Intraspecific Variations in Functional and Molecular Traits of Near-Endemic Onopordum alexandrinum Boiss. in Natural and Anthropogenic Habitats along the Western Mediterranean Coast of Egypt: Implications for Conservation


Abstract
:1. Introduction
2. Results
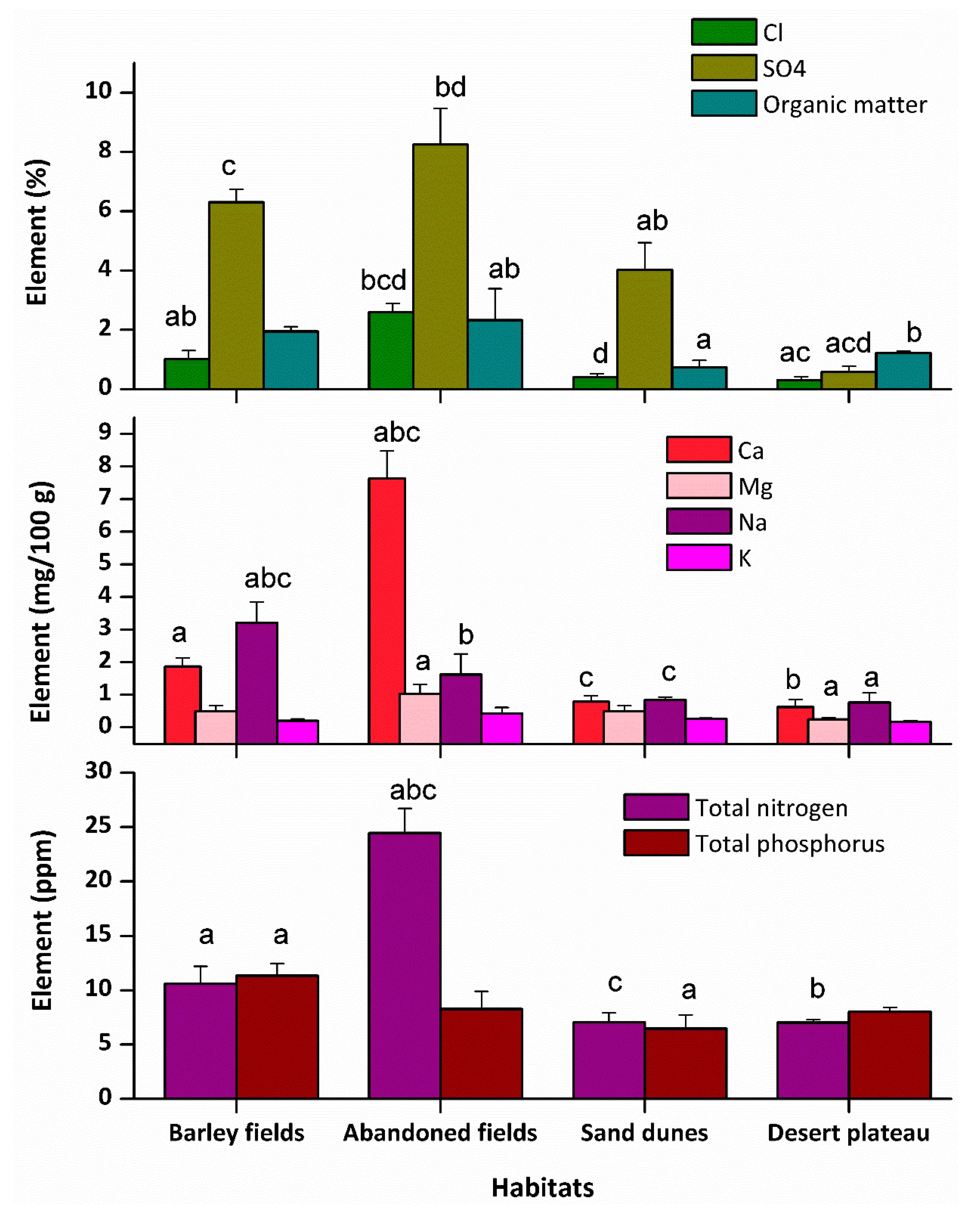
2.1. Soil Properties
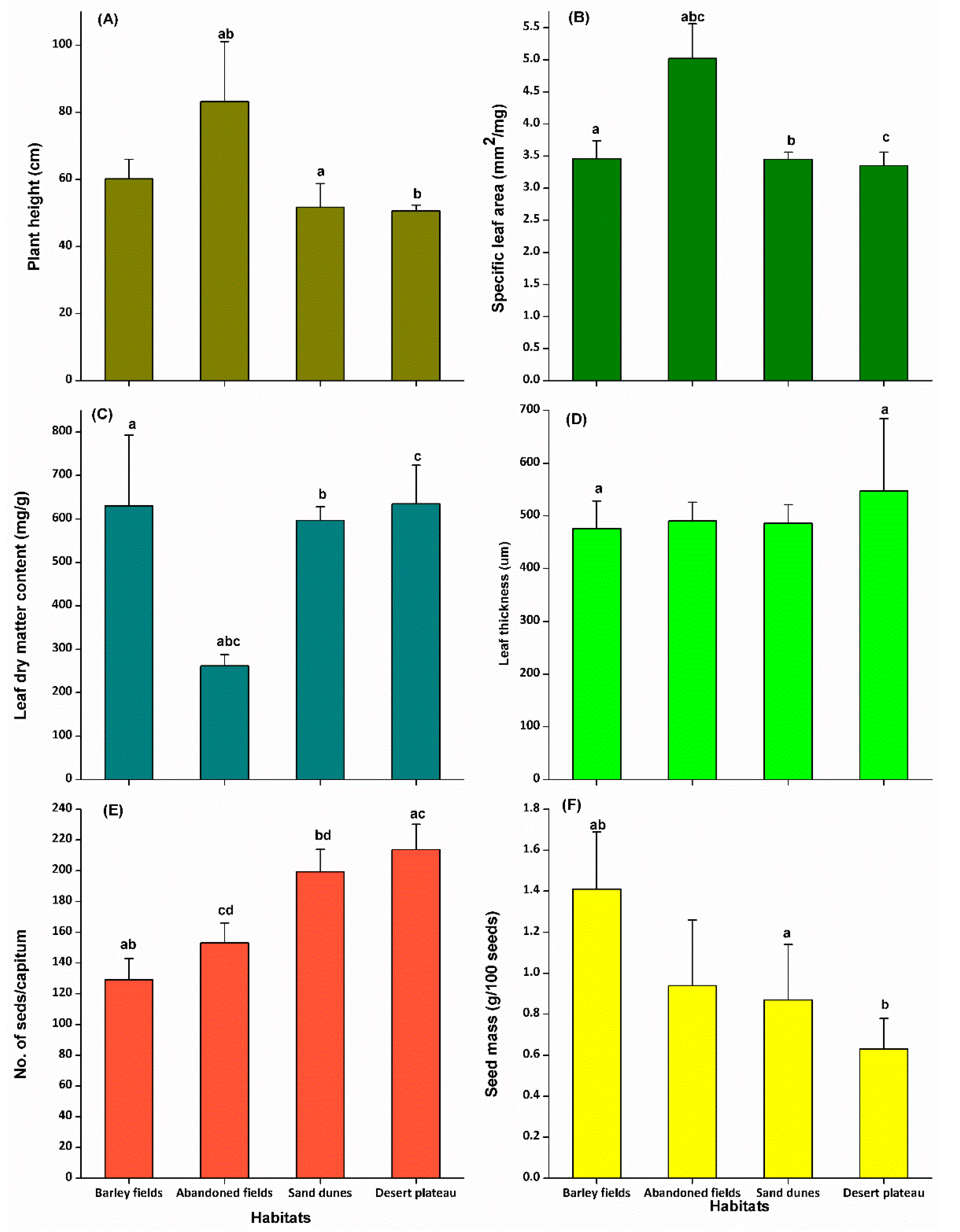
2.2. Functional Traits
2.3. Trait–Soil Correlation
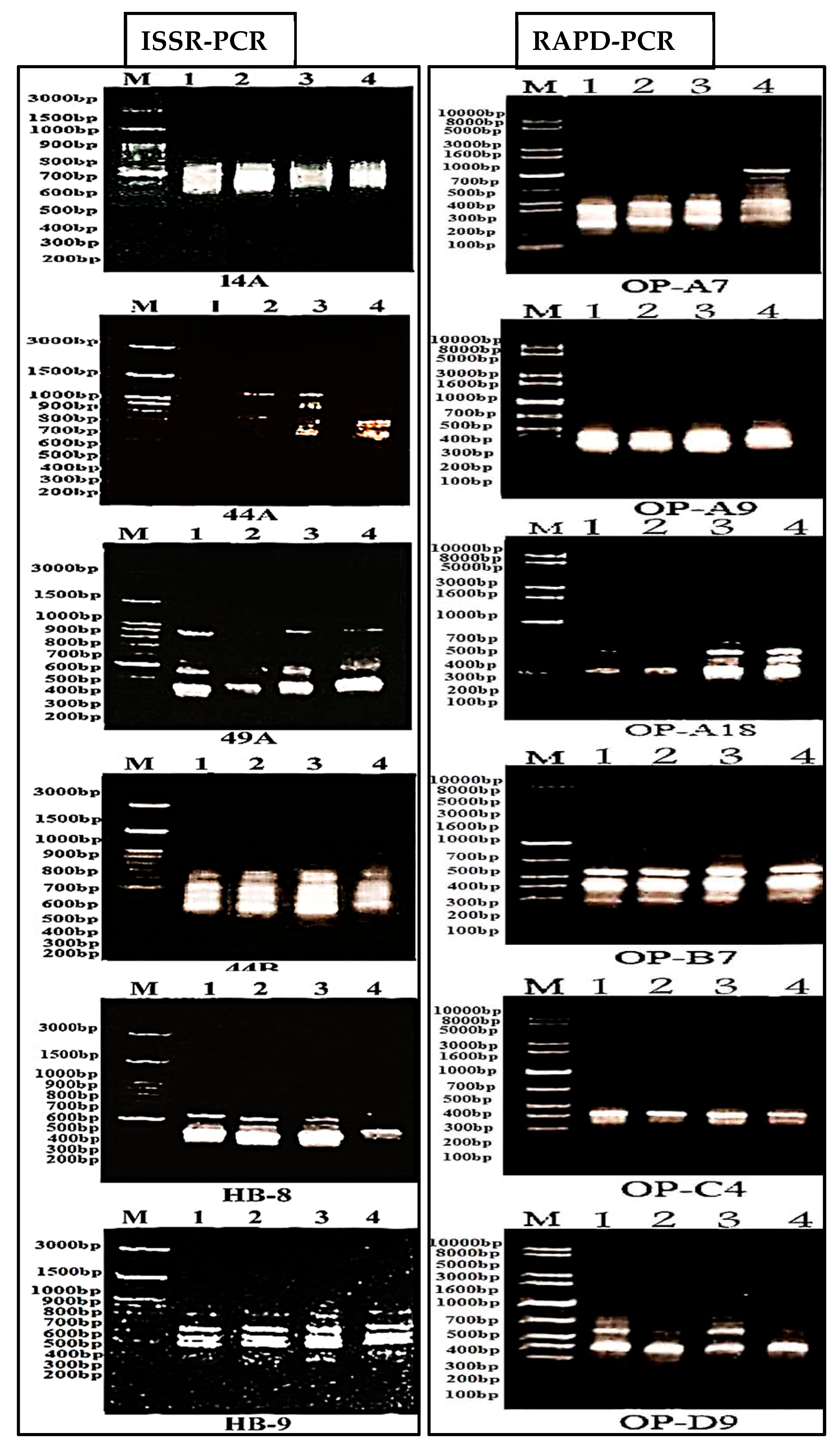
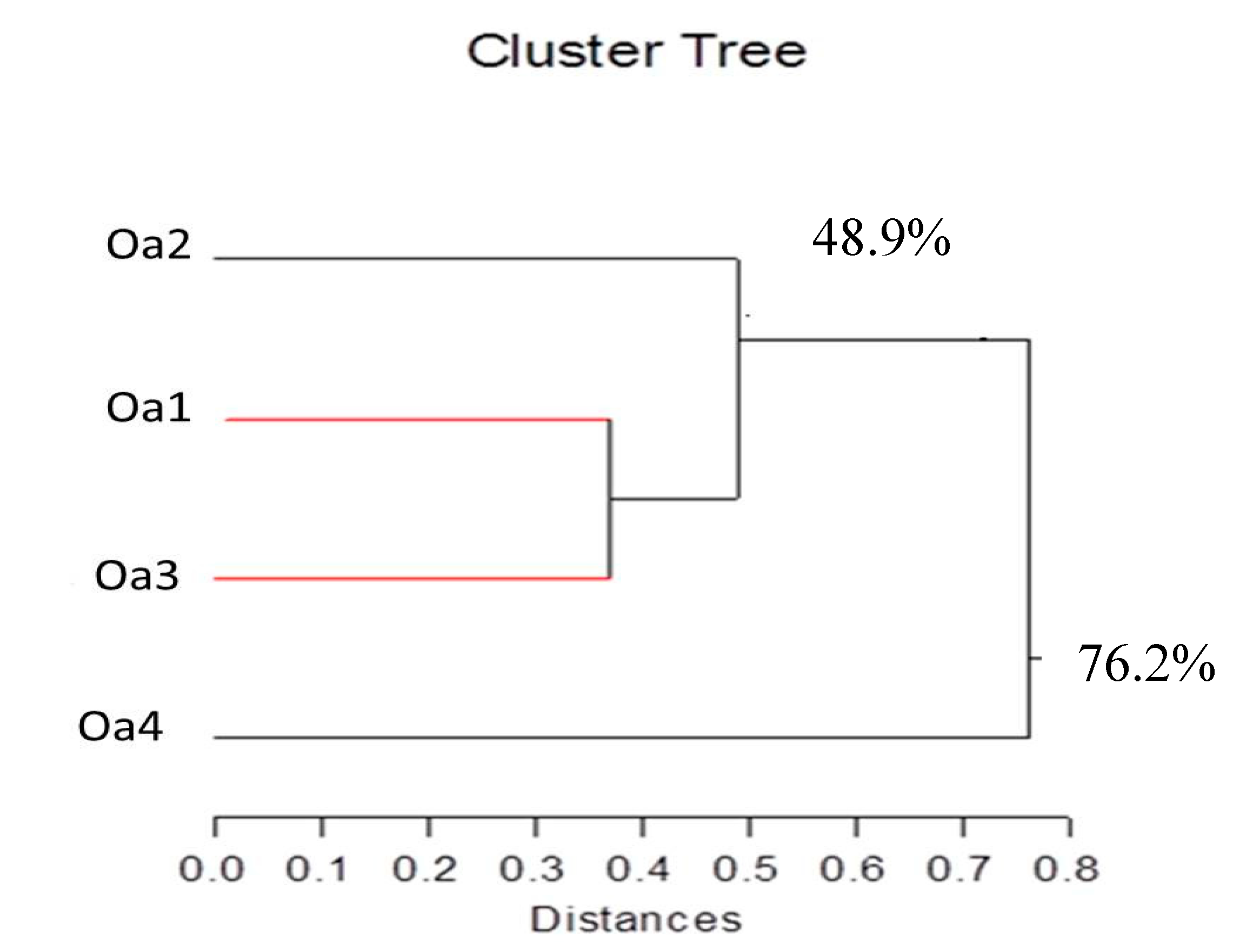
2.4. Molecular Analysis
3. Discussion
Implications for Conservation
4. Materials and Methods
4.1. Study Area and Habitat Selection
4.2. Soil Analysis
4.3. Functional Traits
4.4. Molecular Traits
4.4.1. Sample Collection and DNA Extraction
4.4.2. Random Amplified Polymorphic DNA (RAPD) Amplification
4.4.3. Inter Simple Sequence Repeat (ISSR) Amplification
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Diáz, S.; Cabido, M. Plant functional types and ecosystem function in relation to global change. J. Veg. Sci. 1997, 8, 463–474. [Google Scholar] [CrossRef]
- Ackerly, D.D.; Cornwell, W.K. A trait-based approach to community assembly: Partitioning of species trait values into within- and among-community components. Ecol. Lett. 2007, 10, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Ceulemans, T.; Merck, R.; Hens, M.; Honnay, O. A trait-based analysis of the role of phosphorus vs nitrogen enrichment in plant species loss across North-West European grasslands. J. Appl. Ecol. 2011, 48, 1155–1163. [Google Scholar] [CrossRef]
- Navas, M.L. Trait-based approaches to unravelling the assembly of weed communities and their impact on agro-ecosystem functioning. Weed Res. 2012, 52, 479–488. [Google Scholar] [CrossRef]
- Leimu, R.; Mutikainen, P.; Koricheva, J.; Fischer, M. How general are positive relationships between plant population size, fitness and genetic variation? J. Ecol. 2006, 94, 942–952. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Neel, M.C.; Ellstrand, N. Conservation of genetic diversity in the endangered plant Eriogonum ovalifolium var. vineum (Polygonaceae). Conserv. Genet. 2003, 4, 337–352. [Google Scholar] [CrossRef]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Kirk, H.; Freeland, J.R. Applications and implications of neutral versus non-neutral markers in molecular ecology. Int. J. Mol. Sci. 2011, 12, 3966–3988. [Google Scholar] [CrossRef] [Green Version]
- Geert, H.J.; Wynter, D.T.; Perkins, M.; Machairopoulou, M.; Neill, S.P. Context dependency of relationships between biodiversity and ecosystem functioning is different for multiple ecosystem functions. Oikos 2009, 118, 1892–1900. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; Lamarque, P.; Colace, M.P.; Garden, D.; Girel, J.; Pellet, G.; Douzet, R. Using plant functional traits to understand the landscape distribution of multiple ecosystem services. J. Ecol. 2011, 99, 135–147. [Google Scholar] [CrossRef]
- Boyer, K.E.; Kertesz, J.S.; Bruno, J.F. Biodiversity effects on productivity and stability of marine macroalgal communities: The role of environmental context. Oikos 2009, 118, 1062–1072. [Google Scholar] [CrossRef]
- Ilves, A.; Lanno, K.; Sammul, M.; Tali, K. Genetic variability, population size and reproduction potential in Ligularia sibirica (L.) populations in Estonia. Conserv. Genet. 2013, 14, 661–669. [Google Scholar] [CrossRef]
- Hajek, P.; Kurjak, D.; von Wühlisch, G.; Delzon, S.; Schuldt, B. Intraspecific variation in wood anatomical, hydraulic, and foliar traits in ten European beech provenances differing in growth yield. Front. Plant Sci. 2016, 7, 791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, P.B.; Wright, I.J.; Cavender-Bares, J.; Craine, J.M.; Oleksyn, J.; Westoby, M. The evolution of plant functional variation: Traits, spectra, and strategies. Int. J. Plant Sci. 2003, 164, 143–164. [Google Scholar] [CrossRef]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.G.; Douzet, R.; Aubert, S.; Lavorel, S. A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits. Funct. Ecol. 2010, 24, 1192–1201. [Google Scholar] [CrossRef]
- Schaal, B.A.; Leverich, W.J.; Rogstad, S.H. Comparison of methods for assessing genetic variation in plant conservation biology. In Genetics and Conservation of Rare Plants; Falk, D.A., Holsinger, K.E., Eds.; Oxford University Press: New York, NY, USA, 1991; pp. 123–134. [Google Scholar]
- Siefert, A.; Violle, C.; Chalmandrier, L.; Albert, C.H.; Taudiere, A.; Fajardo, A.; Aarssen, L.W.; Baraloto, C.; Carlucci, M.B.; Cianciaruso, M.V. A global meta-analysis of the relative extent of intraspecific trait variation in plant communities. Ecol. Lett. 2015, 18, 1406–1419. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Jung, V.; Violle, C.; Mondy, C.; Hoffmann, L.; Muller, S. Intraspecific variability and trait-based community assembly. J. Ecol. 2010, 98, 1134–1140. [Google Scholar] [CrossRef]
- Long, W.; Zang, R.; Schamp, B.S.; Ding, Y. Within- and among-species variation in specific leaf area drive community assembly in a tropical cloud forest. Oecologia 2011, 167, 1103–1113. [Google Scholar] [CrossRef]
- Pontes, D.S.L.; Soussana, J.F.; Louault, F.; Aandueza, D.; Carreìre, P. Leaf traits affect the above-ground productivity and quality of pasture grasses. Funct. Ecol. 2007, 21, 844–853. [Google Scholar] [CrossRef]
- Lecerf, A.; Chauvet, E. Intraspecific variability in leaf traits strongly affects alder leaf decomposition in a stream. Basic Appl. Ecol. 2008, 9, 598–605. [Google Scholar] [CrossRef] [Green Version]
- Miner, B.G.; Sultan, S.E.; Morgan, S.G.; Padilla, D.K.; Relyea, R.A. Ecological consequences of phenotypic plasticity. Trends Ecol. Evol. 2005, 20, 685–692. [Google Scholar] [CrossRef] [Green Version]
- Bossdorf, O.; Richards, C.L.; Pigliucci, M. Epigenetics for ecologists. Ecol. Lett. 2008, 11, 106–115. [Google Scholar] [CrossRef]
- Violle, C.; Enquist, B.J.; McGill, B.J.; Jiang, L.; Albert, C.H.; Hulshof, C. The return of the variance: Intraspecific variability in community ecology. Trends Ecol. Evol. 2012, 27, 244–252. [Google Scholar] [CrossRef]
- Vijayan, K. Inter Simple Sequence Repeat (ISSR) polymorphism and its application in mulberry genome analysis. Int. J. Indust. Entomol. 2005, 10, 79–86. [Google Scholar]
- Powell, W.; Morgante, M.; Andre, C. The comparison of RFLP, RAPD, AFLP and ISSR (microsatellite) marker for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Russell, J.R.; Fuller, J.D.; Macaulay, M.; Hatz, B.G.; Jahoor, A.; Powell, W.; Waugh, R. Direct comparison of levels of genetic variation among barley accessions detected by RFLPs, AFLPs, SSRs and RAPDs. Theor. Appl. Genet. 1997, 95, 714–722. [Google Scholar] [CrossRef]
- Joshi, C.P.; Nguyen, H.T. RAPD (Random amplified polymorphic DNA) analysis based on inter varietal genetic relationships among hexaploid wheats. Plant Sci. 1993, 93, 95–103. [Google Scholar] [CrossRef]
- Heikal, Y.M. Genetic Diversity Assessment of Genus Mesembryanthemum Population (Aizoaceae) in EGYPT. Ph.D. Thesis, Botany Department, Mansoura University, Mansoura, Egypt, 2015. [Google Scholar]
- Shaltout, K.H.; Hosni, H.A.; El-Fahar, R.A.; Ahmed, D.A. Flora and vegetation of the different habitats of the western Mediterranean region of Egypt. Taeckholmia 2015, 35, 45–76. [Google Scholar]
- Shaltout, K.H.; Ahmed, D.A.; Diab, M.; El-Khalafy, M. Re-assessment of near-enedmic taxa in Egyptian flora. Taeckholmia 2018, 38, 61–83. [Google Scholar] [CrossRef]
- Abdelaal, M.; Ahmed, D.; Fois, M.; Fenu, G.; Bacchetta, G. Floristic patterns and ecological drivers of sand dune ecosystem along the Mediterranean coast of Egypt. Arid Land Res. Manag. 2019, 33, 388–411. [Google Scholar] [CrossRef]
- Radford, E.A.; Catullo, G.; de Montmollin, B. Important Plant Areas of the South and East Mediterranean Region: Priority Sites for Conservation; IUCN: Gland, Switzerland; Malaga, Spain, 2011; pp. 1–104. [Google Scholar]
- Wilson, P.J.; Thompson, K.; Hodgson, J.G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 1999, 143, 155–162. [Google Scholar] [CrossRef]
- Al Haj Khaled, R.; Duru, M.; Theau, J.P.; Plantureux, S.; Cruz, P. Variation in leaf traits through seasons and N-availability levels and its consequences for ranking grassland species. J. Veg. Sci. 2005, 16, 391–398. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Westoby, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Cornelissen, J.H.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.; et al. A handbook of protocols for standardized and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Lind, E.M.; Borer, E.; Seabloom, E.; Adler, P.; Bakker, J.D.; Blumenthal, D.M.; Crawley, M.; Davies, K.; Firn, J.; Gruner, D.S.; et al. Life-history constraints in grassland plant species: A growth-defense trade-off is the norm. Ecol. Lett. 2013, 16, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B. The worldwide ‘‘fast-slow’’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA trait base: A database of life-history traits of Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Ryser, P.; Urbas, P. Ecological significance of leaf life span among Central European grass species. Oikos 2000, 91, 41–50. [Google Scholar] [CrossRef]
- Gross, N.; Suding, K.N.; Lavorel, S. Leaf dry matter content and lateral spread predict response to land use change for six subalpine grassland species. J. Veg. Sci. 2007, 18, 289–300. [Google Scholar] [CrossRef]
- Grime, J.P. The role of plasticity in exploiting environmental heterogeneity. In Exploitation of Environmental Heterogeneity in Plants; Caldwell, M.M., Pearcy, R., Eds.; Academic press: San Diego, CA, USA, 1994; pp. 1–18. [Google Scholar]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef] [Green Version]
- Kühner, A.; Kleyer, M. A parsimonious combination of functional traits predicting plant response to disturbance and soil fertility. J. Veg. Sci. 2008, 19, 681–692. [Google Scholar] [CrossRef]
- Swenson, N.G. The functional ecology and diversity of tropical tree assemblages through space and time: From local to regional and from traits to transcriptomes. Int. Sch. Res. Not. 2012, 1–16. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Seedling survival and seed size: A synthesis of the literature. J. Ecol. 2004, 92, 372–383. [Google Scholar] [CrossRef]
- Leishman, M. Does the seed size/number trade-off model determine plant community structure? An assessment of the model mechanisms and their generality. Oikos 2001, 93, 294–302. [Google Scholar] [CrossRef]
- Kitajima, K.; Myers, J.A. Seedling ecophysiology: Strategies toward achievement of positive net carbon balance. In Seedling Ecology and Evolution; Leck, M.A., Parker, V.T., Simpson, R.L., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 172–188. [Google Scholar]
- Yang, W.P.; Oiiverira, A.C.; Godwin, I. Comparison of DNA marker technologies in characterizing plant genome diversity: Variability in Chinese sorghums. Crop Sci. 1996, 36, 1669–1676. [Google Scholar] [CrossRef]
- Gilbert, J.E.; Lewis, R.V.; Wilkinson, M.J. Developing an appropriate strategy to assess genetic variability in plant germplasm collections. Theor. Appl. Genet. 1999, 98, 1125–1131. [Google Scholar] [CrossRef]
- Qian, W.; Ge, S.; Hong, D.Y. Assessment of genetic variation of Oryza granulata detected by RAPDs and ISSRs. Acta Bot. Sin. 2000, 42, 741–750. [Google Scholar]
- Levi, A.; Thomas, C.E.; Simmons, A.M.; Thies, J.A. Analysis based on RAPD and ISSR markers reveals closer similarities among Citrullus and Cucumis species than with Praecitrullus fistulosus (Stocks) Pangalo. Genet. Resour. Crop Evol. 2005, 52, 465–472. [Google Scholar] [CrossRef]
- Goulao, L.; Oliveira, C.M. Molecular characterization of cultivars of apple (Malus x domestica Borkh.) using microsatellite (SSR and ISSR) markers. Euphytica 2001, 122, 81–89. [Google Scholar] [CrossRef]
- Gorji, A.M.; Poczai, P.; Polgar, Z.; Taller, J. Efficiency of arbitrarily amplified dominant markers (SCoT, ISSR and RAPD) for diagnostic fingerprinting in tetraploid potato. Am. J. Potato Res. 2011, 88, 226–237. [Google Scholar] [CrossRef]
- Fernández, M.E.; Figueiras, A.M.; Benito, C. The use of ISSR and RAPD markers for detecting DNA polymorphism, genotype identification and genetic diversity among barley cultivars with known origin. Theor. Appl. Genet. 2002, 104, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Tanyolac, B. Inter-simple sequence repeat (ISSR) and RAPD variation among wild barley (Hordeum vulgare subsp. spontaneum) populations from west Turkey. Genet. Resour. Crop Evol. 2003, 50, 611–614. [Google Scholar] [CrossRef]
- Farajpour, M.; Ebrahimi, M.; Amiri, R.; Noori, S.A.S.; Sanjari, S.; Golzari, R. Study of genetic variation in yarrow using inter-simple sequence repeat (ISSR) and random amplified polymorphic DNA (RAPD) markers. Afr. J. Biotechnol. 2011, 10, 11137–11141. [Google Scholar]
- Patel, R.; Förster, D.; Kitchener, A.; Rayan, M.; Mahamed, S.; Werner, L.; Lenz, D.; Pfestorf, H.; Kramer-Schadt, S.; Radchuk, V.; et al. Two species of the genus Catopuma with diverging histories: An island endemic forest specialist and a wide-spread habitat generalist. R. Soc. Open Sci. 2016, 3, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.L.; Hu, Y.Q.; Sun, H. Allozyme analysis of Nouelia insignis and its meanings of biogeography and conservation biology. Acta Bot. Yunnanica 2003, 25, 563–571. [Google Scholar]
- Chapman, M.A.; Abbott, R.J. The origin of a novel form of Senecio (Asteraceae) restricted to sand dunes in southern Sicily. New Phytol. 2005, 166, 1051–1062. [Google Scholar] [CrossRef]
- Hatmaker, E.A.; Staton, M.E.; Dattilo, A.J.; Hadziabbic, D.; Rinehart, T.A.; Schilling, E.E.; Trigiano, R.N.; Wadl, P.A. Popularion structure and genetic diversity within the endangered species Pityopsis ruthii (Asteraceae). Front. Plant Sci. 2018, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Groves, H.; Burdon, J.J.; Kaye, P.E. Demography and genetics of Onopordum in southern New South Wales. J. Ecol. 1990, 78, 47–55. [Google Scholar] [CrossRef]
- O’Hanlon, P.C.; Peakall, R.; Briese, D.T. Amplified fragment length polymorphism (AFLP) reveals introgression in weedy Onopordum thistles: Hybridization and invasion. Mol. Ecol. 1999, 8, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Badr, A.; Morsy, W.; Abdelfattah, S.; Shams, S.; Shehab, A. Genetic diversity in Artemisia monosperma and Artemisia judaica populations in Egypt based on morphological, karyological and molecular variations. J. Med. Plants Res. 2011, 6, 66–78. [Google Scholar]
- Badr, A.; El-Shazly, H.; Helail, H.; Nagah, S.; El Ghanim, W. Genetic diversity of Artemisia populations in central and north Saudi Arabia based on morphological variation and RAPD polymorphism. Plant Syst. Evol. 2012, 298, 871–886. [Google Scholar] [CrossRef]
- Xiao, L.; Fayer, R.; Ryan, U.; Upton, S.J. Cryptosporidium taxonomy: Recent advances and implications for public health. Clin. Microbiol. Rev. 2004, 17, 72–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loveless, M.D.; Hamrick, J.L. Ecological determinants of genetic structure in plant populations. Annu. Rev. Ecol. Evol. Syst. 1984, 15, 65–95. [Google Scholar] [CrossRef]
- Millar, C.; Libby, W. Strategies for conserving clinal, ecotypic, and disjunct population diversity in widespread species. In Genetics and Conservation of Rare Plants; Falk, D.A., Holsingern, K.E., Eds.; Oxford University, Press: New York, NY, USA, 1991; pp. 149–170. [Google Scholar]
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S. Factor influencing levels of genetic diversity in woody plant species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- El-Midany, M.M.; Galal, T.M.; Farahat, E.A.; Hassan, L.M. Phytosociology of rainfed barley along the western Mediterranean Coast, Egypt. Taeckholmia 2019, 39, 18–33. [Google Scholar] [CrossRef]
- Slatkin, M. Gene flow and the geographic structure of natural populations. Science 1987, 236, 787–792. [Google Scholar] [CrossRef]
- Putman, A.I.; Carbone, I. Challenges in analysis and interpretation of microsatellite data for population genetic studies. Ecol. Evol. 2014, 4, 4399–4428. [Google Scholar] [CrossRef]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002; p. 607. [Google Scholar]
- Spielman, D.; Brook, B.W.; Frankham, R. Most species are not driven to extinction before genetic factors impact them. Proc. Natl. Acad. Sci. USA 2004, 101, 15261–15264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endels, P.; Jacquemyn, H.; Brys, R.; Hermy, M. Genetic erosion explains deviation from demographic response to disturbance and year variation in relic populations of the perennial Primula Vulgaris. J. Ecol. 2007, 95, 960–972. [Google Scholar] [CrossRef]
- Aguilar, R.; Ashworth, L.; Galetto, L.; Aizen, M.A. Plant re-productive susceptibility to habitat fragmentation: Review and synthesis through a meta-analysis. Ecol. Lett. 2006, 9, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Mashaly, I.A. Ecological Studies on Zygophyllum aegyptium in the Deltaic Mediterranean Coast of Egypt. Pak. J. Biol. Sci. 2002, 5, 152–160. [Google Scholar]
- Selim, A.A. Geology of El-Sallum Area, Western Mediterranean Coastal Zone, U.A.R. Ph.D. Thesis, Alexandria University, Alexandria, Egypt, 1969. [Google Scholar]
- UNESCO. Map of the World Distribution of Arid Regions; MAB Tech. Notes 7; United Nations Educational, Scientific and Cultural Organization; UNESCO: Paris, France, 1977; pp. 1–54. [Google Scholar]
- Rivas-Martnez, S.; Rivas-Sáenz, S.; Penas-Merino, A. Worldwide bioclimatic classification system. Glob. Geobot. 2011, 1, 1–638. [Google Scholar]
- Allen, S.E.; Grimshaw, H.M.; Rowland, A.P. Chemical analysis. In Methods of Plant Ecology; Moore, P.D., Chapman, S.B., Eds.; Blackwell Scientific Publications: Oxford, London, UK, 1986; pp. 285–344. [Google Scholar]
- Vile, D.; Garnier, E.; Shipley, B.; Laurent, G.; Navas, M.L.; Roumet, C.; Lavorel, S.; Díaz, S.; Hodgson, J.G.; Lloret, F.; et al. Specific leaf area and dry matter content estimate thickness in laminar leaves. Ann. Bot. 2005, 96, 11. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA mini preparation. Version III. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Williams, J.G.K.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.G.K.; Hamafay, M.K.; Rafalski, J.A.; Tingey, S.W. Genetic analysis using random amplified polymorphic DNA markers. Method Enzymol. 1993, 218, 704–740. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular cloning. A Laboratory Manual, 2nd ed.; Cold Spring Harbor, Laboratory Press: New York, NY, USA, 1989; pp. 3–9. [Google Scholar]
- Wilkinson, L. SYSTAT; Version 7, 13 new statistical routines and major revision of graphics; SPSS Inc.: Chicago, IL, USA, 1997. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Traits | Soil Variables | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Cl− | SO4−− | Ca++ | Mg++ | Na+ | K+ | OM | Total N | Total P | |
| Plant height | −0.25 | −0.01 | −0.09 | −0.02 | −0.12 | 0.14 | 0.01 | −0.27 | −0.30 |
| Leaf thickness | −0.21 | 0.53 | −0.40 | −0.10 | −0.12 | 0.25 | 0.20 | −0.25 | −0.38 |
| SLA | −0.37 | 0.47 | 0.55 | 0.61 | −0.36 | 0.14 | −0.01 | 0.65 | 0.49 |
| LDMC | 0.34 | −0.44 | 0.48 | 0.31 | 0.11 | −0.22 | −0.20 | 0.40 | 0.51 |
| Seed number | −0.58 | 0.17 | −0.63 | −0.69 | −0.85 | −0.19 | −0.58 | −0.64 | −0.42 |
| Seed mass | 0.06 | −0.13 | −0.08 | −0.34 | 0.15 | 0.12 | 0.17 | 0.05 | 0.09 |
| RAPD | ||||
| Genus | Onopordum Alexandrinum | |||
| Code | Oa1 | Oa2 | Oa3 | Oa4 |
| Bp | OP-A7 | |||
| 1100 | 0 | 0 | 0 | 1 |
| 1000 | 0 | 0 | 0 | 1 |
| 450 | 1 | 1 | 1 | 1 |
| 400 | 1 | 1 | 1 | 1 |
| 280 | 1 | 1 | 1 | 1 |
| 260 | 1 | 1 | 1 | 1 |
| 200 | 1 | 1 | 1 | 1 |
| Amplified bands | 5 | 5 | 5 | 7 |
| OP-A9 | ||||
| 630 | 0 | 0 | 0 | 1 |
| 480 | 1 | 1 | 1 | 1 |
| 300 | 1 | 1 | 1 | 1 |
| 230 | 1 | 1 | 1 | 1 |
| Amplified bands | 3 | 3 | 3 | 4 |
| OP-A18 | ||||
| 700 | 0 | 0 | 1 | 0 |
| 500 | 1 | 1 | 1 | 1 |
| 400 | 1 | 0 | 1 | 1 |
| 300 | 1 | 1 | 1 | 1 |
| Amplified bands | 3 | 2 | 4 | 3 |
| OP-B7 | ||||
| 1000 | 0 | 0 | 1 | 0 |
| 700 | 1 | 0 | 1 | 0 |
| 580 | 1 | 1 | 1 | 1 |
| 450 | 1 | 1 | 1 | 1 |
| 400 | 1 | 1 | 1 | 1 |
| 390 | 1 | 1 | 1 | 1 |
| 290 | 1 | 1 | 1 | 1 |
| Amplified bands | 6 | 5 | 7 | 5 |
| OP-C4 | ||||
| 580 | 1 | 0 | 1 | 0 |
| 500 | 1 | 1 | 1 | 1 |
| 400 | 1 | 1 | 1 | 1 |
| 360 | 1 | 1 | 1 | 1 |
| Amplified bands | 4 | 3 | 4 | 3 |
| OP-D9 | ||||
| 1000 | 0 | 0 | 1 | 0 |
| 930 | 1 | 0 | 1 | 0 |
| 800 | 1 | 1 | 1 | 1 |
| 700 | 1 | 1 | 1 | 1 |
| 570 | 1 | 1 | 1 | 1 |
| 400 | 1 | 1 | 1 | 1 |
| Amplified bands | 5 | 4 | 6 | 4 |
| ISSR | ||||
| Genus | Onopordum Alexandrinum | |||
| Code | Oa1 | Oa2 | Oa3 | Oa4 |
| Bp | 14 A | |||
| 660 | 1 | 1 | 1 | 1 |
| 585 | 1 | 1 | 1 | 1 |
| 530 | 1 | 1 | 1 | 1 |
| 440 | 1 | 1 | 1 | 1 |
| 400 | 0 | 1 | 0 | 0 |
| 390 | 1 | 0 | 0 | 0 |
| 375 | 0 | 0 | 0 | 0 |
| Amplified bands | 5 | 5 | 4 | 4 |
| 44A | ||||
| 1065 | 0 | 1 | 1 | 0 |
| 1040 | 0 | 0 | 0 | 1 |
| 865 | 0 | 0 | 1 | 0 |
| 835 | 0 | 0 | 0 | 1 |
| 690 | 1 | 1 | 1 | 1 |
| 600 | 1 | 0 | 1 | 1 |
| 535 | 0 | 0 | 1 | 1 |
| 530 | 1 | 0 | 0 | 0 |
| 475 | 1 | 0 | 1 | 1 |
| 460 | 0 | 1 | 0 | 0 |
| 390 | 0 | 0 | 0 | 1 |
| Amplified bands | 4 | 3 | 6 | 7 |
| 49A | ||||
| 885 | 1 | 1 | 1 | 1 |
| 615 | 0 | 0 | 0 | 1 |
| 540 | 0 | 0 | 0 | 1 |
| 450 | 1 | 0 | 1 | 1 |
| 400 | 1 | 1 | 1 | 1 |
| 285 | 1 | 1 | 1 | 1 |
| 255 | 1 | 0 | 1 | 1 |
| Amplified bands | 5 | 3 | 5 | 7 |
| 44B | ||||
| 970 | 0 | 0 | 0 | 1 |
| 620 | 1 | 1 | 1 | 1 |
| 560 | 1 | 1 | 1 | 1 |
| 475 | 1 | 1 | 1 | 0 |
| 440 | 1 | 1 | 1 | 1 |
| 385 | 1 | 1 | 1 | 1 |
| 325 | 1 | 1 | 1 | 1 |
| 295 | 1 | 1 | 1 | 1 |
| Amplified bands | 7 | 7 | 7 | 7 |
| HB-08 | ||||
| 545 | 0 | 1 | 1 | 0 |
| 500 | 0 | 0 | 0 | 1 |
| 470 | 1 | 1 | 1 | 0 |
| 440 | 0 | 0 | 0 | 1 |
| 390 | 1 | 1 | 1 | 0 |
| 350 | 1 | 1 | 1 | 1 |
| 315 | 1 | 1 | 1 | 1 |
| 285 | 1 | 1 | 1 | 0 |
| Amplified bands | 5 | 6 | 6 | 4 |
| HB-09 | ||||
| 730 | 1 | 1 | 1 | 1 |
| 570 | 1 | 1 | 1 | 1 |
| 480 | 1 | 1 | 1 | 1 |
| 430 | 1 | 1 | 1 | 1 |
| 335 | 1 | 1 | 1 | 1 |
| Amplified bands | 5 | 5 | 5 | 5 |
| Primers | Range of Band Products (bp) | Total Bands | Polymorphic Bands | |||||
|---|---|---|---|---|---|---|---|---|
| Total | Oa1 | Oa2 | Oa3 | Oa4 | ||||
| RAPD | OP-A7 | 200–1100 | 7 | 2 | 0 | 0 | 0 | 2 |
| OP-A9 | 230–630 | 4 | 1 | 0 | 0 | 0 | 1 | |
| OP-A18 | 300–700 | 4 | 2 | 1 | 0 | 2 | 1 | |
| OP-B7 | 290–1000 | 7 | 2 | 1 | 0 | 2 | 0 | |
| OP-C4 | 360–580 | 4 | 1 | 1 | 0 | 1 | 0 | |
| OP-D9 | 400–1000 | 6 | 2 | 1 | 0 | 2 | 0 | |
| Total bands | 32 | 10 | 4 | 0 | 7 | 4 | ||
| Percentage (%) | 31.25 | 12.5 | 0 | 21.88 | 12.5 | |||
| ISSR | 14 A | 390–660 | 6 | 2 | 1 | 1 | 0 | 0 |
| 44A | 390–1065 | 11 | 10 | 3 | 2 | 5 | 6 | |
| 49A | 255–885 | 7 | 4 | 2 | 0 | 2 | 4 | |
| 44B | 295–970 | 8 | 2 | 1 | 1 | 1 | 1 | |
| HB08 | 285–545 | 8 | 6 | 3 | 4 | 4 | 2 | |
| HB09 | 335–730 | 5 | 0 | 0 | 0 | 0 | 0 | |
| Total bands | 45 | 24 | 10 | 8 | 12 | 13 | ||
| Percentage (%) | 53.33 | 22.22 | 17.78 | 26.67 | 28.89 | |||
| Accessions | Oa1 | Oa2 | Oa3 | Oa4 |
|---|---|---|---|---|
| Oa1 | 1.000 | |||
| Oa2 | 0.513 | 1.000 | ||
| Oa3 | 0.630 | 0.509 | 1.000 | |
| Oa4 | 0.300 | 0.200 | 0.213 | 1.000 |
| RAPD | ISSR | ||
|---|---|---|---|
| Codes | Nucleotide Sequence 5’to 3’ | Codes | Nucleotide Sequence 5’to 3’ |
| OP-A7 | 5’ GAA AGG GGT G 3’ | 14A | CTC TCT CTC TCT CTC TTG |
| OP-A9 | 5´ GGG TAA CGC C 3’ | 44A | CAC ACA CAC ACA AG |
| OP-A18 | 5´AGG TGA CCG T 3’ | 49A | GTAGATTATGTTCCTTCTCC |
| OP-B7 | 5´ GAA AGG GGT G 3’ | 44B | CTC TCT CTC TCT CTC TGC |
| OP-C4 | 5´ CCG CAT CTA C 3’ | HB8 | GAG AGA GAG AGA GG |
| OP-D9 | 5´ GGT GAC GCA G 3’ | HB9 | GTG TGT GTG TGT GG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yahia, A.; Mashaly, I.; El-Bana, M.; Rizk, R.; El-Sherbeny, G. Intraspecific Variations in Functional and Molecular Traits of Near-Endemic Onopordum alexandrinum Boiss. in Natural and Anthropogenic Habitats along the Western Mediterranean Coast of Egypt: Implications for Conservation. Plants 2020, 9, 1041. https://doi.org/10.3390/plants9081041
Yahia A, Mashaly I, El-Bana M, Rizk R, El-Sherbeny G. Intraspecific Variations in Functional and Molecular Traits of Near-Endemic Onopordum alexandrinum Boiss. in Natural and Anthropogenic Habitats along the Western Mediterranean Coast of Egypt: Implications for Conservation. Plants. 2020; 9(8):1041. https://doi.org/10.3390/plants9081041
Chicago/Turabian StyleYahia, Aya, Ibrahim Mashaly, Magdy El-Bana, Rehab Rizk, and Ghada El-Sherbeny. 2020. "Intraspecific Variations in Functional and Molecular Traits of Near-Endemic Onopordum alexandrinum Boiss. in Natural and Anthropogenic Habitats along the Western Mediterranean Coast of Egypt: Implications for Conservation" Plants 9, no. 8: 1041. https://doi.org/10.3390/plants9081041