

Differential and Time-Discrete SEIRS Models with Vaccination: Local Stability, Validation and Sensitivity Analysis Using Bulgarian COVID-19 Data


Abstract
:1. Introduction
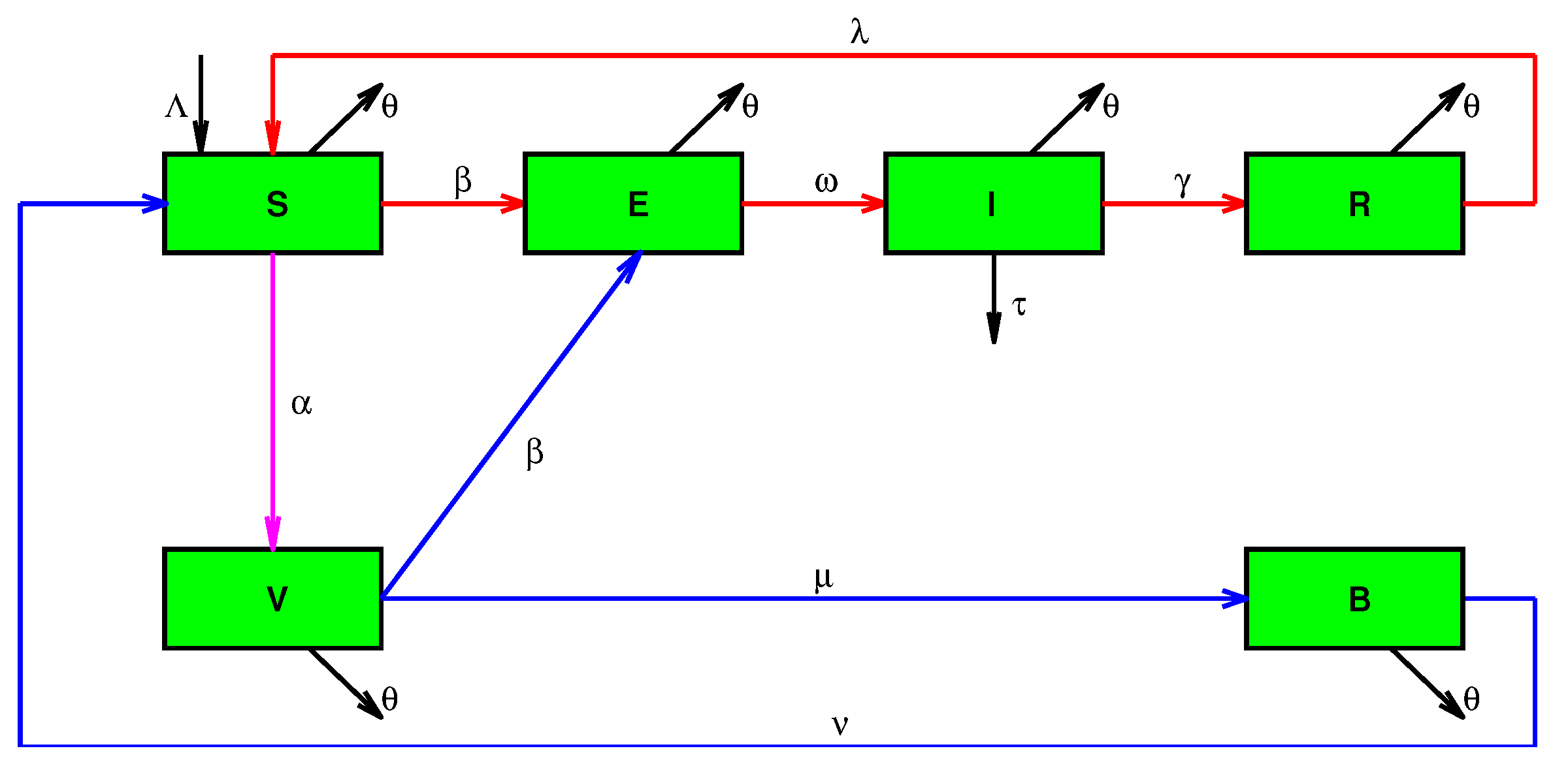
2. The Differential SEIR-VB Model
- —susceptible individuals (unvaccinated, not fully vaccinated, vaccinated people for whom the vaccine is ineffective, fully vaccinated or recovered individuals who have lost their immunity).
- —exposed individuals. These are virus carriers in the latent stage, during which they are not virus spreaders. They usually have no symptoms.
- —infectious individuals. These are virus carriers and virus spreaders of extremely high infectivity. The former are likely to transmit the virus in case of contact.
- —Recovered individuals with disease-acquired immunity. These individuals have disease-acquired immunity. They have recovered, and thus are protected from the disease.
- —vaccinated susceptible individuals. These are fully vaccinated individuals for whom the vaccine is effective. However, they have not developed antibodies. They can do so after a certain period of time or otherwise they will become exposed individuals before that. It is worth pointing out that, due to vaccine imperfections, some of the vaccinated individuals cannot develop antibodies, and they cannot pass from group to group .
- —vaccinated individuals with vaccination-acquired immunity. These are vaccinated individuals who are well protected from future infection because they have antibodies.
- —birth rate;
- —vaccination rate;
- —vaccine effectiveness;
- —vaccination parameter;
- —transmission rate;
- —recovery rate;
- —latency rate;
- —natural mortality rate;
- —mortality rate of infectious people;
- —reinfection rate of recovered individuals;
- —reinfection rate of vaccinated individuals;
- —antibody rate.
3. SEIRS-VB Model with Time-Independent Coefficients
4. The Disease-Free Equilibrium Point and Basic Reproduction Number
5. Local Stability Analysis of the Disease-Free Equilibrium Point
6. Time-Discrete SEIRS-VB Model
6.1. Construction of a New Family of Semi-Implicit Difference Schemes
6.2. Parameter Identification Algorithm
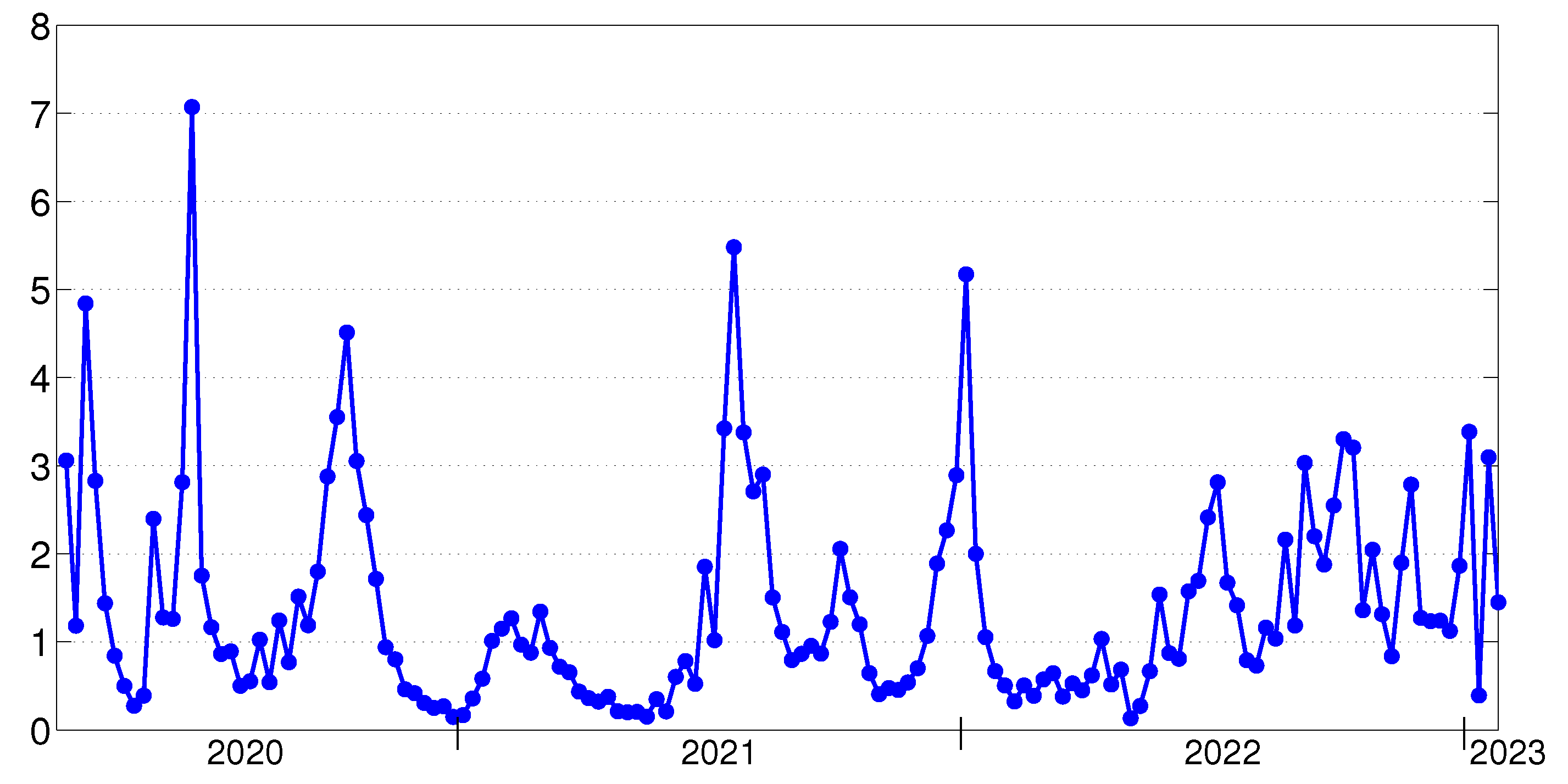
- is the number of the active cases.
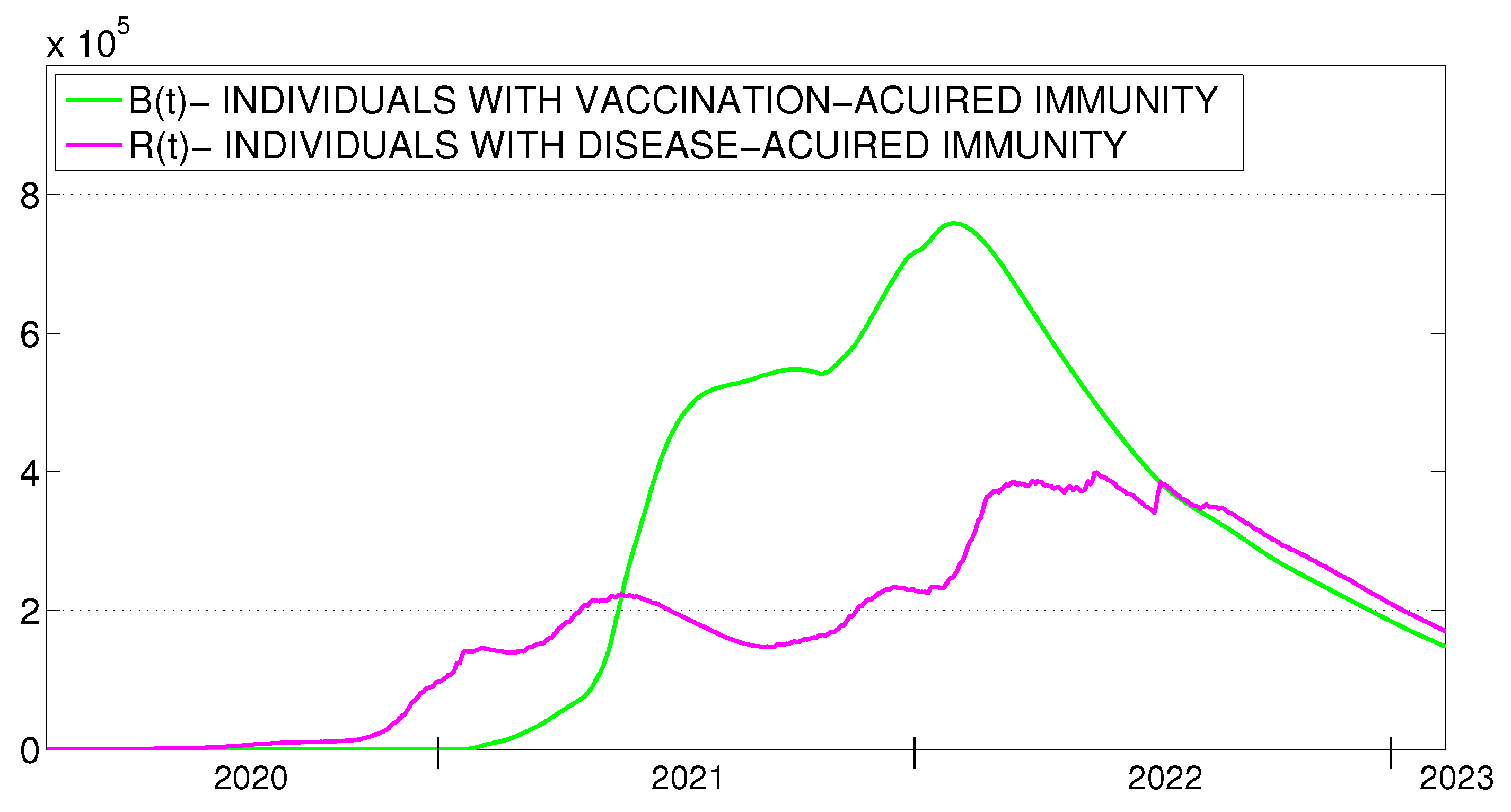
- is the cumulative number of the individuals recovered from the disease to time . Unlike , individuals who have already lost disease-acquired immunity are counted in .
- is the cumulative number of COVID-19 deaths.
- is the cumulative number of the fully vaccinated individuals.
- Since the values and are given and non-decreasing with respect to k, we find via (23) the non-negative values , .
- Since the values are given, we find the values of the vaccination parameterwhere is the vaccine effectiveness.
- Now, we are able to calculate the number of exposed individuals
- Finally, if and , we calculatefor
- If one or more of the calculated values are negative or some of the values , , are equal to zero for some k, the algorithm must stop. Otherwise, the algorithm continues and the problem can be solved.
7. Validation of SEIRS-VB Models
7.1. Time-Discrete SEIRS-VB Model
- The following parameters are constant during the time period under consideration:
- is the average birth rate for 2015–2020;
- is the average natural mortality rate for 2015–2020;
- , where is the incubation (latency) period for the dominant variant of SARS-CoV-2;
- , where is the average time taken for antibodies to develop;
- , where is the duration of the immune responses in individuals with vaccination-acquired immunity;
- , where is the duration of the immune responses in recovered individuals.
- Several vaccines were in use during the COVID-19 mass vaccination campaign in Bulgaria—Comirnaty (Pfizer/BioNTech), Comirnaty Original/Omicron BA.1, Comirnaty Original/Omicron BA.4-5, Spikevax (COVID-19 Vaccine Moderna), Spikevax Bivalent Original/Omicron BA.1, COVID-19 Vaccine Janssen, Vaxzevria (AstraZeneca), Nuvaxovid (NVX-CoV2373), and COVID-19 Vaccine (inactivated, adjuvanted) Valneva, Vidprevtyn Beta. Taking into account the number of the fully vaccinated and boosted people with these vaccines, and the product information in [43], we applied the average parameters specified in Table 1.
7.2. Differential SEIRS-VB Model
8. Parameter Sensitivity Analysis
9. Discussion
10. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dietz, K.; Heesterbeek, J.A.P. Daniel Bernoulli’s epidemiological model revisited. Math. Biosci. 2002, 180, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Kermack, W.; McKendrick, A. A Contribution to the Mathematical Theory of Pandemics. Proc. R. Soc. Lond. Ser. A 1927, 115, 700–721. [Google Scholar] [CrossRef]
- Khan, A.H. Modeling the Spread of COVID-19 Pandemic in Morocco. Challenges in Modeling of an Outbreak’s Prediction, Forecasting and Decision Making for Policy Makers, Infosys Science Foundation Series in Mathematical Sciences; Springer: Singapore, 2021; pp. 377–408. [Google Scholar] [CrossRef]
- Margenov, S.; Popivanov, N.; Ugrinova, I.; Hristov, T. Mathematical Modeling and Short-Term Forecasting of the COVID-19 Epidemic in Bulgaria: SEIRS Model with Vaccination. Mathematics 2022, 10, 2570. [Google Scholar] [CrossRef]
- Kabanikhin, S.; Bektemessov, I.; Krivorotko, O.; Bektemessov, Z. Determination of the coefficients of nonlinear ordinary differential equations systems using additional statistical information. Int. J. Math. Phys. 2019, 10, 36–42. [Google Scholar] [CrossRef]
- Krivorotko, O.; Kabanikhin, S.; Sosnovskaya, M.; Andornaya, D. Sensitivity and Identifiability Analysis of COVID-19 Pandemic Models. Vavilov J. Genet. Breed. 2021, 25, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Marinov, T.; Marinova, R. Dynamics of COVID-19 using inverse problem for coefficient identification in SIR epidemic models. Chaos Solitons Fractals X 2020, 5, 100041. [Google Scholar] [CrossRef]
- Marinov, T.; Marinova, R. Inverse problem for adaptive SIR model: Application to COVID-19 in Latin America. Infect. Dis. Model. 2022, 5, 134–148. [Google Scholar] [CrossRef]
- Leonov, A.S.; Nagornov, O.V.; Tyuflin, S.A. Inverse problem for coefficients of equations describing propagation of COVID-19 epidemic. J. Phys. Conf. Ser. 2021, 2036, 012028. [Google Scholar] [CrossRef]
- Hethcote, H. The Mathematics of Infectious Diseases. Siam Rev. 2000, 42, 599–653. Available online: https://www.jstor.org/stable/2653135 (accessed on 15 February 2023). [CrossRef]
- Li, M.; Muldowney, J. Global stability for the SEIR model in epidemiology. Math. Biosci. 1995, 125, 155–164. [Google Scholar] [CrossRef]
- Korobeinikov, A.; Maini, P. A Lyapunov function and global properties for SIR and SEIR epidemiological models with nonlinear incidence. Math. Biosci. Eng. 2004, 1, 57–60. [Google Scholar] [CrossRef]
- Ghostine, R.; Gharamti, M.; Hassrouny, S.; Hoteit, I. An Extended SEIR Model with Vaccination for Forecasting the COVID-19 Pandemic in Saudi Arabia Using an Ensemble Kalman Filter. Mathematics 2021, 9, 636. [Google Scholar] [CrossRef]
- Al-Shbeil, I.; Djenina, N.; Jaradat, A.; Al-Husban, A.; Ouannas, A.; Grassi, G. A New COVID-19 Pandemic Model Including the Compartment of Vaccinated Individuals: Global Stability of the Disease-Free Fixed Point. Mathematics 2023, 11, 576. [Google Scholar] [CrossRef]
- Xu, D.-G.; Xu, X.-Y.; Yang, C.-H.; Gui, W.-H. Global Stability of a Variation Epidemic Spreading Model on Complex Networks. Math. Probl. Eng. 2015, 2015, 365049. [Google Scholar] [CrossRef]
- Wangari, I. Condition for Global Stability for a SEIR Model Incorporating Exogenous Reinfection and Primary Infection Mechanisms. Comput. Math. Methods Med. 2020, 2020, 9435819. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Wang, L. Global Stability in Some Seir Epidemic Models. In Mathematical Approaches for Emerging and Reemerging Infectious Diseases: Models, Methods, and Theory. The IMA Volumes in Mathematics and its Applications 126; Springer: New York, NY, USA, 2002. [Google Scholar] [CrossRef]
- Lobatog, F.; Plattg, M.; Libottea, B.; Silva Neto, A. Formulation and Solution of an Inverse Reliability Problem to Simulate the Dynamic Behavior of COVID-19 Pandemic. Trends Comput. Appl. Math. 2021, 22, 91–107. [Google Scholar] [CrossRef]
- Georgiev, S.; Vulkov, L. Coefficient Identification for SEIR Model and Economic Forecasting in the Propagation of COVID-19. In Advanced Computing in Industrial Mathematics, Studies in Computational Intelligence; Springer: Berlin/Heidelberg, Germany, 2023; Volume 1076, pp. 34–44. Available online: https://link.springer.com/content/pdf/10.1007/978-3-031-20951-2_4 (accessed on 16 February 2023).
- Ibeas, A.; De la Sen, M.; Alonso-Quesada, S.; Zamani, I.; Shafiee, M. Observer design for SEIR discrete-time epidemic models. In Proceedings of the 13th International Conference on Control Automation Robotics & Vision (ICARCV), Singapore, 10–12 December 2014; pp. 1321–1326. [Google Scholar] [CrossRef]
- Leonov, A.; Nagornov, O.; Tyuflin, S. Modeling of Mechanisms of Wave Formation for COVID-19 Epidemic. Mathematics 2023, 11, 167. [Google Scholar] [CrossRef]
- Li, B.; Eskandari, Z.; Avazzadeh, Z. Dynamical Behaviors of an SIR Epidemic Model with Discrete Time. Fractal Fract. 2022, 6, 659. [Google Scholar] [CrossRef]
- Carcione, J.; Santos, J.; Bagaini, C.; Ba, J. A Simulation of a COVID-19 Epidemic Based on a Deterministic SEIR Model. Front. Public Health 2020, 8, 230. [Google Scholar] [CrossRef]
- Khalsaraei, M.; Shokri, A.; Ramos, H.; Yao, S.-W.; Molayi, M. Efficient Numerical Solutions to a SIR Epidemic Model. Mathematics 2022, 10, 3299. [Google Scholar] [CrossRef]
- Costa, J.A.; Martinez, A.C.; Geromel, J.C. On the Continuous-time and Discrete-Time Versions of an Alternative Epidemic Model of the SIR Class. J. Control Electr. Syst. 2022, 33, 38–48. [Google Scholar] [CrossRef]
- Qin, H.; Chen, X.; Zhou, B. A Family of Transformed Difference Schemes for Nonlinear Time-Fractional Equations. Fractal Fract. 2023, 7, 96. [Google Scholar] [CrossRef]
- Alharbi, W.; Shater, A.; Ebaid, A.; Cattani, C.; Areshi, M.; Jalal, M.; Alharbi, M. Communicable disease model in view of fractional calculus. AIMS Math. 2023, 8, 10033–10048. [Google Scholar] [CrossRef]
- He, Z.-Y.; Abbes, A.; Jahanshahi, H.; Alotaibi, N.D.; Wang, Y. Fractional-Order Discrete-Time SIR Epidemic Model with Vaccination: Chaos and Complexity. Mathematics 2022, 10, 165. [Google Scholar] [CrossRef]
- Islam, M.R.; Peace, A.; Medina, D.; Oraby, T. Integer Versus Fractional Order SEIR Deterministic and Stochastic Models of Measles. Int. J. Environ. Res. Public Health 2020, 17, 2014. [Google Scholar] [CrossRef] [PubMed]
- De la Sen, M.; Alonso-Quesada, S.; Ibeas, A. On a Discrete SEIR Epidemic Model with Exposed Infectivity, Feedback Vaccination and Partial Delayed Re-Susceptibility. Mathematics 2021, 9, 520. [Google Scholar] [CrossRef]
- Singh, R.A.; Lal, R.; Kotti, R.R. Time-discrete SIR model for COVID-19 in Fiji. Epidemiol. Infect. 2022, 150, e75. [Google Scholar] [CrossRef]
- Wacker, B.; Schlüter, J. Time-continuous and time-discrete SIR models revisited: Theory and applications. Adv. Differ. Eqs. 2020, 2020, 556. [Google Scholar] [CrossRef]
- Zhao, Z.; Niu, Y.; Luo, L.; Hu, Q.; Yang, T.; Chu, M.; Chen, Q.; Lei, Z.; Rui, J.; Song, C.; et al. The optimal vaccination strategy to control COVID-19: A modeling study in Wuhan City, China. Infect. Dis. Poverty 2021, 10, 140. [Google Scholar] [CrossRef]
- Angelov, G.; Kovacevic, R.; Stilianakis, N.I.; Veliov, V. Optimal vaccination strategies using a distributed model applied to COVID-19. Cent. Eur. J. Oper. Res. 2023, 31, 499–521. [Google Scholar] [CrossRef]
- Heffernan, J.; Smith, R.; Wahl, L. Perspectives on the basic reproductive ratio. J. R. Soc. Interface 2005, 2, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Van Den Driessche, P. Reproduction numbers of infectious disease models. Infect. Dis. Model. 2017, 2, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Barbashin, E. Introduction to the Theory of Stability; Wolters-Noordhoff Publishing: Groningen, The Netherlands, 1970; p. 223. [Google Scholar]
- Tiwari, S.; Porwal, P.; Barve, T. Transmission Dynamics of Coronavirus and the Effect of Vaccination Using SEIR Model. Serdica Math. J. 2021, 47, 161–178. [Google Scholar]
- Castillo-Chavez, C.; Feng, Z.; Huang, W. Mathematical approaches for emerging and reemerging infectious diseases: An introduction. In The IMA Volumes in Mathematics and Its Applications; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2002; pp. 229–250. [Google Scholar]
- Hartman, P. Ordinary Differential Equations, 2nd ed.; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 2002; p. 624. [Google Scholar]
- The Open Data Portal of the Republic of Bulgaria. Available online: https://data.egov.bg (accessed on 16 February 2023).
- The Official Bulgarian Unified Information Portal. Available online: https://coronavirus.bg/ (accessed on 16 February 2023).
- European Medicines Agency. Vaccines Authorised in the European Union (EU) to Prevent COVID-19. Available online: https://www.ema.europa.eu/en/human-regulatory/overview/public-health-threats/coronavirus-disease-COVID-19/treatments-vaccines/vaccines-COVID-19/COVID-19-vaccines-authorised (accessed on 16 February 2023).
- Deressa, C.; Mussa, Y.; Duressa, G. Optimal control and sensitivity analysis for transmission dynamics of Coronavirus. Results Phys. 2020, 19, 103642. [Google Scholar] [CrossRef]
- Wachira, C.; Lawi, G.; Omondi, L. Sensitivity and Optimal Control Analysis of an Extended SEIR COVID-19 Mathematical Model. J. Math. 2022, 2022, 1476607. [Google Scholar] [CrossRef]
- Ma, C.; Li, X.; Zhao, Z.; Liu, F.; Zhang, K.; Wu, A.; Nie, X. Understanding Dynamics of Pandemic Models to Support Predictions of COVID-19 Transmission: Parameter Sensitivity Analysis of SIR-Type Models. IEEE J. Biomed. Health Inform. 2022, 26, 2458–2468. [Google Scholar] [CrossRef]
- Zine, H.; Lotfi, E.M.; Mahrouf, M.; Boukhouima, A.; Aqachmar, Y.; Hattaf, K.; Torres, D.; Yousfi, N. Modeling the Spread of COVID-19 Pandemic in Morocco. In Analysis of Infectious Disease Problems (COVID-19) and Their Global Impact, Infosys Science Foundation Series in Mathematical Sciences; Springer: Singapore, 2021; pp. 599–616. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Sadoff, J.; Le Gars, M.; Shukarev, G.; Heerwegh, D.; Truyers, C.; de Groot, A.M.; Stoop, J.; Tete, S.; Van Damme, W.; Leroux-Roels, I.; et al. Interim results of a phase 1-2a trial of Ad26.COV2.S COVID-19 vaccine. N. Engl. J. Med. 2021, 384, 1824–1835. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zeng, G.; Pan, H.X.; Li, C.; Hu, Y.; Chu, K.; Han, W.; Chen, Z.; Tang, R.; Yin, W.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18–59 years: A randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect. Dis. 2021, 21, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Margenov, S.; Popivanov, N.; Ugrinova, I.; Harizanov, S.; Hristov, T. Parameters Identification and Forecasting of COVID-19 Transmission Dynamics in Bulgaria with Mass Vaccination Strategy. AIP Conf. Proc. 2022, 2505, 080010. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Values |
|---|---|---|
| birth rate | ||
| natural mortality rate | : | |
| latency period | 7 days 1, 6 days 2, 5 days 3, 4 days 4 | |
| time taken for antibodies to develop | 14 days | |
| duration of vaccine-based immunity | 180 days | |
| duration of disease-based immunity | 180 days | |
| vaccine effectiveness | 0.85 2, 0.70 3, 0.45 4 |
| Parameter | Description | Sensitivity Index |
|---|---|---|
| vaccination parameter | ||
| transmission rate | 1 | |
| recovery rate | ||
| mortality rate of infectious people | ||
| birth rate | 1 | |
| natural mortality rate | ||
| latency rate | ||
| reinfection rate of recovered individuals | 0 | |
| antibody rate | ||
| reinfection rate of vaccinated individuals | 0.0407 |
| IV 1/RG 2 | −4% | −3% | −2% | −1% | +1% | +2% | +3% | +4% |
|---|---|---|---|---|---|---|---|---|
| A | 2.8757 | 2.1369 | 1.4116 | 6.9948 | 6.8726 | 1.3627 | 2.0267 | 2.6797 |
| 7.6732 | 5.7465 | 3.8256 | 1.9101 | 1.9051 | 3.8052 | 5.7007 | 7.5917 | |
| 2.1109 | 1.5800 | 1.0513 | 5.2469 | 5.2282 | 1.0439 | 1.5632 | 2.0810 | |
| 1.7604 | 1.3192 | 8.7878 | 4.3904 | 4.3834 | 8.7598 | 1.3129 | 1.7492 | |
| 1.7604 | 1.3192 | 8.7878 | 4.3904 | 4.3834 | 8.7598 | 1.3129 | 1.7492 | |
| 4.0000 | 3.0000 | 2.0000 | 1.0000 | 1.0000 | 2.0000 | 3.0000 | 4.0000 | |
| 8.5025 | 6.2780 | 4.1212 | 2.0294 | 1.9694 | 3.8811 | 5.7375 | 7.5406 | |
| 2.1812 | 1.6168 | 1.0655 | 5.2667 | 5.1491 | 1.0184 | 1.5109 | 1.9927 | |
| 4.1466 | 3.0837 | 2.0385 | 1.0108 | 9.9416 | 1.9721 | 2.9341 | 3.8806 | |
| 3.4809 | 2.5785 | 1.6980 | 8.3874 | 8.1893 | 1.6187 | 2.4000 | 3.1633 | |
| 6.6911 | 4.9657 | 3.2759 | 1.6210 | 1.5880 | 3.1438 | 4.6683 | 6.1622 |
| IV 1/RG 2 | −4% | −3% | −2% | −1% | +1% | +2% | +3% | +4% |
|---|---|---|---|---|---|---|---|---|
| A | 2.6254 | 1.9474 | 1.2842 | 6.3518 | 6.2181 | 1.2307 | 1.8270 | 2.4111 |
| 6.8446 | 5.1343 | 3.4235 | 1.7120 | 1.7126 | 3.4258 | 5.1396 | 6.8539 | |
| 1.8274 | 1.3712 | 9.1456 | 4.5749 | 4.5791 | 9.1624 | 1.3750 | 1.8342 | |
| 1.3908 | 1.0432 | 6.9554 | 3.4781 | 3.4789 | 6.9586 | 1.0439 | 1.3921 | |
| 1.3908 | 1.0432 | 6.9554 | 3.4781 | 3.4789 | 6.9586 | 1.0439 | 1.3921 | |
| 4.0000 | 3.0000 | 2.0000 | 1.0000 | 1.0000 | 2.0000 | 3.0000 | 4.0000 | |
| 8.5060 | 6.2805 | 4.1228 | 2.0302 | 1.9702 | 3.8827 | 5.7398 | 7.5436 | |
| 2.1824 | 1.6177 | 1.0661 | 5.2696 | 5.1519 | 1.0190 | 1.5118 | 1.9938 | |
| 1.3093 | 9.7225 | 6.4176 | 3.1773 | 3.1159 | 6.1718 | 9.1691 | 1.2109 | |
| 1.1444 | 8.4707 | 5.5739 | 2.7511 | 2.6817 | 5.2962 | 7.8456 | 1.0332 | |
| 1.5961 | 1.1823 | 7.7856 | 3.8454 | 3.7530 | 7.4162 | 1.0992 | 1.4483 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Margenov, S.; Popivanov, N.; Ugrinova, I.; Hristov, T. Differential and Time-Discrete SEIRS Models with Vaccination: Local Stability, Validation and Sensitivity Analysis Using Bulgarian COVID-19 Data. Mathematics 2023, 11, 2238. https://doi.org/10.3390/math11102238
Margenov S, Popivanov N, Ugrinova I, Hristov T. Differential and Time-Discrete SEIRS Models with Vaccination: Local Stability, Validation and Sensitivity Analysis Using Bulgarian COVID-19 Data. Mathematics. 2023; 11(10):2238. https://doi.org/10.3390/math11102238
Chicago/Turabian StyleMargenov, Svetozar, Nedyu Popivanov, Iva Ugrinova, and Tsvetan Hristov. 2023. "Differential and Time-Discrete SEIRS Models with Vaccination: Local Stability, Validation and Sensitivity Analysis Using Bulgarian COVID-19 Data" Mathematics 11, no. 10: 2238. https://doi.org/10.3390/math11102238