1. Introduction
A socially sound aim of the fishery is the sustainable provision of humankind with seafood. This subsumes the rational exploitation and preservation of the fish populations. A single responsible owner could ensure this. However, the catch is carried out by agents focused on their profit. Uncontrolled fishing by such agents can collapse, known as Tragedy of the Commons [
1,
2]. In the second half of the twentieth century, it was realized that the resources of the world ocean are finite, and overexploitation needs to be prevented [
3]. In particular, this resulted in the 1982 UN Convention on the Law of the Sea and the Exclusive Economic Zone (EEZ) regime. This and other measures have not led to a solution to the problem of fish stock depletion [
4,
5]. It is estimated that one-third of the world’s marine fish stocks are currently overfished [
6].
Many management strategies aim to avoid the negative consequences of fishing and achieve the mentioned socially significant government goal [
7,
8,
9,
10]. Among them are taxes, setting TAC (Total Allowable Catch), granting exclusive ownership to a single firm, forming a fishers’ cooperative, and establishing marine reserves [
11]. Furthermore, if the TAC is fixed, it can be allocated between the fishers through a mechanism of quotas. The most common form is ITQ (Individual Transferable Quotas), which first were implemented in New Zealand in 1986 [
12]. It would be naïve to assume that there is a simple tool that can solve a complex fishery problem. The main reason is that several factors are not included in any concrete model, and their impact can be quite substantial [
13] where ITQs are discussed.
Since many agents on the level of states and firms are involved in fisheries, it is natural that game theory came into play [
3,
14]. Although game theory can usually provide only qualitative results, its impact on management is significant. In particular, the report [
15] on the results of the Expert Consultation on the Management of Shared Fish Stocks (FAO 2002) reviews the basic concepts of game theory and discusses its applications.
In this paper, we consider a catch management method based on taxes. This method is more related to optimization than to the game theory. However, it is also connected with the Stackelberg leader-follower game. Using taxes is often mentioned in the economic literature [
16,
17,
18,
19]. Nevertheless, past research [
11] suggests that taxes have “seldom been attempted, at least not as the sole regulatory instrument.” In [
3], it is mentioned that presumably, tax schemes cannot be designed to foster intra-industry cooperation among the fishers. The role of taxes is underestimated. Indeed, taxation has solid mathematical grounds in optimization, where the resource prices are related to Lagrange multipliers [
20]. These prices can stimulate optimal cooperative behavior via a decentralized management scheme. This method is well known for flow control in communication networks [
21].
We distinguish two levels of management: the leader (e.g., a government) and the follower (agent, resource use). As a rule, agents pursue short-term commercial goals and do not think about the environmental consequences of their activities. On the contrary, the government wants to economically force the agent to act optimally in light of a long-term perspective. It usually implements a “hard” control to satisfy the constraints. This traditional way often spawns corruption.
Another approach subsumes a significant reduction of the role of the government. It brings to the fore specific “soft” purely economic mechanisms (e.g., taxes, etc.), making them responsible for the constraint compliance. This paper outlines the basics of this approach and shows its capabilities. It became evident that assigning a tax per unit of fish production equal to the derivative of the value function (corresponding to the specified optimal yield problem) forces the fisher to adhere to the optimal catching strategy with respect to the infinite horizon. In this case, the population size stabilizes at the universal limiting value, not depending on the utility function. These ideas were expressed in [
22].
Indeed, it is not easy to determine the correct taxes. First, they depend on the dynamics of the fish population, which can only be modelled approximately. Moreover, the real stock of fish may be unknown. Second, optimal taxes depend on the agent reward functions, which are even more difficult to model. Third, as already mentioned, there are many factors that cannot be taken into account within a single model. For these reasons, only qualitative investigations in fishery management prevail at this time. Even prior studies [
23,
24,
25] using real data mainly result in qualitative conclusions. Thus, we are doubtful that the correct real-world tax can be obtained within the simple models under consideration. Furthermore, the traditional methods of fish stock evaluation based on random samples seem not to be entirely reliable. We believe that Artificial Intelligence will be able to make substantial progress in quantitative results. Comprehensive fisheries datasets have been collected. As is mentioned in [
26], the dataset [
27] contains information about 1433 stocks belonging to 387 unique species.
The aim of our research is to study the method of long-term optimal control of natural resources based on taxation. The proposed taxation mechanism is based on internal prices, settled by a responsible resource owner (government) to ensure sustainable development. The internal prices reduce a complex problem of optimal long-term exploitation to a sequence of one-year optimization problems and stimulate a cooperative behavior of competing agents. We also consider evolutionary and spatial aspects of this problem.
2. Methods
The concept of a price of a product (good) can be defined in different ways. In optimization theory, one first finds the maximum of a possible gain that this product can
bring. Here,
is the initial stock of a product, while
defines the gain from the stock.
is defined as an indirect utility. Then it is usually assumed [
28] that the price
equals the marginal gain:
We call this price “internal.” In a two-dimensional case the prices of goods are determined by partial derivatives:
As an example, consider the problem of optimal long-term fishing described by a simple discrete model
where
and
are the current amount and annual catch of fish
We assume that the annual reward is given by a utility function
. For an illustration, we consider the simplest class of nonlinear ecology models, where
are strictly increasing strictly concave smooth functions, satisfying the condition
. For instance,
and
. The condition
implies the representation
for a smooth function
. Moreover, the concavity of
implies that
is decreasing. Sometimes
is called a trophic function, which reflects a negative impact of the inter- and intra-specific competition. A useful collection of modern population growth functions and their discussion are presented in [
29].
Let us first discuss the simplest model of population dynamics without fishing. On the one hand, to prevent the variables of this model from going to infinity, one should require a slow population growth, for instance,
, for all sufficiently large
(say, for
, where
is a constant). On the other hand, it is necessary to assume that
, otherwise the population will “die by itself.” Under these conditions, there exists a unique positive equilibrium. For convenience, we also select a scale such that
For an initial population stock
the global profit is the supremum of the discounted sum of rewards over all admissible sequence of catches:
From an economic point of view, the discounting reflects the lesser significance of the future income compared to the past one. From a mathematical point of view, it ensures that the sum is finite.
We will consider the so-called “highly productive” populations with
. In this case, the optimal catch satisfies the strict inequality
for all
[
11]. For low-productive populations, the equality
is possible for large
. Computation of
is carried out according to well-known dynamic programming schemes [
30]. Recall that the initial piece
of the optimal sum (2) from
to
satisfies the recursion:
with
. By induction in
n, we easily obtain (see, e.g., [
22,
31]):
Property 1. Eachis a strictly increasing concave function satisfying the condition.
It is natural to consider these functions as elements of the Banach space of continuous functions with the sup-norm. The right-hand side of Equation (3) maps this space into itself.
Property 2. The mapping determined by the right-hand side of Equation (3) is a contraction with the coefficient γ.
Hence, the sequence of functions
converges uniformly to a monotone concave value function
. It was also shown that this function is continuously differentiable. We set the price of a fish unit equal to
. From the strict concavity of at least one of the functions
or
, it follows that
is strictly decreasing. Of course, for other classes of models, the concavity property of
is rare, but its monotonicity often takes place. According to Lebesgue’s theorem [
32], a monotone function is differentiable almost everywhere. Therefore, in this case, we also can talk about the price of a fish (or a resource) for “almost all”
.
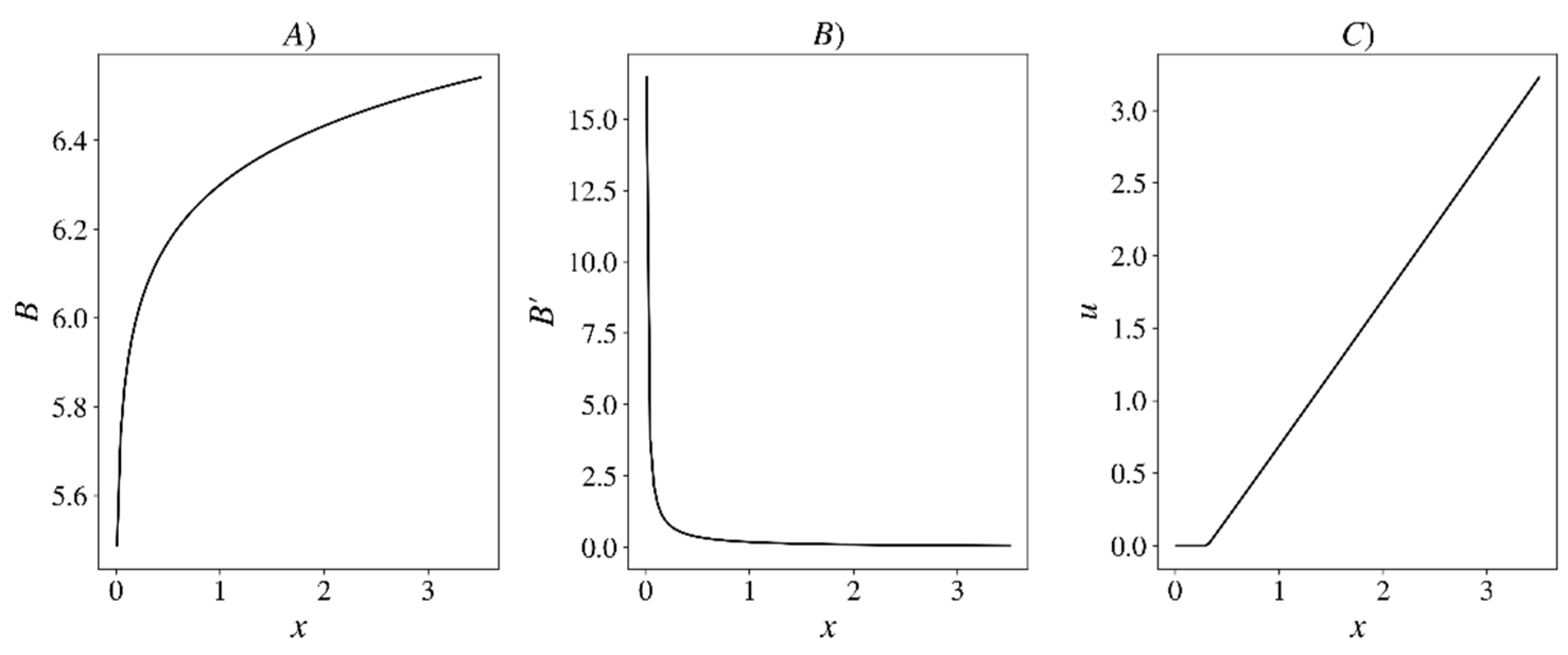
Let us mention some other useful characteristics of the fishery (
Figure 1). Their justification is given in the monograph [
19].
Property 3. Optimal fishingis a continuous and non-decreasing function. For small, we have, and for large, it strictly increases.
This statement is, to some extent, consistent with the concept of minimum possible population size [
33]. Indeed, in the infinite series (2), the optimal catch
depends on
only through the state variable
According to Property 3,
for small
. Therefore
. Taking the derivative, we obtain
. By assumption, the factor in square brackets is greater than 1. It follows that
Using the next property, one can prove that the dynamical system (Equation (1)) with
has a unique positive globally stable equilibrium [
22].
Property 4. The function x-u(x) is strictly increasing.
Indeed, this property implies that the right side of Equation (1) is increasing.
In the absence of fishing, population dynamics obey the simplified version of Equation (1), namely
In some popular models,
can be a unimodal trophic function: recall Ricker’s “humpy” function
[
34]. Complex aperiodic regimes can arise in this model for a large constant
. Somewhat surprisingly, optimal fishing has a stabilizing effect here. The following result was proven in [
19].
Property 5. For any parameterin the Ricker model with fishing, each trajectory tends to some fixed equilibrium.
The next property is of key importance (see
Appendix A for a proof).
Property 6. Letat some point x, then In other words, the price generated by the “global utility function” must match the price from the “local utility function.” Note that Equation (4), the concavity of
and the inequality
implies that:
Now the complex search for the optimal harvest vector in the multi-year problem is reduced to a simple one-year optimization problem. Using Equation (4), it is easy to prove the following result (see
Appendix A).
Property 7. Each globally optimal catchis an optimal solution to the elementary optimization problem Therefore, it is natural to interpret the internal price as a tax per unit of fish caught.
Now, let us consider a slightly more general case when there are
agents (fishers). The dynamics
of a population, exploited by
agents are described by the Equation
where
and
. If the agents are far-sighted and act cooperatively, then their common objective function has the form
Let
be the correspondent value function. To stimulate such cooperative behavior, the leader can settle the universal (the same for all agents) price
per unit fish. The next result was proven in [
31].
Theorem 1. The solutions of the problemsfor allcoincide with the far-sighted cooperative optimal strategies.
This subsumes the assertion that the optimal myopic solutions automatically satisfy the constraint Note that in the absence of a catch tax, each agent will strive to catch all the fish. In addition, when , a conflict situation arises between the agents, and they must agree on quotas. Theorem 1 shows that the tax eliminates these problems. The next result shows that the competition problem remains, although it takes a hidden form.
Theorem 2. The tax increases with the number of agents:for k>m. Moreover, a strict inequality holds everywhere, except maybe a nowhere dense closed set.
This implies that with an increase in the number of agents, the tax on catch increases and, accordingly, the optimal volumes of catch and profits of all agents decrease. Therefore, the fisher community will resist the entrance of new members. The reason for increasing the tax with an increase in the number of agents is economically clear. The continuous time case was considered in [
35]. Technically, this case is more complicated, and the formulation of the results requires some modification. However, some analogs of Theorems 1 and 2 remain valid.
Above we considered traditional one-region population models. Now, we turn to the case of two regions to analyze harvesting in the case of spatial population dynamics using the ideas of an evolutionary ecology within some novel models. It is important to mention the ambiguity of the outcomes of the evolutionary process in the framework of the models under consideration. In [
36], when searching for migration routes (described by matrices) depending on the spatial distribution of food resources, it was found that the final matrix depends on the choice of the initial matrix. However, unexpectedly it turned out that all such final matrices have, in fact, the same positive eigenvector. This (Perron) vector turns out to be unique [
37]. Each of its components is the relative time spent by the population in the
-th area. Therefore, any adaptive behavior is reduced to the restructuring of the sojourn times. Hence, the mentioned matrix is only one of many
forms of a particular migration, and the true
content of the migration is the Perron vector. The solution of the optimal harvesting problem can be supplemented with an economic filling based on the idea of internal prices of the fish population for different areas. This allows to set effective economic mechanisms: taxation, trade exchanges, etc.
Let the reservoir be divided, for example, into two regions (1 and 2), in each of which a catch is performed. It is assumed that the fish population migrates between these areas. Formally, let
and
be the current fish stocks of a given population in regions 1 and 2, respectively. Then its displacement per unit of time is given by a linear model:
where
. Since the total population size does not change during “pure” migration, the sum of the elements in each column is equal to 1. Such matrices are called Markov. It is convenient to represent two-dimensional Markov matrices in the following form:
where
. Off-diagonal elements
and
characterize the intensity of population movement. Thus,
is the fraction of the population that can move from region 2 to region 1. Similarly, α defines the fraction of the population moving from the first region to the second region. It is convenient to represent a matrix
as a point
in a plane. Together, they fill the square
with the vertices
. Each point of
is a convex combination of these vertices, which, in fact, constitute a “convex” basis in
. Let us construct the corresponding “basis” matrices:
Let us discuss the adaptation of the migration route from the point of view of evolutionary ecology. Assume that the original population with the migration matrix
can generate “mutants” with routes
close to
. Namely:
where
is small and
is one of the basis matrices. In the proposed modeling scheme, the route of the mutant will change from time to time. Therefore, to complete the picture, one should specify the order in which the basis matrices are generated. The number of mutants in the model is specified in advance. Of course, the smallest number of them is 1. It has been established that with an increase in the number of mutants, the result does not change much.
Here the process of competition of the initial population (numbered by
) with its community of mutants (numbered by
) is of key importance. Assume that
-th population is described by the equation
,
. If these populations are “similar” (for example, they are mutants of the original population), then their competition is given in the form [
19]:
, where
We will use this approach for the construction of evolution mechanisms.
Consider the “basic example”, which describes the interaction of the initial population
with a single mutant
. In the sequel for the specified population vectors, we will use the lower symbol for the indication of the time moment. We assume that in the
-th region, the trophic function for each population is the same and equals to
. The routes of their movement are specified given by the matrices:
The model of joint competition and migration is a composition of two operators
and
. As an illustration, we present formulas for the transformation of the vector
under
:
and formulas for its transformation under
:
We assumed that the initial amount of population is about 1, while the initial mutant amounts are small. The computations were carried in several stages. Each stage corresponded to a 100-year forecast, which is a significant time in terms of evolution. The results were summarized at the end of each stage. We note right away that the competition outcome depends little on the choice of initial population sizes.
Most importantly, at the end of a stage, a “strong” mutant can be revealed, whose amount can significantly exceed the amount of the original population (for example, by 10 or more times). If there are several strong mutants, then we choose the strongest. Furthermore, we assume that such a mutant sets the direction of natural selection. Namely, let
be the migration matrix of a strong mutant. Then we deform the matrix of the original population as follows:
where the adaptation rate parameter
is small. If no strong mutants are found, then
it is saved.
After that comes the second stage. We act similarly since for each mutant a new route (specified in Equation (12)) is formed with another . In the end, a stage comes where strong mutants are no longer formed. This means that the final matrix is evolutionarily stable (ES-matrix) since it is not displaced by its mutants with close migration routes. Of course, only ES migration matrices can be realized in nature. In the extreme version of this concept, the number of mutants can be arbitrary. We used only two mutants such that the enumeration of the basis matrices occurs in opposite directions (i.e., clockwise: and counterclockwise).
For example, consider an applied problem of changing the routes of fish migration depending on the ratio of region food resources under optimal fishing conditions. For example, assume that if
is the total harvest in the region 1, then its distribution over specific populations are uniform. Namely, the catch operator
is defined there as follows:
Similar formulas hold true for the components , in the region 2. It is interesting to what extent do the obtained results depend on the number of generated mutants. Preliminary computations showed that they are preserved even for large number of mutants.
Next, we introduce the parameter , which characterizes the share of the 1-region in the reservoir. Denote by the total amount of food in the reservoir. Then is the amount of food in the first region, and is the amount of food in region 2. We will discuss the simplest case, i.e., when food supplies are fixed and do not change in each region.
The optimization block of the model considers harvesting in both areas for a multi-year period. Denote by
the optimal discounted income for
years with initial fish stocks equal to
(in the first region) and
(in the second region). During the exploitation, the amount of fish changes. We denote the corresponding trajectory by
. By definition,
where the maximization is performed over all admissible catches
and
in both regions:
. As above,
is the discount factor (put
). Furthermore,
are concave increasing utility functions with
. We put
. Note that
.
We represent any
-population dynamics as
We will use the simplest variant of the non-linear growth, assuming that
is smooth, strictly increasing, and concave in the first argument. These assumptions are satisfied by the scheme from [
38]:
where
and
are the mortality and growth rates. Put
,
for brevity. Now, taking into account the annual fishing:
(in the first region) and
(in the second region), the dynamics of one population takes the form:
Rewrite Equations (21) and (22) as
. Note that
are strictly increasing and concave in each variable. By the dynamic programming principle:
where the maximum is taken over
Evidently,
. Each function
is continuous. Two-dimensional analogs of the previous properties still hold true.
Furthermore, B inherits the “good” properties of the utility and growth functions. It is strictly increasing and concave in each variable. Thus, the partial derivatives , exist almost everywhere, and can be regarded as an indirect utility function of the initial fish stocks by area equal to and . Its partial derivatives and can be interpreted as internal fish prices in the first and second regions, respectively.
Statement 1. Let the optimal catchatbe positive, then there existsand Statement 2. Let the optimal catchatbe positive, then there existsand.
The quantity, e.g.,
can be used as a tax per unit of fish caught in the first region. Let us discuss the solution of the following simple optimization problem for the fisher of the first region:
To find out how much the controls differ in Equations (6) and (25), let us consider two cases for Equation (25). Let , then the objective function is strictly decreasing since its derivative is negative: . It follows that is optimal. Now assume that . Then by the strict concavity of the desired control satisfies the Equation , that is . Clearly, this relation coincides with Equation (24). Similarly, can be used as an effective catch tax in the second region. Thus, the solution of the multi-step optimization problem (Equation (12)) reduces to the solution of two one-step optimization problems in the form of Equation (25).
3. Results
We will use the following global characteristics where is the average asymptotic value of a sufficiently representative bundle of trajectories of the model given by Equations (21) and (22). The search for an optimal harvest strategy and the construction of the accompanying ES migration matrices includes two successive stages (blocks). The first stage (block) is optimization. For a given migration matrix a search for optimal catches (functions ) is performed using the dynamic programming method. This corresponds to a numerically implemented mapping . The second stage (block) is evolution. For fixed catch functions a competitive interaction of the initial population (with the migration matrix with several mutants whose matrices are close to , is realized (Equation (12)). When a strong mutant is detected as a result of selection, we perform some deformation of (Equation (17)). Formally, a mapping is implemented numerically.
If, after multiple repetitions of stages 1 and 2, this process converges, then a stable evolutionary matrix
and optimal catches
are determined for a given value of food in areas 1 and 2. In each case, the corresponding computations were carried out for initial values varying on a certain grid. Let
be the total supply of food in the reservoir, and the amounts of food
and
for each of the two regions are given by a parameter
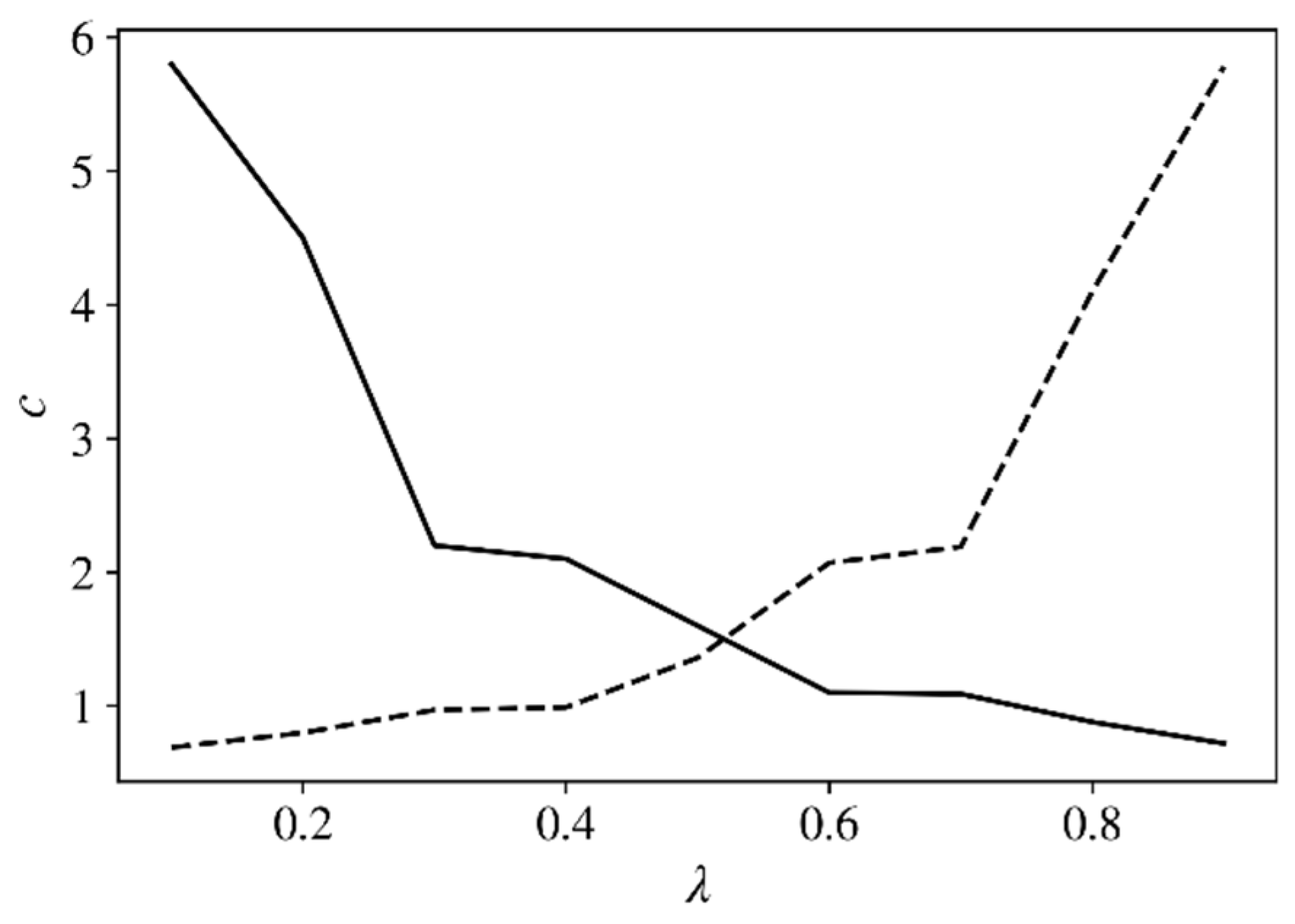
from the segment [0.1, 0.9]. The dynamics of the internal prices for various values of
are presented in
Figure 2.
In the first area, as expected, it turned out that the average number and catch of fish increased with an increase in the parameter
due to the growth of food there. The opposite changes occur in the second region. Of course, the smaller the amount of fish, the higher its price. According to obtained results, as
increases, the Perron vector
runs through the values:
Finally, if, for example
, then an additional benefit is possible from the sale of cheap fish from the second region to the owner of the first region. Harvesting can be considered an evolutionary game of “giveaway.” The influence of harvesting on the evolution of the fish migration route can be used to optimize the fishing of one population in two neighboring states A (region 1) and B (region 2) within the same reservoir. In fact, these countries compete for fish resources. What are ecological and economic mechanisms effective here? One of the paradoxical approaches is covered in [
39]. Below, we present this in more detail. We will proceed in the spirit of hierarchical management, and at the same time, we will somewhat specify the previous agreements on fishing strategies. Namely, we assume that the Center, based on the solution of the problem given in Equation (19), issues recommendations to these states on the catch quota, which should not exceed the optimal values
and
, and depending on
. Of course, in some years, it is possible to catch even less (liberal fishing strategy). Assume that the initial migration matrix is of the form:
and the food resources of the water areas of these countries are the same. It is convenient to use chess terminology.
First, consider an equilibrium “innocuous” situation. If the countries follow the Leader’s recommendations (taking into account
and the solution of Equation (12)), then the migration matrix is preserved, and the following average values arise:
;
. Clearly, for
we have
In the second stage,
A loses material but gains a positional advantage. Let country
A harvest 75% of the recommended amount, and country
B harvest 100%. Then the
A-water area becomes “less lethal” and, therefore, more attractive for fish. There is a “luring” of the fish population into the
A-water area. As a result of evolution, the migration matrix is transformed to:
The correspondent average values:
;
;
. Clearly, for
we have:
In fact, A played a “giveaway” with B here, but gained a positional advantage by constructing a more profitable migration matrix.
In the third stage, A realizes positional advantage and obtains a material advantage. Assume that considering the matrix, a coordinating center issues new recommendations for the optimal catch, which are now 100% implemented by both participants. This results in ; ; . It is remarkable that if now both countries and adhere to this catch, then the migration matrix is no longer “spoiled.” Therefore, it is evolutionarily stable under the indicated controls. Most importantly, after the “luring” procedure, the catch in country A will always be greater than in country B. Since , A has the opportunity for additional trade benefits by selling its “cheap” fish to B. To restore the status quo, country B must likely apply the “lure” procedure in turn. An endless series of -lures and -lures occurs.


{kind=link}
{kind=link}

