
Ribosome Biogenesis Serves as a Therapeutic Target for Treating Endometriosis and the Associated Complications

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse and Reagents
2.2. Mouse Model of Induced Endometriosis
2.3. Immune Cell Profiling by Flow Cytometry Analysis
2.4. Behavior Assessments
2.5. Sandwich ELISA
2.6. Real-Time qPCR
2.7. Immunohistochemistry (IHC) and Immunofluorescent (IF) Staining
2.8. Statisitc Analysis
3. Results
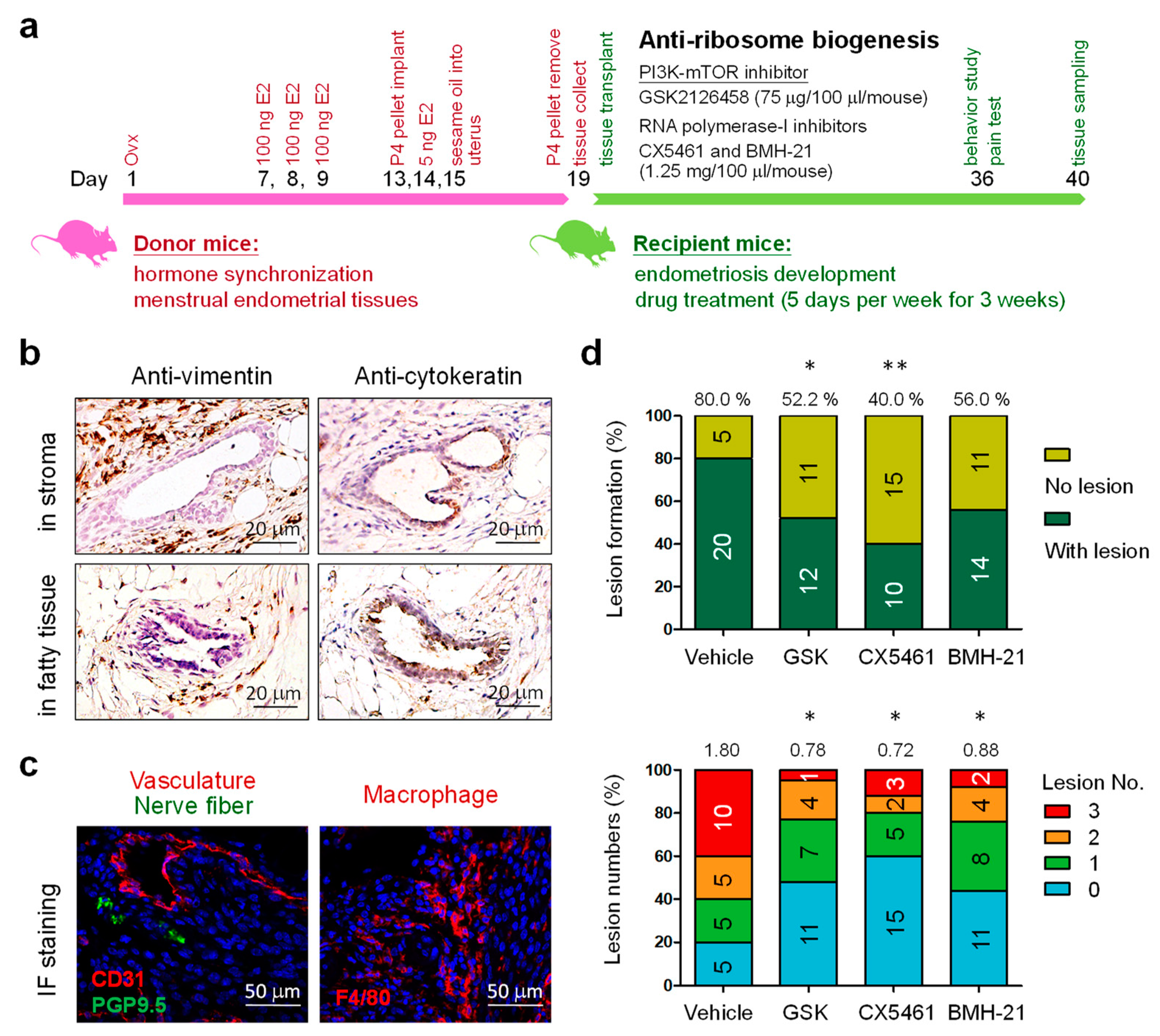
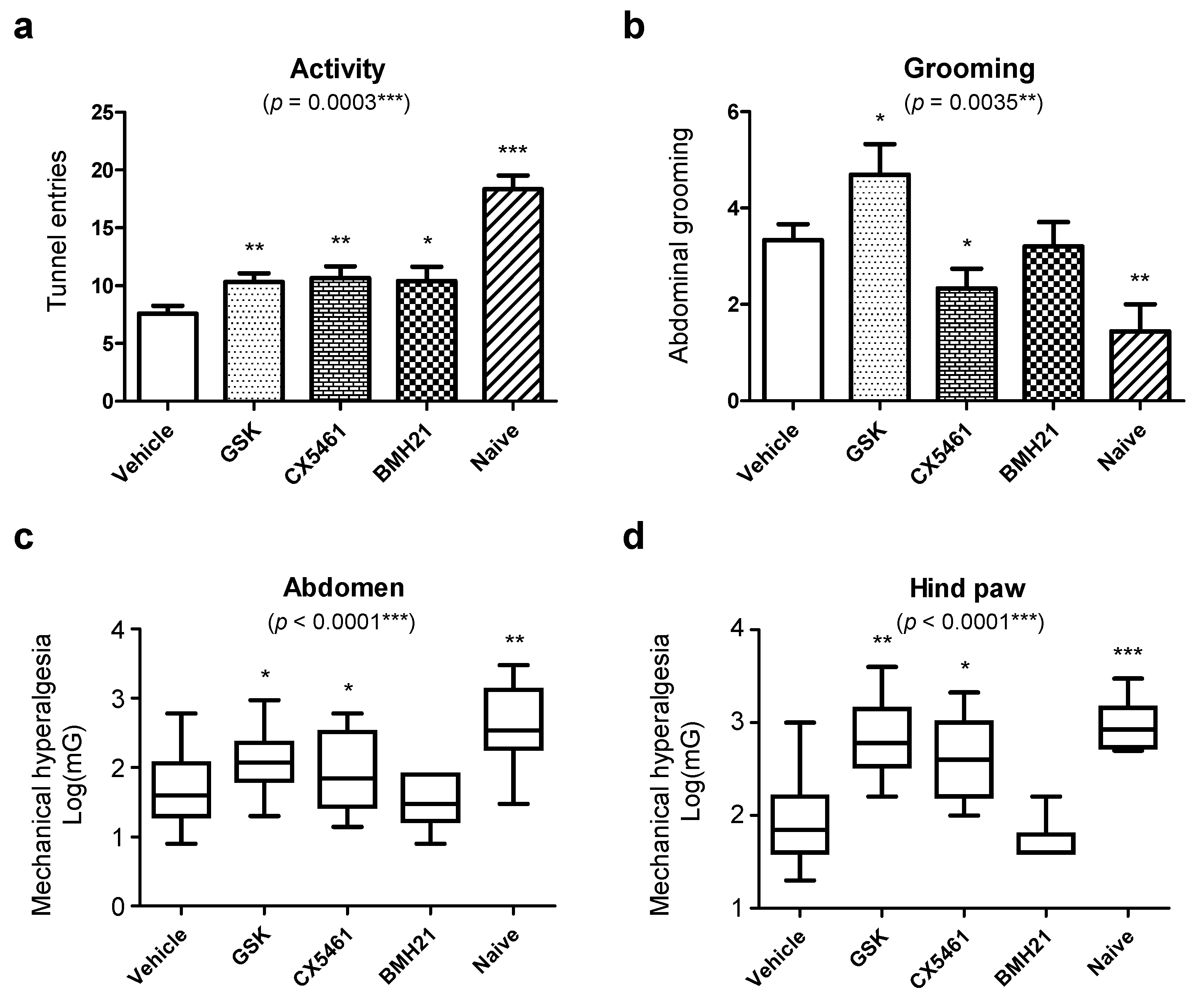
3.1. Blockage of Ribosome Biogenesis Led to Reduced Formation of Endometriotic Lesions
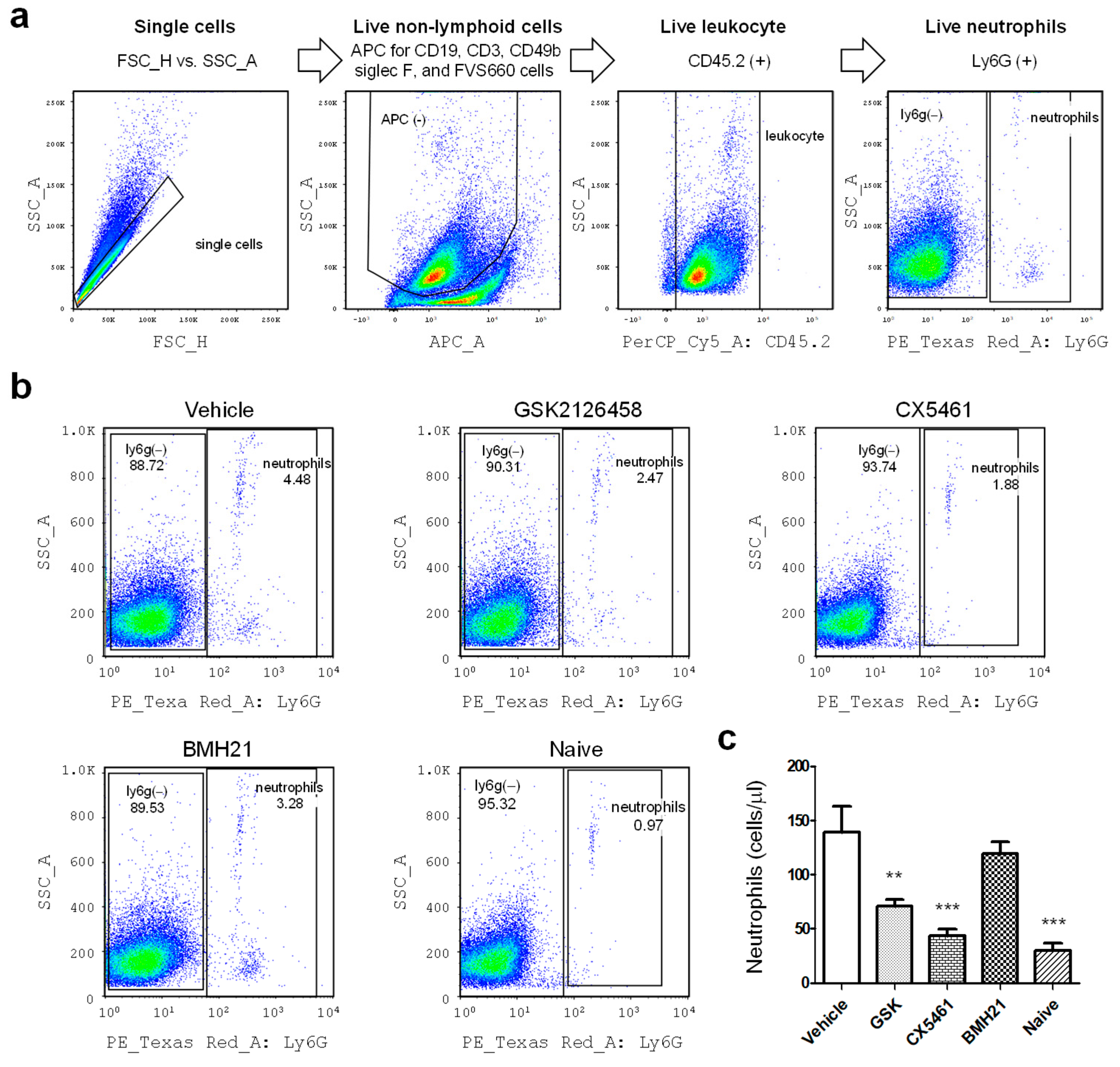
3.2. Anti-Ribosome Biogenesis Modifies the Peritoneal Immune Milieu
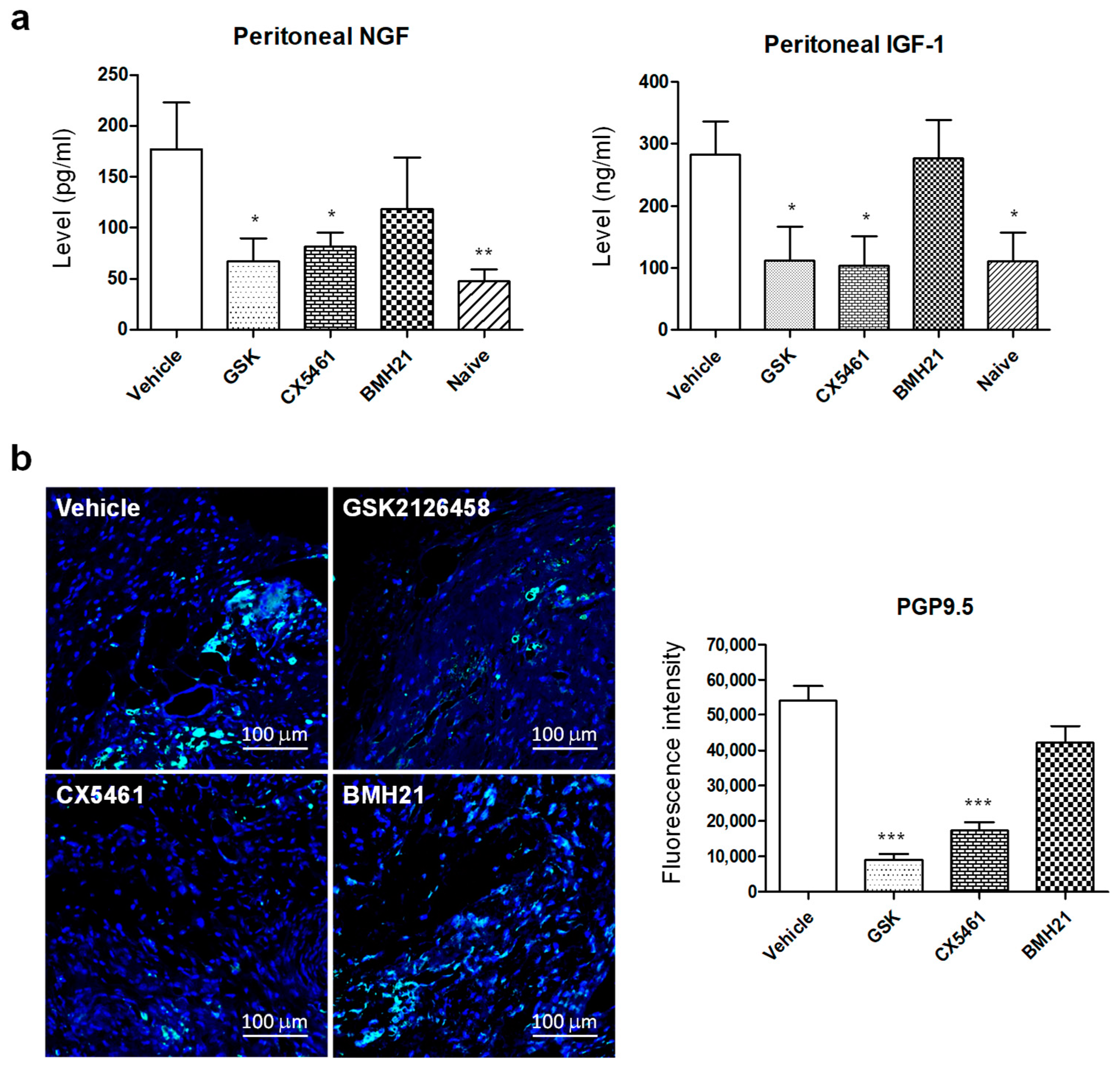
3.3. Anti-Ribosome Biogenesis Agents Suppressed Concentrations of Peritoneal Neurotrophic Factors and Nerve Fiber Growth in Endometriotic Lesions
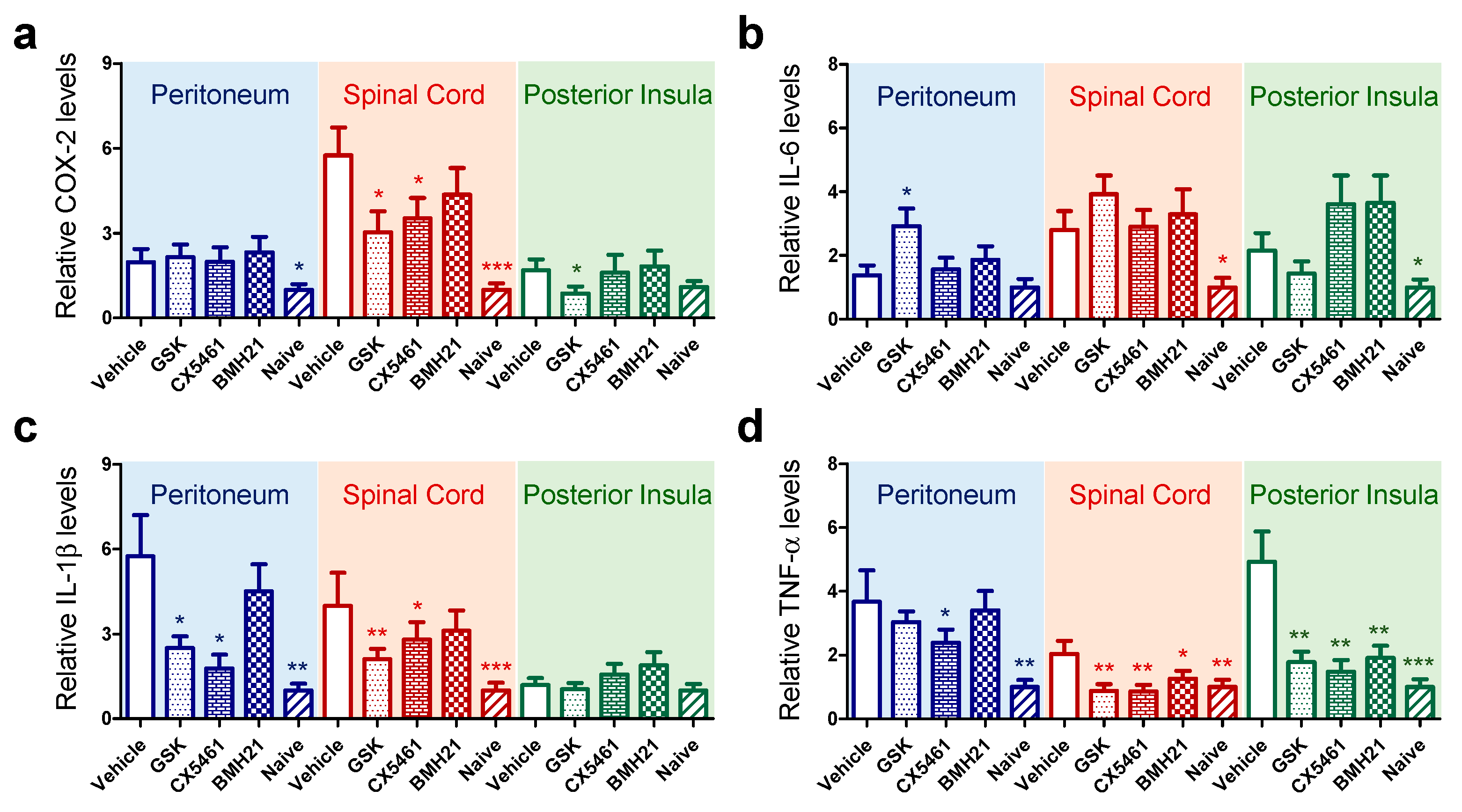
3.4. Anti-Ribosome Biogenesis Suppressed Chronic Inflammation-Induced Hyperalgesia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Nicholes, K.; Shih, I.M. The Origin and Pathogenesis of Endometriosis. Annu. Rev. Pathol. Mech. 2020, 15, 71–95. [Google Scholar] [CrossRef] [Green Version]
- Bulun, S.E.; Yilmaz, B.D.; Sison, C.; Miyazaki, K.; Bernardi, L.; Liu, S.; Kohlmeier, A.; Yin, P.; Milad, M.; Wei, J. Endometriosis. Endocr. Rev. 2019, 40, 1048–1079. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.N.; Yu, J.; Torres, P.B.; Schickedanz, A.C.; Park, J.K.; Mueller, M.D.; Sidell, N. Mechanistic and Therapeutic Implications of Angiogenesis in Endometriosis. Reprod. Sci. 2009, 16, 140–146. [Google Scholar] [CrossRef]
- Wells, M. Recent advances in endometriosis with emphasis on pathogenesis, molecular pathology, and neoplastic transformation. Int. J. Gynecol. Pathol. 2004, 23, 316–320. [Google Scholar] [CrossRef]
- Wilbur, M.A.; Shih, I.M.; Segars, J.H.; Fader, A.N. Cancer Implications for Patients with Endometriosis. Semin. Reprod. Med. 2017, 35, 110–116. [Google Scholar] [CrossRef]
- Prowse, A.H.; Manek, S.; Varma, R.; Liu, J.; Godwin, A.K.; Maher, E.R.; Tomlinson, I.P.; Kennedy, S.H. Molecular genetic evidence that endometriosis is a precursor of ovarian cancer. Int. J. Cancer 2006, 119, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Dupuis-Sandoval, F.; Poirier, M.; Scott, M.S. The emerging landscape of small nucleolar RNAs in cell biology. Wiley Interdiscip. Rev. RNA 2015, 6, 381–397. [Google Scholar] [CrossRef] [PubMed]
- Lafontaine, D.L.J. Noncoding RNAs in eukaryotic ribosome biogenesis and function. Nat. Struct. Mol. Biol. 2015, 22, 11–19. [Google Scholar] [CrossRef]
- Pelletier, J.; Thomas, G.; Volarevic, S. Ribosome biogenesis in cancer: New players and therapeutic avenues. Nat. Rev. Cancer 2018, 18, 51–63. [Google Scholar] [CrossRef]
- Su, H.; Xu, T.; Ganapathy, S.; Shadfan, M.; Long, M.; Huang, T.H.; Thompson, I.; Yuan, Z.M. Elevated snoRNA biogenesis is essential in breast cancer. Oncogene 2014, 33, 1348–1358. [Google Scholar] [CrossRef]
- Iadevaia, V.; Liu, R.; Proud, C.G. mTORC1 signaling controls multiple steps in ribosome biogenesis. Semin. Cell Dev. Biol. 2014, 36, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Lai, M.T.; Chen, Y.; Yang, C.W.; Chang, H.W.; Lu, C.C.; Chen, C.M.; Chan, C.; Chung, C.; Tseng, C.C.; et al. Up-regulation of ribosome biogenesis by MIR196A2 genetic variation promotes endometriosis development and progression. Oncotarget 2016, 7, 76713–76725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, C.M.; Barbara, C.; Prentice, A.; Brenton, J.D.; Charnock-Jones, D.S. Models of endometriosis and their utility in studying progression to ovarian clear cell carcinoma. J. Pathol. 2016, 238, 185–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greaves, E.; Critchley, H.O.D.; Horne, A.W.; Saunders, P.T.K. Relevant human tissue resources and laboratory models for use in endometriosis research. Acta Obstet. Gynecol. Scand. 2017, 96, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Greaves, E.; Cousins, F.L.; Murray, A.; Esnal-Zufiaurre, A.; Fassbender, A.; Horne, A.W.; Saunders, P.T. A novel mouse model of endometriosis mimics human phenotype and reveals insights into the inflammatory contribution of shed endometrium. Am. J. Pathol. 2014, 184, 1930–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greaves, E.; Horne, A.W.; Jerina, H.; Mikolajczak, M.; Hilferty, L.; Mitchell, R.; Fleetwood-Walker, S.M.; Saunders, P.T. EP(2) receptor antagonism reduces peripheral and central hyperalgesia in a preclinical mouse model of endometriosis. Sci. Rep. 2017, 7, 44169. [Google Scholar] [CrossRef] [Green Version]
- Woods, S.J.; Hannan, K.M.; Pearson, R.B.; Hannan, R.D. The nucleolus as a fundamental regulator of the p53 response and a new target for cancer therapy. Biochim. Biophys. Acta 2015, 1849, 821–829. [Google Scholar] [CrossRef]
- Deisenroth, C.; Zhang, Y. Ribosome biogenesis surveillance: Probing the ribosomal protein-Mdm2-p53 pathway. Oncogene 2010, 29, 4253–4260. [Google Scholar] [CrossRef] [Green Version]
- Poortinga, G.; Quinn, L.M.; Hannan, R.D. Targeting RNA polymerase I to treat MYC-driven cancer. Oncogene 2015, 34, 403–412. [Google Scholar] [CrossRef]
- Horne, A.W.; Ahmad, S.F.; Carter, R.; Simitsidellis, I.; Greaves, E.; Hogg, C.; Morton, N.M.; Saunders, P.T.K. Repurposing dichloroacetate for the treatment of women with endometriosis. Proc. Natl. Acad. Sci. USA 2019, 116, 25389–25391. [Google Scholar] [CrossRef] [Green Version]
- Knight, S.D.; Adams, N.D.; Burgess, J.L.; Chaudhari, A.M.; Darcy, M.G.; Donatelli, C.A.; Luengo, J.I.; Newlander, K.A.; Parrish, C.A.; Ridgers, L.H.; et al. Discovery of GSK2126458, a Highly Potent Inhibitor of PI3K and the Mammalian Target of Rapamycin. ACS Med. Chem. Lett. 2010, 1, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Bywater, M.J.; Poortinga, G.; Sanij, E.; Hein, N.; Peck, A.; Cullinane, C.; Wall, M.; Cluse, L.; Drygin, D.; Anderes, K.; et al. Inhibition of RNA polymerase I as a therapeutic strategy to promote cancer-specific activation of p53. Cancer Cell 2012, 22, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Forster, R.; Sarginson, A.; Velichkova, A.; Hogg, C.; Dorning, A.; Horne, A.W.; Saunders, P.T.K.; Greaves, E. Macrophage-derived insulin-like growth factor-1 is a key neurotrophic and nerve-sensitizing factor in pain associated with endometriosis. FASEB J. 2019, 33, 11210–11222. [Google Scholar] [CrossRef] [Green Version]
- Ghosn, E.E.; Cassado, A.A.; Govoni, G.R.; Fukuhara, T.; Yang, Y.; Monack, D.M.; Bortoluci, K.R.; Almeida, S.R.; Herzenberg, L.A.; Herzenberg, L.A. Two physically, functionally, and developmentally distinct peritoneal macrophage subsets. Proc. Natl. Acad. Sci. USA 2010, 107, 2568–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassado Ados, A.; D’Imperio Lima, M.R.; Bortoluci, K.R. Revisiting mouse peritoneal macrophages: Heterogeneity, development, and function. Front. Immunol. 2015, 6, 225. [Google Scholar] [CrossRef] [Green Version]
- Barcena de Arellano, M.L.; Arnold, J.; Lang, H.; Vercellino, G.F.; Chiantera, V.; Schneider, A.; Mechsner, S. Evidence of neurotrophic events due to peritoneal endometriotic lesions. Cytokine 2013, 62, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Greaves, E.; Collins, F.; Esnal-Zufiaurre, A.; Giakoumelou, S.; Horne, A.W.; Saunders, P.T. Estrogen receptor (ER) agonists differentially regulate neuroangiogenesis in peritoneal endometriosis via the repellent factor SLIT3. Endocrinology 2014, 155, 4015–4026. [Google Scholar] [CrossRef] [PubMed]
- Morotti, M.; Vincent, K.; Becker, C.M. Mechanisms of pain in endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 209, 8–13. [Google Scholar] [CrossRef]
- As-Sanie, S.; Kim, J.; Schmidt-Wilcke, T.; Sundgren, P.C.; Clauw, D.J.; Napadow, V.; Harris, R.E. Functional Connectivity is Associated With Altered Brain Chemistry in Women With Endometriosis-Associated Chronic Pelvic Pain. J. Pain 2016, 17, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Kobayashi, H.; Kajiwara, H.; Kanayama, S.; Yamada, Y.; Furukawa, N.; Noguchi, T.; Haruta, S.; Yoshida, S.; Sakata, M.; Sado, T.; et al. Molecular pathogenesis of endometriosis-associated clear cell carcinoma of the ovary (review). Oncol. Rep. 2009, 22, 233–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurman, R.J.; Shih, I.M. Molecular pathogenesis and extraovarian origin of epithelial ovarian cancer—Shifting the paradigm. Hum. Pathol. 2011, 42, 918–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, I.; Bast, R.C., Jr. Minireview: Human ovarian cancer: Biology, current management, and paths to personalizing therapy. Endocrinology 2012, 153, 1593–1602. [Google Scholar] [CrossRef] [Green Version]
- Buerger, C.; Shirsath, N.; Lang, V.; Berard, A.; Diehl, S.; Kaufmann, R.; Boehncke, W.H.; Wolf, P. Inflammation dependent mTORC1 signaling interferes with the switch from keratinocyte proliferation to differentiation. PLoS ONE 2017, 12, e0180853. [Google Scholar] [CrossRef] [Green Version]
- Brighenti, E.; Calabrese, C.; Liguori, G.; Giannone, F.A.; Trere, D.; Montanaro, L.; Derenzini, M. Interleukin 6 downregulates p53 expression and activity by stimulating ribosome biogenesis: A new pathway connecting inflammation to cancer. Oncogene 2014, 33, 4396–4406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brighenti, E.; Giannone, F.A.; Fornari, F.; Onofrillo, C.; Govoni, M.; Montanaro, L.; Treré, D.; Derenzini, M. Therapeutic dosages of aspirin counteract the IL-6 induced pro-tumorigenic effects by slowing down the ribosome biogenesis rate. Oncotarget 2016, 7, 63226–63241. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Chen, J.; Xiong, J.W.; Peng, J. Haploinsufficiency of Def activates p53-dependent TGFbeta signalling and causes scar formation after partial hepatectomy. PLoS ONE 2014, 9, e96576. [Google Scholar]
- Huang, W.; Chen, F.; Ma, Q.; Xin, J.; Li, J.; Chen, J.; Zhou, B.; Chen, M.; Li, J.; Peng, J. Ribosome biogenesis gene DEF/UTP25 is essential for liver homeostasis and regeneration. Sci. China Life Sci. 2020, 63, 1651–1664. [Google Scholar] [CrossRef]
- Li, H.; Li, Q.; Li, W.; Xie, L.; Zhou, M.; Xie, J. The Role of PARP-1 in Host-Pathogen Interaction and Cellular Stress Responses. Crit. Rev. Eukaryot. Gene Expr. 2015, 25, 175–190. [Google Scholar] [CrossRef]
- Huang, K.Y.; Shin, J.W.; Huang, P.J.; Ku, F.M.; Lin, W.C.; Lin, R.; Hsu, W.M.; Tang, P. Functional profiling of the Tritrichomonas foetus transcriptome and proteome. Mol. Biochem. Parasitol. 2013, 187, 60–71. [Google Scholar] [CrossRef]
- Chernoff, N.; Rogers, E.H.; Zehr, R.D.; Gage, M.I.; Malarkey, D.E.; Bradfield, C.A.; Liu, Y.; Schmid, J.E.; Jaskot, R.H.; Richards, J.H.; et al. Toxicity and recovery in the pregnant mouse after gestational exposure to the cyanobacterial toxin, cylindrospermopsin. J. Appl. Toxicol. 2011, 31, 242–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Yang, Y.; Zhang, J.; Chen, J.; Wei, X.; He, J.; Luo, L. Ribosome biogenesis protein Urb1 acts downstream of mTOR complex 1 to modulate digestive organ development in zebrafish. J. Genet. Genomics 2017, 44, 567–576. [Google Scholar] [CrossRef]
- Lee, D.F.; Kuo, H.P.; Chen, C.T.; Hsu, J.M.; Chou, C.K.; Wei, Y.; Sun, H.L.; Li, L.Y.; Ping, B.; Huang, W.C.; et al. IKK beta suppression of TSC1 links inflammation and tumor angiogenesis via the mTOR pathway. Cell 2007, 130, 440–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Viel, S.; Besson, L.; Marotel, M.; Walzer, T.; Marcais, A. Regulation of mTOR, Metabolic Fitness, and Effector Functions by Cytokines in Natural Killer Cells. Cancers 2017, 9, 132. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.I.; Yeo, S.G.; Gen, Y.; Ju, H.R.; Kim, S.H.; Park, D.C. Differences in autophagy-associated mRNAs in peritoneal fluid of patients with endometriosis and gynecologic cancers. Eur. J. Obstet. Gynecol. Reprod. Biol. X 2019, 2, 100016. [Google Scholar] [CrossRef] [PubMed]
- Rogers-Broadway, K.R.; Kumar, J.; Sisu, C.; Wander, G.; Mazey, E.; Jeyaneethi, J.; Pados, G.; Tsolakidis, D.; Klonos, E.; Grunt, T.; et al. Differential expression of mTOR components in endometriosis and ovarian cancer: Effects of rapalogues and dual kinase inhibitors on mTORC1 and mTORC2 stoichiometry. Int. J. Mol. Med. 2019, 43, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Poli-Neto, O.B.; Meola, J.; Rosa, E.S.J.C.; Tiezzi, D. Transcriptome meta-analysis reveals differences of immune profile between eutopic endometrium from stage I-II and III-IV endometriosis independently of hormonal milieu. Sci. Rep. 2020, 10, 313. [Google Scholar] [CrossRef]
- Yagyu, T.; Tsuji, Y.; Haruta, S.; Kitanaka, T.; Yamada, Y.; Kawaguchi, R.; Kanayama, S.; Tanase, Y.; Kurita, N.; Kobayashi, H. Activation of mammalian target of rapamycin in postmenopausal ovarian endometriosis. Int. J. Gynecol. Cancer 2006, 16, 1545–1551. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Jo, M.; Lee, E.; Lee, D.Y.; Choi, D. Involvement of endoplasmic reticulum stress in regulation of endometrial stromal cell invasiveness: Possible role in pathogenesis of endometriosis. Mol. Hum. Reprod. 2019, 25, 101–110. [Google Scholar] [CrossRef]
- McKinnon, B.D.; Kocbek, V.; Nirgianakis, K.; Bersinger, N.A.; Mueller, M.D. Kinase signalling pathways in endometriosis: Potential targets for non-hormonal therapeutics. Hum. Reprod. Update 2016, 22, 382–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leconte, M.; Nicco, C.; Ngo, C.; Chereau, C.; Chouzenoux, S.; Marut, W.; Guibourdenche, J.; Arkwright, S.; Weill, B.; Chapron, C.; et al. The mTOR/AKT inhibitor temsirolimus prevents deep infiltrating endometriosis in mice. Am. J. Pathol. 2011, 179, 880–889. [Google Scholar] [CrossRef] [PubMed]
- Kacan, T.; Yildiz, C.; Baloglu Kacan, S.; Seker, M.; Ozer, H.; Cetin, A. Everolimus as an mTOR Inhibitor Suppresses Endometriotic Implants: An Experimental Rat Study. Geburtshilfe Frauenheilkd. 2017, 77, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, S.; Pouly, J.L.; Canis, M. In vitro and in vivo effects of MK2206 and chloroquine combination therapy on endometriosis: Autophagy may be required for regrowth of endometriosis. Br. J Pharmacol. 2018, 175, 1637–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Qin, X.; Lu, X.; Jiang, J. Effects of inhibiting the PI3K/Akt/mTOR signaling pathway on the pain of sciatic endometriosis in a rat model. Can. J. Physiol. Pharmacol. 2019, 97, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Laganà, A.S.; Salmeri, F.M.; Ban Frangež, H.; Ghezzi, F.; Vrtačnik-Bokal, E.; Granese, R. Evaluation of M1 and M2 macrophages in ovarian endometriomas from women affected by endometriosis at different stages of the disease. Gynecol. Endocrinol. 2020, 36, 441–444. [Google Scholar] [CrossRef]
- Laganà, A.S.; Salmeri, F.M.; Vitale, S.G.; Triolo, O.; Götte, M. Stem Cell Trafficking During Endometriosis: May Epigenetics Play a Pivotal Role? Reprod. Sci. 2018, 25, 978–979. [Google Scholar] [CrossRef]
- Pluchino, N.; Taylor, H.S. Endometriosis and Stem Cell Trafficking. Reprod. Sci. 2016, 23, 1616–1619. [Google Scholar] [CrossRef]
- Turi, Z.; Lacey, M.; Mistrik, M.; Moudry, P. Impaired ribosome biogenesis: Mechanisms and relevance to cancer and aging. Aging 2019, 11, 2512–2540. [Google Scholar] [CrossRef]
- Bassler, J.; Hurt, E. Eukaryotic Ribosome Assembly. Annu. Rev. Biochem. 2019, 88, 281–306. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, C.Y.-Y.; Chiang, A.-J.; Yan, M.-J.; Lai, M.-T.; Su, Y.-Y.; Huang, H.-Y.; Chang, C.-Y.; Li, Y.-H.; Li, P.-F.; Chen, C.-M.; et al. Ribosome Biogenesis Serves as a Therapeutic Target for Treating Endometriosis and the Associated Complications. Biomedicines 2022, 10, 185. https://doi.org/10.3390/biomedicines10010185
Chang CY-Y, Chiang A-J, Yan M-J, Lai M-T, Su Y-Y, Huang H-Y, Chang C-Y, Li Y-H, Li P-F, Chen C-M, et al. Ribosome Biogenesis Serves as a Therapeutic Target for Treating Endometriosis and the Associated Complications. Biomedicines. 2022; 10(1):185. https://doi.org/10.3390/biomedicines10010185
Chicago/Turabian StyleChang, Cherry Yin-Yi, An-Jen Chiang, Man-Ju Yan, Ming-Tsung Lai, Yun-Yi Su, Hsin-Yi Huang, Chan-Yu Chang, Ya-Hui Li, Pei-Fen Li, Chih-Mei Chen, and et al. 2022. "Ribosome Biogenesis Serves as a Therapeutic Target for Treating Endometriosis and the Associated Complications" Biomedicines 10, no. 1: 185. https://doi.org/10.3390/biomedicines10010185