
Vascular Effects of Low-Dose ACE2 Inhibitor MLN-4760—Benefit or Detriment in Essential Hypertension?

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Guide for the Use and Care of Laboratory Animals
2.2. Experimental Model and Blood Pressure Determination
2.3. In Vivo Study—Integrated Blood Pressure Response
2.4. In Vitro Studies
2.4.1. Preparation of Blood Samples for Determination of Biochemical Parameters in Plasma
2.4.2. Analysis of Angiotensins
2.4.3. Measurement of H2S Concentration
2.4.4. Vasoactive Responses of Thoracic Aorta
2.4.5. Vasoactive Responses of Femoral and Mesenteric Arteries
2.4.6. Total NO Synthase Activity
2.4.7. RNA Isolation and Real-Time PCR
2.4.8. Western Blotting
2.4.9. Measurement of ACE2 Activity
2.5. Evaluation of Angiogenesis in the Model of Chick Chorioallantoic Membrane
2.6. Statistical Analysis
2.7. Drugs
3. Results
3.1. General Characteristics of Experimental Animals
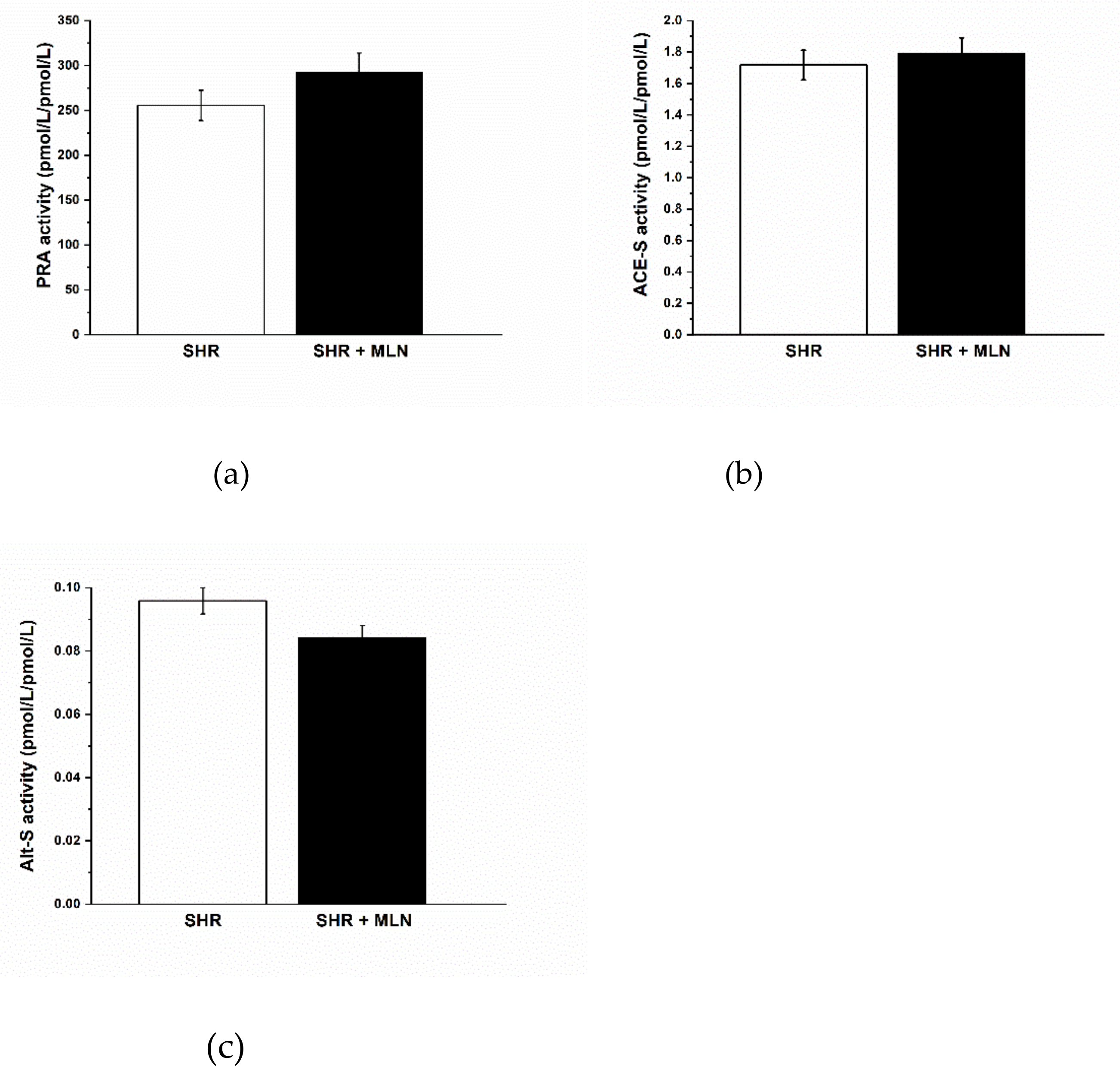
3.2. Analysis of Angiotensins in Plasma
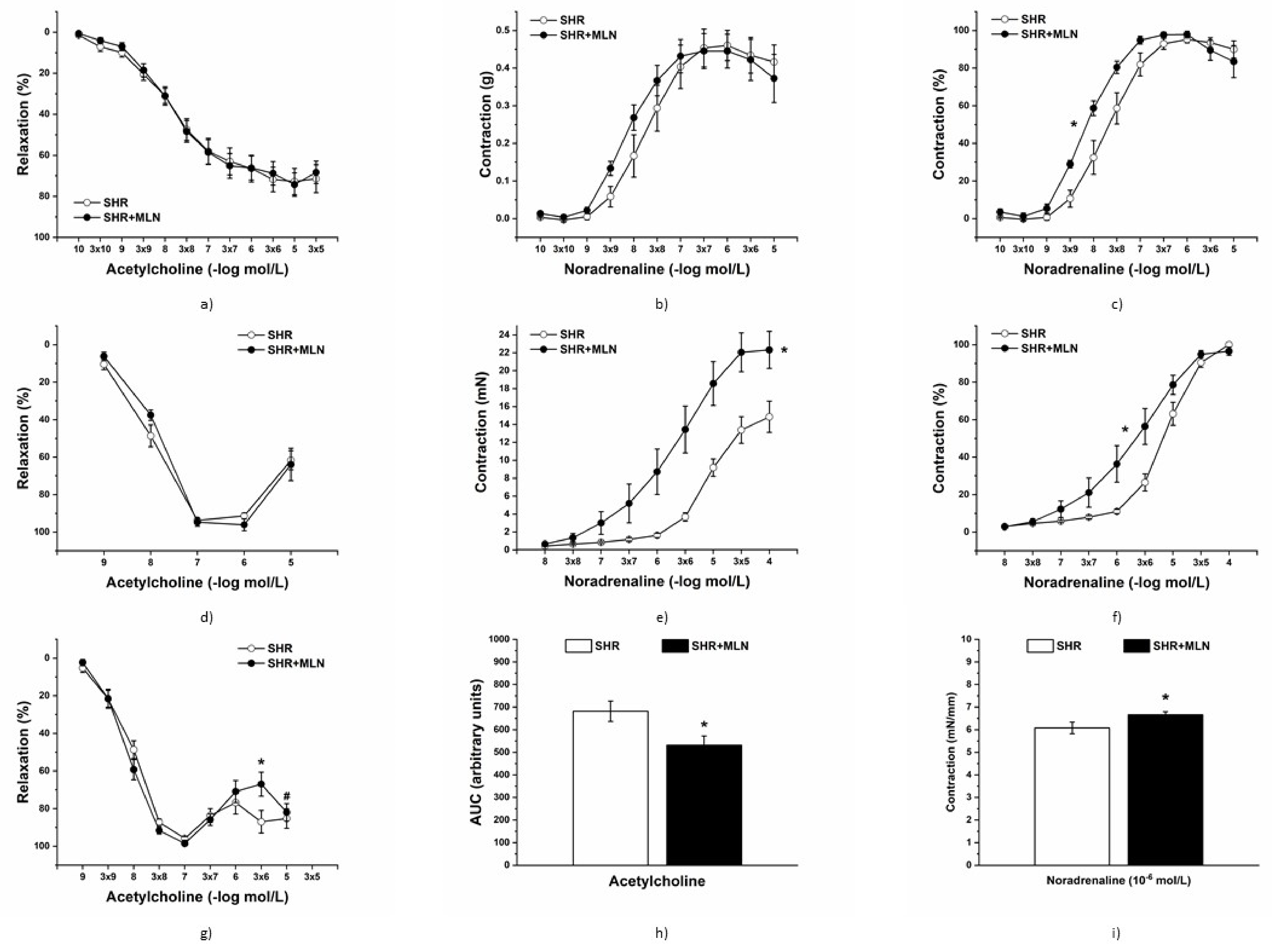
3.3. In Vivo Vasoactive Responses
3.4. Endothelial Function and Contractile Properties of Isolated Arteries
3.4.1. Analyses of Mas Receptor Pathway
3.4.2. Analyses of NO Pathway
3.4.3. Analyses of H2S Pathway
3.5. Effect of MLN-4760 on Angiogenesis in Chick CAM Model
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, Y.Y.; Ma, Y.T.; Zhang, J.Y.; Xie, X. COVID-19 and the cardiovascular system. Nat. Rev. Cardiol. 2020, 17, 259–260. [Google Scholar] [CrossRef] [Green Version]
- Zemlin, A.E.; Wise, O.J. Coronavirus disease 2019 (COVID-19) and the renin-angiotensin system: A closer look at angiotensin-converting enzyme 2 (ACE2). Ann. Clin. Biochem. 2020, 57, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 180, 281–292. [Google Scholar] [CrossRef]
- Dobrocsyova, V.; Krskova, K.; Capcarova, M.; Zorad, S. Modulation of Adipogenesis and Oxidative Status by Quercetin and Ochratoxin A: Positive or Negative Impact on Rat Adipocyte Metabolism? Molecules 2019, 24, 3726. [Google Scholar] [CrossRef] [Green Version]
- South, A.M.; Diz, D.I.; Chappell, M.C. COVID-19, ACE2, and the cardiovascular consequences. Am. J. Physiol. 2020, 318, H1084–H1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovren, F.; Pan, Y.; Quan, A.; Teoh, H.; Wang, G.; Shukla, P.C.; Levitt, K.S.; Oudit, G.Y.; Al-Omran, M.; Stewart, D.J.; et al. Angiotensin converting enzyme-2 confers endothelial protection and attenuates atherosclerosis. Am. J. Physiol. 2008, 295, H1377–H1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousif, M.H.M.; Dhaunsi, G.S.; Makki, B.M.; Qabazard, B.A.; Akhtar, S.; Benter, I.F. Characterization of Angiotensin-(1-7) effects on the cardiovascular system in an experimental model of type-1 diabetes. Pharmacol. Res. 2012, 66, 269–275. [Google Scholar] [CrossRef]
- Zicha, J.; Kunes, J. Ontogenetic aspects of hypertension development: Analysis in the rat. Physiol. Rev. 1999, 79, 1227–1282. [Google Scholar] [CrossRef]
- Cebova, M.; Kristek, F. Age-dependent ultrastructural changes of coronary artery in spontaneously hypertensive rats. Gen. Physiol. Biophys. 2011, 30, 364–372. [Google Scholar] [CrossRef]
- Puzserova, A.; Ilovska, V.; Balis, P.; Slezak, P.; Bernatova, I. Age-related alterations in endothelial function of femoral artery in young SHR and WKY rats. Biomed. Res. Int. 2014, 2014, 658479. [Google Scholar] [CrossRef] [Green Version]
- Berenyiova, A.; Drobna, M.; Cebova, M.; Kristek, F.; Cacanyiova, S. Changes in the vasoactive effects of nitric oxide, hydrogen sulfide and the structure of the rat thoracic aorta: The role of age and essential hypertension. J. Physiol. Pharmacol. 2018, 69, 4. [Google Scholar] [CrossRef]
- Kodavanti, U.P.; Schladweiler, M.C.; Ledbetter, A.D.; Watkinson, W.P.; Campen, M.J.; Winsett, D.W.; Richards, J.R.; Crissman, K.M.; Hatch, G.E.; Costa, D.L. The spontaneously hypertensive rat as a model of human cardiovascular disease: Evidence of exacerbated cardiopulmonary injury and oxidative stress from inhaled emission particulate matter. Toxicol. App. Pharmacol. 2000, 164, 250–263. [Google Scholar] [CrossRef]
- Wang, Y.; Fu, W.; Xue, Y.; Lu, Z.; Li, Y.; Yu, P.; Yu, X.; Xu, H.; Sui, D. Ginsenoside Rcameliorates endothelial insulin resistance via upregulation of angiotensin-converting enzyme 2. Front. Pharmacol. 2021, 12, 620524. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zeng, Z.; Cao, Y.; Liu, Y.; Ping, F.; Liang, M.; Xue, Y.; Xi, C.; Zhou, M.; Jiang, W. Angiotensin-converting enzyme 2 prevents lipopolysaccharide-induced rat acute lung injury via suppressing the ERK1/2 and NF-κB signaling pathways. Sci. Rep. 2016, 6, 27911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernatova, I. Endothelial dysfunction in experimental models of arterial hypertension: Cause or consequence? Biomed. Res. Int. 2014. [Google Scholar] [CrossRef]
- Cacanyiova, S. The vasoactive role of nitric oxide: Physiological and morphological aspects. Curr. Pharm. Biotechnol. 2011, 12, 1294–1304. [Google Scholar] [CrossRef]
- Jiang, T.; Yu, J.T.; Zhu, X.C.; Zhang, Q.Q.; Tan, M.S.; Cao, L.; Wang, H.F.; Lu, J.; Gao, Q.; Zhang, Y.D.; et al. Angiotensin-(1-7) induces cerebral ischaemic tolerance by promoting brain angiogenesis in a Mas/eNOS-dependent pathway. Br. J. Pharmacol 2014, 171, 4222–4232. [Google Scholar] [CrossRef]
- Sampaio, W.O.; Souza dos Santos, R.A.; Faria-Silva, R.; Tapias da Mata Machado, R.; Schiffrin, E.L.; Touyz, R.M. Angiotensin-(1-7) through receptor Mas mediates endothelial nitric oxide synthase activation via Akt-dependent pathways. Hypertension 2007, 49, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Raffai, G.; Lombard, J.H. Angiotensin-(1-7) Selectively induces relaxation and modulates endothelium-dependent dilation in mesenteric arteries of salt-fed rats. J. Vasc. Res. 2016, 53, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Savoia, C.; Arrabito, E.; Parente, R.; Nicoletti, C.; Madaro, L.; Battistoni, A.; Filippini, A.; Steckelings, U.M.; Touyz, R.M.; Volpe, M. Mas receptor activation contributes to the improvement of nitric oxide bioavailability and vascular remodeling during chronic AT1R (Angiotensin Type-1 Receptor) blockade in experimental hypertension. Hypertension 2020, 76, 1753–1761. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.P.; Losordo, D.W. Nitric Oxide and Angiogenesis. Circulation 2002, 105, 2133–2135. [Google Scholar] [CrossRef]
- Matsunaga, T.; Weihrauch, D.W.; Moniz, M.C.; Tessmer, J.; Warltier, D.C.; Chilian, W.M. Angiostatin inhibits coronary angiogenesis during impaired production of nitric oxide. Circulation 2002, 105, 2185–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haendeler, J.; Dimmeler, S. Regulation of Angiogenesis by Angiotensin II. In Angiotensin Volume I. Handbook of Experimental Pharmacology; Unger, T., Schölkens, B.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; Volume 163, pp. 99–109. [Google Scholar] [CrossRef]
- Lin, Y.; Zeng, H.; Gao, L.; Gu, T.; Wang, C.; Zhang, H. Hydrogen sulfide attenuates atherosclerosis in a partially ligated carotid artery mouse model via regulating angiotensin converting enzyme 2 expression. Front. Physiol. 2017, 8, 782. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Hsu, C.N.; Lu, P.C. Early short-term treatment with exogenous hydrogen sulfide postpones the transition from prehypertension to hypertension in spontaneously hypertensive rat. Clin. Exp. Hypertens. 2018, 40, 58–64. [Google Scholar] [CrossRef]
- Cacanyiova, M.; Majzunova, M.; Golas, S.; Berenyiova, A. The role of perivascular adipose tissue and endogenous hydrogen sulfide in vasoactive responses of isolated mesenteric arteries in normotensive and spontaneously hypertensive rats. J. Physiol. Pharmacol. 2019, 70, 295–306. [Google Scholar] [CrossRef]
- Guo, Z.; Poglitsch, M.; McWhinney, B.C.; Ungerer, J.P.J.; Ahmed, A.H.; Gordon, R.D.; Wolley, M.; Stowasser, M. Measurement of Equilibrium Angiotensin II in the Diagnosis of Primary Aldosteronism. Clin. Chem. 2020, 66, 483–492. [Google Scholar] [CrossRef]
- Dayar, E.; Kara, E.; Yetik-Anacak, G.; Hocaoglu, N.; Bozkurt, O.; Gidener, S.; Durmus, N. Do penile haemodynamics change in the presence of hydrogen sulphide (H2S) donor in metabolic syndrome-induced erectile dysfunc-tion? Andrologia 2018, 50, e12885. [Google Scholar] [CrossRef] [PubMed]
- Mok, Y.Y.; Atan, M.S.; Ping, C.Y.; Zhong, W.J.; Bhatia, M.; Moochhala, S.; Moore, P.K. Role of hydrogen sulphide in haemorrhagic shock in the rat: Protective effect of inhibitors of hydrogen sulphide biosynthesis. Br. J. Pharmacol. 2004, 143, 881–889. [Google Scholar] [CrossRef]
- Yetik-Anaca, K.G.; Dereli, M.V.; Sevin, G.; Ozzayım, O.; Erac, Y.; Ahmed, A. Resveratrol Stimulates Hydrogen Sulfide (H2S) Formation to Relax Murine Corpus Cavernosum. J. Sex Med. 2015, 12, 2004–2012. [Google Scholar] [CrossRef]
- Cacanyiova, S.; Berenyiova, A.; Malekova, M.; Kristek, F.; Dovinova, I.; Krenek, P.; Pivackova, L.; Pifkova, I. Different vasoactive effects of chronic endothelial and neuronal NO-synthase inhibition in young Wistar rats. J. Physiol. Biochem. 2014, 70, 749–760. [Google Scholar] [CrossRef]
- Cacanyiova, S.; Berenyiova, A. In Vitro Measurement of H2S-Mediated Vasoactive Responses. Methods Mol. Biol. 2019, 2007, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Balis, P.; Berenyiova, A.; Radosinska, J.; Kvandova, M.; Bernatova, I.; Puzserova, A. High concentration of uric acid failed to affect endothelial function of small mesenteric arteries, femoral arteries and aortas from aged Wistar-Kyoto rats. J. Physiol. Pharmacol. 2020, 71, 399–408. [Google Scholar] [CrossRef]
- Liskova, S.; Balis, P.; Micurova, A.; Kluknavsky, M.; Okuliarova, M.; Puzserova, A.; Skratek, M.; Sekaj, I.; Manka, J.; Valovic, P.; et al. Effect of iron oxide nanoparticles on vascular function and nitric oxide production in acute stress-exposed rats. Physiol. Res. 2020, 69, 1067–1083. [Google Scholar] [CrossRef]
- Liskova, S.; Petrova, M.; Karen, P.; Behuliak, M.; Zicha, J. Contribution of Ca2+-Dependent Cl− Channels to Norepinephrine-Induced Contraction of Femoral Artery Is Replaced by Increasing EDCF Contribution during Ageing. BioMed Res. Int. 2014, 2014, 289361. [Google Scholar] [CrossRef]
- Pechanova, O.; Bernatova, I.; Pelouch, V.; Simko, F. Protein remodelling of the heart in NO-deficient hypertension: The effect of captopril. J. Mol. Cell Cardiol. 1997, 29, 3365–3374. [Google Scholar] [CrossRef] [PubMed]
- Merckx, G.; Tay, H.; Monaco, M.L.; Zandvoort, M.; Spiegelaere, W.; Lambrichts, I.; Bronckaers, A. Chorioallantoic Membrane Assay as Model for Angiogenesisin Tissue Engineering: Focus on Stem Cells. Tissue Eng. Part. B 2020, 26, 519–539. [Google Scholar] [CrossRef]
- Mangir, N.; Dikici, S.; Claeyssens, F.; MacNei, S. Using ex Ovo Chick Chorioallantoic Membrane (CAM) Assay to Evaluate the Biocompatibility and Angiogenic Response to Biomaterials. ACS Biomater. Sci. Eng. 2019, 5, 3190–3200. [Google Scholar] [CrossRef]
- Bruce, E.B.; Sakarya, Y.; Kirichenko, N.; Toklu, H.Z.; Sumners, C.; Morgan, D.; Tümer, N.; Scarpace, P.J.; Carter, C.S. ACE2 activatordiminazeneaceturatereducesadipositybutpreservesleanmass in young and old rats. Exp. Gerontol. 2018, 111, 133–140. [Google Scholar] [CrossRef]
- Chappell, M.C. The Angiotensin-(1-7) Axis: Formation and Metabolism Pathways. In Angiotensin; Santos, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar] [CrossRef]
- Zemancikova, A.; Torok, J.; Balis, P.; Valovic, P.; Ulicna, O.; Chomova, M. Modulation of sympathoadrenergic contractions by perivascular adipose tissue in mesenteric arteries of rats with different level of body adiposity. J. Physiol. Pharmacol. 2020, 71, 589–596. [Google Scholar] [CrossRef]
- Félétou, M.; Vanhoutte, P.M. Endothelium-dependent hyperpolarizations: Past beliefs and present facts. Ann. Med. 2007, 39, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Gerova, M.; Kittova, M. Systemic blood pressure response to the inhibition of two hyperpolarizing pathways: A comparison to NO-synthase inhibition. Physiol. Res. 2006, 55, 603–610. [Google Scholar]
- Bernatova, I.; Balis, P.; Goga, R.; Behuliak, M.; Zicha, J.; Sekaj, I. Lack of reactive oxygen species deteriorates blood pressure regulation in acute stress. Physiol. Res. 2016, 65, S381–S390. [Google Scholar] [CrossRef] [PubMed]
- Hamid, S.; Rhaleb, I.A.; Kassem, K.M.; Rhaleb, N.E. Role of Kinins in Hypertension and Heart Failure. Pharmaceuticals 2020, 13, 347. [Google Scholar] [CrossRef] [PubMed]
- Laggner, H.; Hermann, M.; Esterbauer, H.; Muellner, M.K.; Exner, M.; Gmeiner, B.M.; Kapiotis, S. The novel gaseous vasorelaxant hydrogen sulfide inhibits angiotensin-converting enzyme activity of endothelial cells. J. Hypertens. 2007, 10, 2100–2104. [Google Scholar] [CrossRef] [PubMed]
- Drobna, M.; Misak, A.; Holland, T.; Kristek, F.; Grman, M.; Tomasova, L.; Berenyiova, A.; Cacanyiova, S.; Ondrias, K. Captopril partially decreases the effect of H(2)S on rat blood pressure and inhibits H(2)S-induced nitric oxide release from S-nitrosoglutathione. Physiol. Res. 2015, 64, 479–486. [Google Scholar] [CrossRef]
- Cacanyiova, S.; Berenyiova, A.; Kristek, F.; Drobna, M.; Ondrias, K.; Grman, M. The adaptive role of nitric oxide and hydrogen sulphide in vasoactive responses of thoracic aorta is triggered already in young spontaneously hypertensive rats. J. Physiol. Pharmacol. 2016, 67, 501–512. [Google Scholar]
- Wei, X.; Lan, T.; Zhou, Y.; Cheng, J.; Li, P.; Zeng, X.; Yang, Y. Mechanism of α1-Adrenergic Receptor-Induced Increased Contraction of Rat Mesenteric Artery in Aging Hypertension Rats. Gerontology 2021, 67, 323–337. [Google Scholar] [CrossRef]
- Lemos, V.S.; Côrtes, S.F.; Silva, D.M. Campagnole-Santos MJ, Santos RA. Angiotensin-(1-7) is involved in the endothelium-dependent modulation of phenylephrine-induced contraction in the aorta of mRen-2 transgenic rats. Br. J. Pharmacol. 2002, 135, 1743–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Yi-Ming, W.; Li, Z.Z.; Zhao, L.; Yu, Y.S.; Li, D.J.; Xia, C.Y.; Liu, J.G.; Su, D.F. Local RAS and inflammatory factors are involved in cardiovascular hypertrophy in spontaneously hypertensive rats. Pharmacol. Res. 2008, 58, 196–201. [Google Scholar] [CrossRef]
- Balis, P.; Puzserova, A.; Slezak, P.; Sestakova, N.; Pechanova, O.; Bernatova, I. Short-term administration of alibernet red wine extract failed to affect blood pressure and to improve endothelial function in young normotensive and spontaneously hypertensive rats. Physiol. Res. 2013, 62, 631–641. [Google Scholar] [CrossRef]
- Sotomayor, A.M.; Bueno, R.; Pérez-Guerrero, C.; Herrera, M.D. Effect of L-carnitine and propionyl-L-carnitine on endothelial function of small mesenteric arteries from SHR. J. Vasc. Res. 2007, 44, 354–364. [Google Scholar] [CrossRef]
- Silva, D.M.; Gomes-Filho, A.; Olivon, V.C.; Santos, T.M.; Becker, L.K.; Santos, R.A.; Lemos, V.S. Swimming training improves the vasodilator effect of angiotensin-(1-7) in the aorta of spontaneously hypertensive rat. J. Appl. Physiol. 2011, 111, 1272–1277. [Google Scholar] [CrossRef]
- Zhang, F.; Tang, H.; Sun, S.; Luo, Y.; Ren, X.; Chen, A.; Li, P.; Han, Y. Angiotensin-(1-7) induced vascular relaxation in spontaneously hypertensive rats. Nitric Oxide 2019, 88, 1–9. [Google Scholar] [CrossRef]
- Zhao, Y.; Vanhouttep, M.; Leung, S.W. Endothelial nitric oxide synthase-independent release of nitric oxide in the aorta of the spontaneously hypertensive rat. J. Pharmacol. Exp. Ther. 2013, 344, 15–22. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, X.; Patel, K.P. Angiotensin-converting enzyme 2 overexpression improves central nitric oxide-mediated sympathetic outflow in chronic heart failure. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H2402–H2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, P.Y.; Chen, J.J.; Yen, M.H. The expression of heme oxygenase-1 and inducible nitric oxide synthase in aorta during the development of hypertension in spontaneously hypertensive rats. Am. J. Hypertens. 2004, 17, 1127–1134. [Google Scholar] [CrossRef]
- Pechanova, O.; Bernatova, I.; Babal, P.; Martínez, M.C.; Kysela, S.; Stvrtina, S.; Andriantsitohaina, R. Red wine polyphenols prevent cardiovascular alterations in L-NAME-induced hypertension. J. Hypertens. 2004, 22, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Kristek, F.; Faberova, V.; Varga, I. Long-term effect of molsidomine and pentaerythrityl tetranitrate on cardiovascular system of spontaneously hypertensive rats. Physiol. Res. 2003, 52, 709–717. [Google Scholar] [PubMed]
- Dovinova, I.; Cacanyiova, S.; Faberova, V.; Kristek, F. The effect of an NO donor, pentaerythrityl tetranitrate, on biochemical, functional, and morphological attributes of cardiovascular system of spontaneously hypertensive rats. Gen. Physio. Biophys. 2009, 28, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Berenyiova, A.; Golas, S.; Drobna, M.; Cebova, M.; Cacanyiova, S. Fructose Intake Impairs the Synergistic Vasomotor Manifestation of Nitric Oxide and Hydrogen Sulfide in Rat Aorta. Int J. Mol. Sci. 2021, 22, 4749. [Google Scholar] [CrossRef]
- Cacanyiova, S.; Golas, S.; Zemancikova, A.; Majzunova, M.; Cebova, M.; Malinska, H.; Hüttl, M.; Markova, I.; Berenyiova, A. The Vasoactive Role of Perivascular Adipose Tissue and the Sulfide Signaling Pathway in a Nonobese Model of Metabolic Syndrome. Biomolecules 2021, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Nami, B.; Ghanaeian, A.; Ghanaeian, K.; Houri, R.; Nami, N.; Ghasemi-Dizgah, A.; Caluseriu, N. The interaction of the severe acute respiratory syndrome coronavirus 2 spike protein with drug-inhibited angiotensin converting enzyme 2 studied by molecular dynamics simulation. J. Hypertens 2021, 39, 1705–1716. [Google Scholar] [CrossRef] [PubMed]
- Angeli, F.; Reboldi, G.; Verdecchia, P. SARS-CoV-2 infection and ACE2 inhibition. J. Hypertens. 2021, 39, 1555–1558. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | SHR | n | SHR + MLN | n |
|---|---|---|---|---|
| basal BW (g) | 292.00 ± 3.69 | 20 | 296.20 ± 3.84 | 20 |
| end BW (g) | 309.65 ± 4.09 | 20 | 321.05 ± 4.42 | 20 |
| Δ BW (%) | 6.05 ± 0.44 | 20 | 8.38 ± 0.45 *** | 20 |
| basal SBP (mmHg) | 160.78 ± 5.76 | 18 | 162.61 ± 5.53 | 18 |
| end SBP (mm/hg) | 170.06 ± 6.58 | 18 | 174.17 ± 7.21 | 18 |
| Δ SBP (%) | 5.87 ± 2.31 | 18 | 7.16 ± 2.83 | 18 |
| HW (mg) | 1148.42 ± 0.04 | 11 | 1209.49 ± 0.03 | 12 |
| HW/BW (mg/g) | 3.72 ± 0.14 | 11 | 3.78 ± 0.10 | 12 |
| VF (mg) | 7833.22 ± 331.9 | 12 | 8985.02 ± 300.45 * | 12 |
| VF/BW (mg/g) | 25.1 ± 0.86 | 12 | 28.1 ± 0.92 * | 12 |
| ACE2 activity (mU/mg) | 14.61 ± 3.54 | 12 | 19.30 ± 1.39 | 12 |
| H2S plasma (µM) | 11.9 ± 0.7 | 12 | 22.7 ± 2.4 *** | 12 |
| H2S heart (nmol/mg of protein) GLU (mmol/L) | 1062 ± 61 6.97 ± 0.05 | 12 8 | 1534 ± 91 *** 7.13 ± 0.24 | 12 8 |
| TG (mmol/L) | 1.66 ± 0.09 | 8 | 1.70 ± 0.13 | 8 |
| CHOL (mmol/(L) | 2.10 ± 0.05 | 8 | 2.21 ± 0.06 | 8 |
| HDL-C (mmol/L) | 1.34 ± 0.04 | 8 | 1.41 ± 0.03 | 8 |
| Parameter | SHR | n | SHR + MLN | n |
|---|---|---|---|---|
| Ang I (1–10) (pmol/L) | 94.53 ± 6.44 | 7 | 105.93 ± 9.67 | 7 |
| Ang II (1–8) (pmol/L) | 161.05 ± 11.94 | 7 | 186.42 ± 13.13 | 7 |
| Ang 1–7 (pmol/L) | 6.61 ± 0.98 | 7 | 6.36 ± 0.89 | 7 |
| Ang 1–5 (pmol/L) | 20.27 ± 1.25 | 7 | 20.89 ± 2.09 | 7 |
| Ang IV (3–8) (pmol/L) | 8.09 ± 0.73 | 7 | 10.16 ± 0.85 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berenyiova, A.; Bernatova, I.; Zemancikova, A.; Drobna, M.; Cebova, M.; Golas, S.; Balis, P.; Liskova, S.; Valaskova, Z.; Krskova, K.; et al. Vascular Effects of Low-Dose ACE2 Inhibitor MLN-4760—Benefit or Detriment in Essential Hypertension? Biomedicines 2022, 10, 38. https://doi.org/10.3390/biomedicines10010038
Berenyiova A, Bernatova I, Zemancikova A, Drobna M, Cebova M, Golas S, Balis P, Liskova S, Valaskova Z, Krskova K, et al. Vascular Effects of Low-Dose ACE2 Inhibitor MLN-4760—Benefit or Detriment in Essential Hypertension? Biomedicines. 2022; 10(1):38. https://doi.org/10.3390/biomedicines10010038
Chicago/Turabian StyleBerenyiova, Andrea, Iveta Bernatova, Anna Zemancikova, Magdalena Drobna, Martina Cebova, Samuel Golas, Peter Balis, Silvia Liskova, Zuzana Valaskova, Katarina Krskova, and et al. 2022. "Vascular Effects of Low-Dose ACE2 Inhibitor MLN-4760—Benefit or Detriment in Essential Hypertension?" Biomedicines 10, no. 1: 38. https://doi.org/10.3390/biomedicines10010038