation activation in COVID-19. Blood 2021, 138, 2073. [Google Scholar] [CrossRef]
- Yoshimura, T.; Oppenheim, J.J. Chemerin reveals its chimeric nature. J. Exp. Med. 2008, 205, 2187–2190. [Google Scholar] [CrossRef] [Green Version]
- Cash, J.L.; Norling, L.V.; Perretti, M. Resolution of inflammation: Targeting GPCRs that interact with lipids and peptides. Drug. Discov. Today 2014, 19, 1186–1192. [Google Scholar] [CrossRef]
- Cash, J.L.; Christian, A.R.; Greaves, D.R. Chemerin peptides promote phagocytosis in a ChemR23- and Syk-dependent manner. J. Immunol. 2010, 184, 5315–5324. [Google Scholar] [CrossRef] [Green Version]
- Cash, J.L.; Hart, R.; Russ, A.; Dixon, J.P.; Colledge, W.H.; Doran, J.; Hendrick, A.G.; Carlton, M.B.; Greaves, D.R. Synthetic chemerin-derived peptides suppress inflammation through ChemR23. J. Exp. Med. 2008, 205, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Wittamer, V.; Bondue, B.; Guillabert, A.; Vassart, G.; Parmentier, M.; Communi, D. Neutrophil-mediated maturation of chemerin: A link between innate and adaptive immunity. J. Immunol. 2005, 175, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Lavis, P.; Morra, S.; Orte Cano, C.; Albayrak, N.; Corbière, V.; Olislagers, V.; Dauby, N.; Del Marmol, V.; Marchant, A.; Decaestecker, C.; et al. Chemerin plasma levels are increased in COVID-19 patients and are an independent risk factor of mortality. Front. Immunol. 2022, 13, 941663. [Google Scholar] [CrossRef]
- Ebihara, T.; Matsumoto, H.; Matsubara, T.; Togami, Y.; Nakao, S.; Matsuura, H.; Onishi, S.; Kojima, T.; Sugihara, F.; Okuzaki, D.; et al. Resistin associated with cytokines and endothelial cell adhesion molecules is related to worse outcome in COVID-19. Front. Immunol. 2022, 13, 830061. [Google Scholar] [CrossRef]
- Caterino, M.; Gelzo, M.; Sol, S.; Fedele, R.; Annunziata, A.; Calabrese, C.; Fiorentino, G.; D’Abbraccio, M.; Dell’Isola, C.; Fusco, F.M.; et al. Dysregulation of lipid metabolism and pathological inflammation in patients with COVID-19. Sci. Rep. 2021, 11, 294. [Google Scholar] [CrossRef]
- Kukla, M.; Menżyk, T.; Dembiński, M.; Winiarski, M.; Garlicki, A.; Bociąga-Jasik, M.; Skonieczna, M.; Hudy, D.; Maziarz, B.; Kusnierz-Cabala, B.; et al. Anti-inflammatory adipokines: Chemerin, vaspin, omentin concentrations and SARS-CoV-2 outcomes. Sci. Rep. 2021, 11, 21514. [Google Scholar] [CrossRef]
- Blot, M.; Masson, D.; Nguyen, M.; Bourredjem, A.; Binquet, C.; Piroth, L.; LYMPHONIE Study Group. Are adipokines the missing link between obesity, immune response, and outcomes in severe COVID-19? Int. J. Obes. 2021, 45, 2126–2131. [Google Scholar] [CrossRef]
- Sydor, W.; Wizner, B.; Strach, M.; Bociąga-Jasik, M.; Mydel, K.; Olszanecka, A.; Sanak, M.; Małecki, M.; Wójkowska-Mach, J.; Chrzan, R.; et al. CRACoV-HHS: An interdisciplinary project for multi-specialist hospital and non-hospital care for patients with SARS-CoV-2 infection as well hospital staff assessment for infection exposure. Folia Med. Crac. 2021, 61, 5–44. [Google Scholar] [CrossRef]
- Clinical Spectrum of SARS-CoV-2 Infection. Available online: https://www.covid19treatmentguidelines.nih.gov/overview/clinical-spectrum/ (accessed on 1 August 2022).
- Goralski, K.B.; McCarthy, T.C.; Hanniman, E.A.; Zabel, B.A.; Butcher, E.C.; Parlee, S.D.; Muruganandan, S.; Sinal, C.J. Chemerin, a novel adipokine that regulates adipogenesis and adipocyte metabolism. J. Biol. Chem. 2007, 282, 28175–28188. [Google Scholar] [CrossRef]
- Dong, B.; Ji, W.; Zhang, Y. Elevated serum chemerin levels are associated with the presence of coronary artery disease in patients with metabolic syndrome. Intern. Med. 2011, 50, 1093–1097. [Google Scholar] [CrossRef] [Green Version]
- Weigert, J.; Obermeier, F.; Neumeier, M.; Wanninger, J.; Filarsky, M.; Bauer, S.; Aslanidis, C.; Rogler, G.; Ott, C.; Schäffler, A.; et al. Circulating levels of chemerin and adiponectin are higher in ulcerative colitis and chemerin is elevated in Crohn’s disease. Inflamm. Bowel Dis. 2010, 16, 630–637. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Ponce, F.; Gamez-Nava, J.I.; Perez-Guerrero, E.E.; Saldaña-Cruz, A.M.; Vazquez-Villegas, M.L.; Ponce-Guarneros, J.M.; Huerta, M.; Trujillo, X.; Contreras-Haro, B.; Rocha-Muñoz, A.D.; et al. Serum chemerin levels: A potential biomarker of joint inflammation in women with rheumatoid arthritis. PLoS ONE 2021, 16, e0255854. [Google Scholar] [CrossRef]
- Chougule, D.; Nadkar, M.; Venkataraman, K.; Rajadhyaksha, A.; Hase, N.; Jamale, T.; Kini, S.; Khadilkar, P.; Anand, V.; Madkaikar, M.; et al. Adipokine interactions promote the pathogenesis of systemic lupus erythematosus. Cytokine 2018, 111, 20–27. [Google Scholar] [CrossRef]
- Kukla, M.; Adamek, B.; Waluga, M.; Zalewska-Ziob, M.; Kasperczyk, J.; Gabriel, A.; Mazur, W.; Sobala-Szczygieł, B.; Bułdak, R.J.; Zajęcki, W.; et al. Hepatic chemerin and chemokine-like receptor 1 expression in patients with chronic hepatitis C. Biomed. Res. Int. 2014, 2014, 517820. [Google Scholar] [CrossRef] [Green Version]
- Karampela, I.; Christodoulatos, G.S.; Vallianou, N.; Tsilingiris, D.; Chrysanthopoulou, E.; Skyllas, G.; Antonakos, G.; Marinou, I.; Vogiatzakis, E.; Armaganidis, A.; et al. Circulating Chemerin and Its Kinetics May Be a Useful Diagnostic and Prognostic Biomarker in Critically Ill Patients with Sepsis: A Prospective Study. Biomolecules 2022, 12, 301. [Google Scholar] [CrossRef]
- Horn, P.; Metzing, U.B.; Steidl, R.; Romeike, B.; Rauchfuß, F.; Sponholz, C.; Thomas-Rüddel, D.; Ludewig, K.; Birkenfeld, A.L.; Settmacher, U.; et al. Chemerin in peritoneal sepsis and its associations with glucose metabolism and prognosis: A translational cross-sectional study. Crit. Care 2016, 20, 39. [Google Scholar] [CrossRef] [Green Version]
- Bozaoglu, K.; Curran, J.E.; Stocker, C.J.; Zaibi, M.S.; Segal, D.; Konstantopoulos, N.; Morrison, S.; Carless, M.; Dyer, T.D.; Cole, S.A.; et al. Chemerin, a novel adipokine in the regulation of angiogenesis. J. Clin. Endocrinol. Metab. 2010, 95, 2476–2485. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, N.; Naruse, K.; Kobayashi, Y.; Miyabe, M.; Saiki, T.; Enomoto, A.; Takahashi, M.; Matsubara, T. Chemerin promotes angiogenesis in vivo. Physiol. Rep. 2018, 6, e13962. [Google Scholar] [CrossRef] [Green Version]
- Ben Dhaou, C.; Mandi, K.; Frye, M.; Acheampong, A.; Radi, A.; De Becker, B.; Antoine, M.; Baeyens, N.; Wittamer, V.; Parmentier, M. Chemerin regulates normal angiogenesis and hypoxia-driven neovascularization. Angiogenesis 2022, 25, 159–179. [Google Scholar] [CrossRef]
- Scholz, A.; Plate, K.H.; Reiss, Y. Angiopoietin-2: A multifaceted cytokine that functions in both angiogenesis and inflammation. Ann. N. Y. Acad. Sci. 2015, 1347, 45–51. [Google Scholar] [CrossRef]
- Fiedler, U.; Augustin, H.G. Angiopoietins: A link between angiogenesis and inflammation. Trends Immunol. 2006, 27, 552–558. [Google Scholar] [CrossRef]
- Scharpfenecker, M.; Fiedler, U.; Reiss, Y.; Augustin, H.G. The Tie-2 ligand angiopoietin-2 destabilizes quiescent endothelium through an internal autocrine loop mechanism. J. Cell Sci. 2005, 118, 771–780. [Google Scholar] [CrossRef] [Green Version]
- Smadja, D.M.; Guerin, C.L.; Chocron, R.; Yatim, N.; Boussier, J.; Gendron, N.; Khider, L.; Hadjadj, J.; Goudot, G.; Debuc, B.; et al. Angiopoietin-2 as a marker of endothelial activation is a good predictor factor for intensive care unit admission of COVID-19 patients. Angiogenesis 2020, 23, 611–620. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in COVID-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Perpiñan, C.; Bertran, L.; Terra, X.; Aguilar, C.; Binetti, J.; Lopez-Dupla, M.; Rull, A.; Reverté, L.; Yeregui, E.; Gómez-Bertomeu, F.; et al. On Behalf of COVID-Study Group. Resistin and IL-15 as Predictors of Invasive Mechanical Ventilation in COVID-19 Pneumonia Irrespective of the Presence of Obesity and Metabolic Syndrome. J. Pers. Med. 2022, 12, 391. [Google Scholar] [CrossRef]
- Gonçalves, J.J.; da Mata, C.P.S.M.; Lourenço, A.A.; Ribeiro, Á.L.; Ferreira, G.M.; Fraga-Silva, T.F.C.; de Souza, F.M.; Almeida, V.E.S.; Batista, I.A.D.; Avila-Mesquita, C.; et al. Timeline Kinetics of Systemic and Airway Immune Mediator Storm for Comprehensive Analysis of Disease Outcome in Critically Ill COVID-19 Patients. Front. Immunol. 2022, 13, 903903. [Google Scholar] [CrossRef]
- Kukla, M.; Menżyk, T.; Dembiński, M.; Winiarski, M.; Garlicki, A.; Bociąga-Jasik, M.; Skonieczna, M.; Hudy, D.; Maziarz, B.; Kuśnierz-Cabala, B.; et al. Fetuin-A Deficiency but Not Pentraxin 3, FGF-21, or Irisin, Predisposes to More Serious COVID-19 Course. Biomolecules 2021, 11, 1422. [Google Scholar] [CrossRef]
- Silswal, N.; Singh, A.K.; Aruna, B.; Mukhopadhyay, S.; Ghosh, S.; Ehtesham, N.Z. Human resistin stimulates the pro-inflammatory cytokines TNF-alpha and IL-12 in macrophages by NF-kappaB-dependent pathway. Biochem. Biophys. Res. Commun. 2005, 334, 1092–1101. [Google Scholar] [CrossRef]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mol. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef]
- van Zelst, C.M.; Janssen, M.L.; Pouw, N.; Birnie, E.; Castro Cabezas, M.; Braunstahl, G.J. Analyses of abdominal adiposity and metabolic syndrome as risk factors for respiratory distress in COVID-19. BMJ Open Respir. Res. 2020, e000792. [Google Scholar] [CrossRef]
- Kearns, S.M.; Ahern, K.W.; Patrie, J.T.; Horton, W.B.; Harris, T.E.; Kadl, A. Reduced adiponectin levels in patients with COVID-19 acute respiratory failure: A case-control study. Physiol. Rep. 2021, 9, e14843. [Google Scholar] [CrossRef]
- Fogacci, F.; Borghi, C.; Cicero, A.F.G. Misinterpreting data in lipidology in the era of COVID-19. J. Clin. Lipidol. 2020, 14, 543–544. [Google Scholar] [CrossRef]
- Ray, C.A.; Bowsher, R.R.; Smith, W.C.; Devanarayan, V.; Willey, M.B.; Brandt, J.T.; Dean, R.A. Development, validation, and implementation of a multiplex immunoassay for the simultaneous determination of five cytokines in human serum. J. Pharm. Biomed. Anal. 2005, 36, 1037–1044. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Severe COVID-19 | Severe COVID-19 | p-Value | |
|---|---|---|---|
| Age, years | 61 (51–69) | 62 (50–68) | 0.590 |
| Male sex, % | 47 | 53 | 0.400 |
| Body mass index, kg/m² | 28.2 (4.8) | 30.6 (5.4) | <0.001 |
| Waist circumference, cm | 98.4 (14.8) | 107.1 (17.3) | <0.001 |
| Comorbidities | |||
| Diabetes, % | 14 | 26 | 0.037 |
| Hypertension, % | 53 | 57 | 0.516 |
| COPD, % | 6 | 3 | 0.584 |
| Liver diseases, % | 4 | 5 | 0.439 |
| Malignancy, % | 4 | 2 | 0.610 |
| Laboratory data | |||
| WBC, 10³/µL | 5.39 (4.1–6.94) | 5.39 (4.3–8.07) | 0.313 |
| Neutrophiles, 10³/µL | 4.0 (1.9) | 4.9 (2.9) | 0.012 |
| Lymphocytes, 10³/µL | 0.98 (0.47) | 0.84 (0.4) | 0.031 |
| Monocytes, 10³/µL | 0.40 (0.2) | 0.33 (0.19) | 0.025 |
| RBC, 10⁶/µL | 4.58 (0.55) | 4.61 (0.45) | 0.740 |
| Hemoglobin, g/dL | 13.5 (1.5) | 13.7 (1.3) | 0.290 |
| Hematocrit, % | 39.7 (4.3) | 40.0 (3.7) | 0.629 |
| Platelets, 10³/µL | 207 (81) | 215 (77) | 0.522 |
| C-reactive protein, mg/L | 68.95 (32.2–128) | 91 (47.4–158) | <0.001 |
| IL-6, pg/mL | 30.7 (12.3–61) | 32.5 (11.1–62.1) | 0.836 |
| Procalcitonin, ng/mL | 0.09 (0.05–0.18) | 0.11 (0.06–0.22) | <0.001 |
| Ferritin, µg/L | 689 (419–1162] | 878 (540–1486) | <0.001 |
| D-dimer, mg/L | 0.7 (0.5–1.3) | 0.77 (0.54–1.17) | 0.535 |
| LDH, U/I | 348 (278–419) | 378 (311–501) | <0.001 |
| ALT, U/I | 37 (26–56) | 42.5 (27–61) | 0.033 |
| AST, U/I | 48 (35–66) | 53 (40–74) | 0.003 |
| GGT, U/I | 47 (28–87) | 58 (37–107) | 0.001 |
| Alkaline phosphatase, U/I | 59 (48–77) | 62 (48–80) | 0.568 |
| Bilirubin, µmol/L | 7.1 (3.2) | 7.0 (3.0) | 0.696 |
| Glucose, mmol/L | 6.56 (5.28–8.26) | 7.16 (6.24–9.09) | <0.001 |
| HbA1c, % | 5.9 (5.6–6.3) | 6.0 (5.7–6.5) | 0.012 |
| Albumin, g/L | 38.5 (3.8) | 36.2 (3.2) | <0.001 |
| Creatinine, µmol/L | 75 (63.3–94.4) | 77.9 (63.8–98) | 0.322 |
| eGFR, mL/min/L,73 m² | 89 (67–90) | 87.5 (67–90) | 0.623 |
| Non-Severe COVID-19 | Severe COVID-19 | p-Value | |
|---|---|---|---|
| Day 1 | |||
| TNFα, pg/mL | 19.2 (16–24.7) | 23.6 (19.7–29) | <0.001 |
| Angiopoietin2, pg/mL | 1548 (1108–2268) | 1477 (1043–2213) | 0.577 |
| IL-1β, pg/mL | 17.3 (10.8–22.8) | 24.7 (16–29.1) | <0.001 |
| Pentraxin, pg/mL | 5213 (27,430–8301) | 8781 (5091–12,744) | <0.001 |
| Resistin, pg/mL | 17,961 (13,593–28,441) | 23,098 (15,761–31,369) | 0.032 |
| Chemerin, pg/mL | 8774 (3228) | 9183 (4277) | 0.396 |
| Adiponectin, ng/mL | 8536 (5374–54,126) | 5384 (4361–8387) | <0.001 |
| Leptin, pg/mL | 1578 (885–3546) | 2343 (1092–4235) | 0.091 |
| Day 7 | |||
| TNFα, pg/mL | 21.3 (17.3–26.2) | 25.5 (21.5–28.9) | <0.001 |
| Angiopoietin2, pg/mL | 1246 (899–1764) | 1260 (911–1598) | 0.547 |
| IL-1β, pg/mL | 17.9 (14.1–25.2) | 26.1 (20.4–30.2) | <0.001 |
| Pentraxin3, pg/mL | 3080 (1928–4890) | 3443 (2563–5157) | 0.095 |
| Resistin, pg/mL | 16,635 (7866) | 17,984 (9060) | 0.212 |
| Chemerin, pg/mL | 7256 (5607–9511) | 7394 (5747–9603) | 0.565 |
| Adiponectin, ng/mL | 7977 (5714–47,591) | 6308 (4839–9285) | <0.001 |
| Leptin, pg/mL | 1461 (745–2988) | 1702 (981–3438) | 0.101 |
| Day 28 | |||
| TNFα, pg/mL | 18.7 (15.2–22.2) | 20.5 (17.5–24) | 0.002 |
| Angiopoietin2, pg/mL | 1662 (1156–2105) | 1662 (1174–2134) | 0.877 |
| IL-1β, pg/mL | 16 (10.8–21.2) | 2..4 (14.9–26.8) | <0.001 |
| Pentraxin3, pg/mL | 1707 (1044–2353) | 1954 (1191–2785) | 0.130 |
| Resistin, pg/mL | 11,609 (8632–15,307) | 11,740 (8339–15,999) | 0.951 |
| Chemerin, pg/mL | 9171 (7345–10,594) | 9618 (7490–11,310) | 0.252 |
| Adiponectin, ng/mL | 11,693 (6698–62,374) | 7283 (5674–10,450) | <0.001 |
| Leptin, pg/mL | 1263 (755–2168) | 1452 (847–3006) | 0.194 |
| Day 1 | Day 7 | Day 28 | p-Value | |
|---|---|---|---|---|
| TNFα, pg/mL | 21.9 (17.2–26.6) | 24.2 (19.2–27.6) | 19.5 (16.6–23.1) | p < 0.001 * |
| IL-1β, pg/mL | 21.2 (12.5–26.9) | 23.6 (15–28.4) | 18.3 (12.5–24.7) | p < 0.001 # |
| Pentraxin 3, pg/mL | 6643 (3852–11,148) | 3369 (2179–5007) | 1839 (1111–2597) | p < 0.001 * |
| Chemerin, pg/mL | 8916 (6661–10,773) | 7359 (5705–9533) | 9324 (7371–11,033) | p < 0.001 # |
| Adiponectin, ng/mL | 6559 (4640–13,041) | 7134 (5170–14,854) | 8455 (6080–19,191) | p < 0.001 * |
| Resistin, pg/mL | 21,600 (14,123–29,587) | 5586 (11,257–21,676) | 11,629 (8415–15,690) | p < 0.001 * |
| Angiopoietin2, pg/mL | 1502 (1070–2247) | 1256 (907–1678) | 1662 (1173–2125) | p < 0.001 # |
| Leptin, pg/mL | 1824 (989–3981) | 1569 (858–3153) | 1372 (820–2652) | p < 0.001 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulicka-Grodzicka, J.; Surdacki, A.; Surmiak, M.; Sanak, M.; Wizner, B.; Sydor, W.; Bociąga-Jasik, M.; Strach, M.; Korkosz, M.; Skladany, L.; et al. Chemerin as a Potential Marker of Resolution of Inflammation in COVID-19 Infection. Biomedicines 2022, 10, 2462. https://doi.org/10.3390/biomedicines10102462
Sulicka-Grodzicka J, Surdacki A, Surmiak M, Sanak M, Wizner B, Sydor W, Bociąga-Jasik M, Strach M, Korkosz M, Skladany L, et al. Chemerin as a Potential Marker of Resolution of Inflammation in COVID-19 Infection. Biomedicines. 2022; 10(10):2462. https://doi.org/10.3390/biomedicines10102462
Chicago/Turabian StyleSulicka-Grodzicka, Joanna, Andrzej Surdacki, Marcin Surmiak, Marek Sanak, Barbara Wizner, Wojciech Sydor, Monika Bociąga-Jasik, Magdalena Strach, Mariusz Korkosz, Lubomir Skladany, and et al. 2022. "Chemerin as a Potential Marker of Resolution of Inflammation in COVID-19 Infection" Biomedicines 10, no. 10: 2462. https://doi.org/10.3390/biomedicines10102462