Water Shortage Strongly Alters Formation of Calcium Oxalate Druse Crystals and Leaf Traits in Fagopyrum esculentum

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Design
2.2. Biochemical Analysis
2.3. Physiological Analysis
2.4. Optical Measurements
2.5. Morphological Measurements
2.6. Leaf Bulk Element Analysis
2.7. Statistical Analyses
3. Results
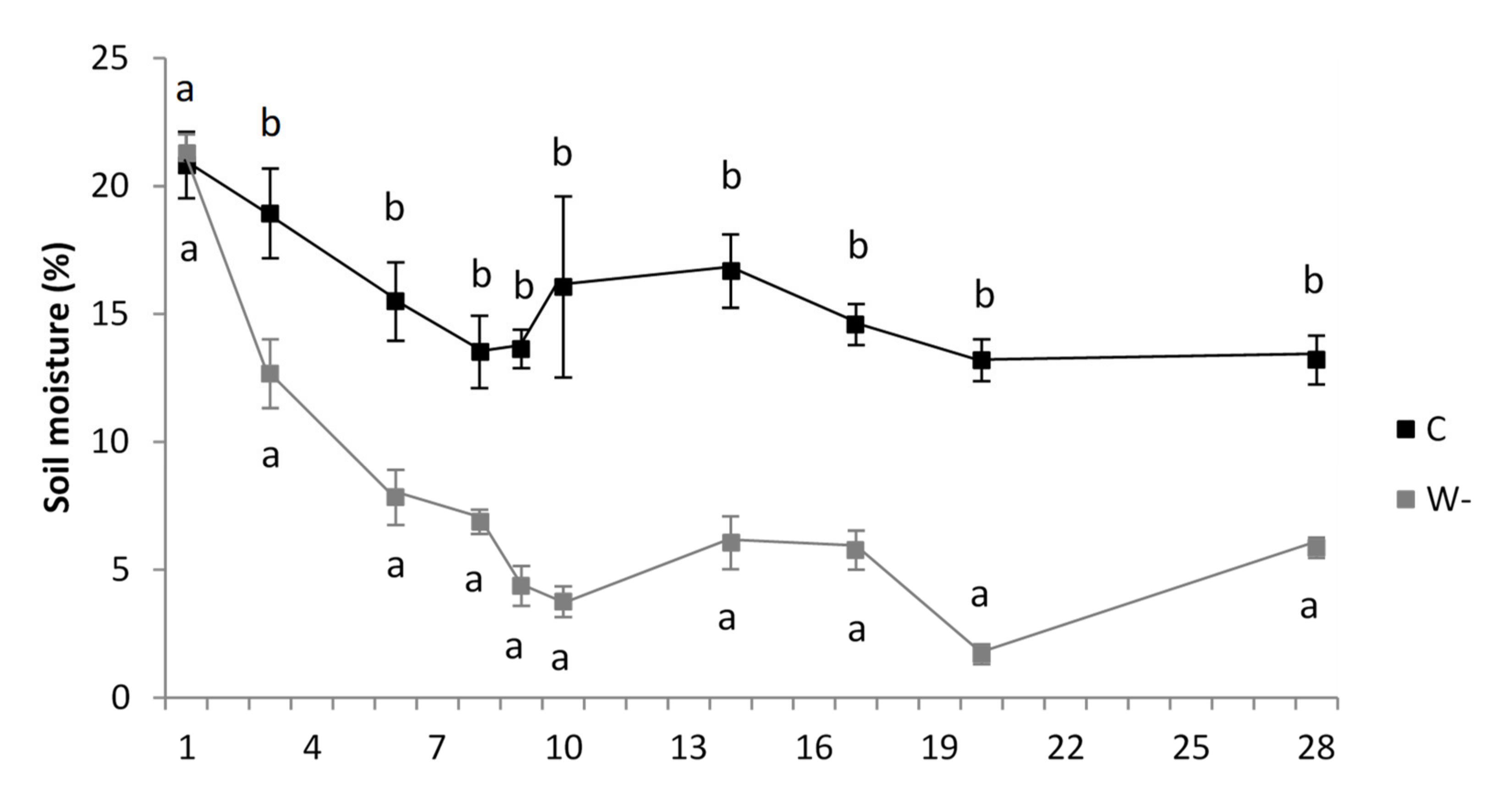
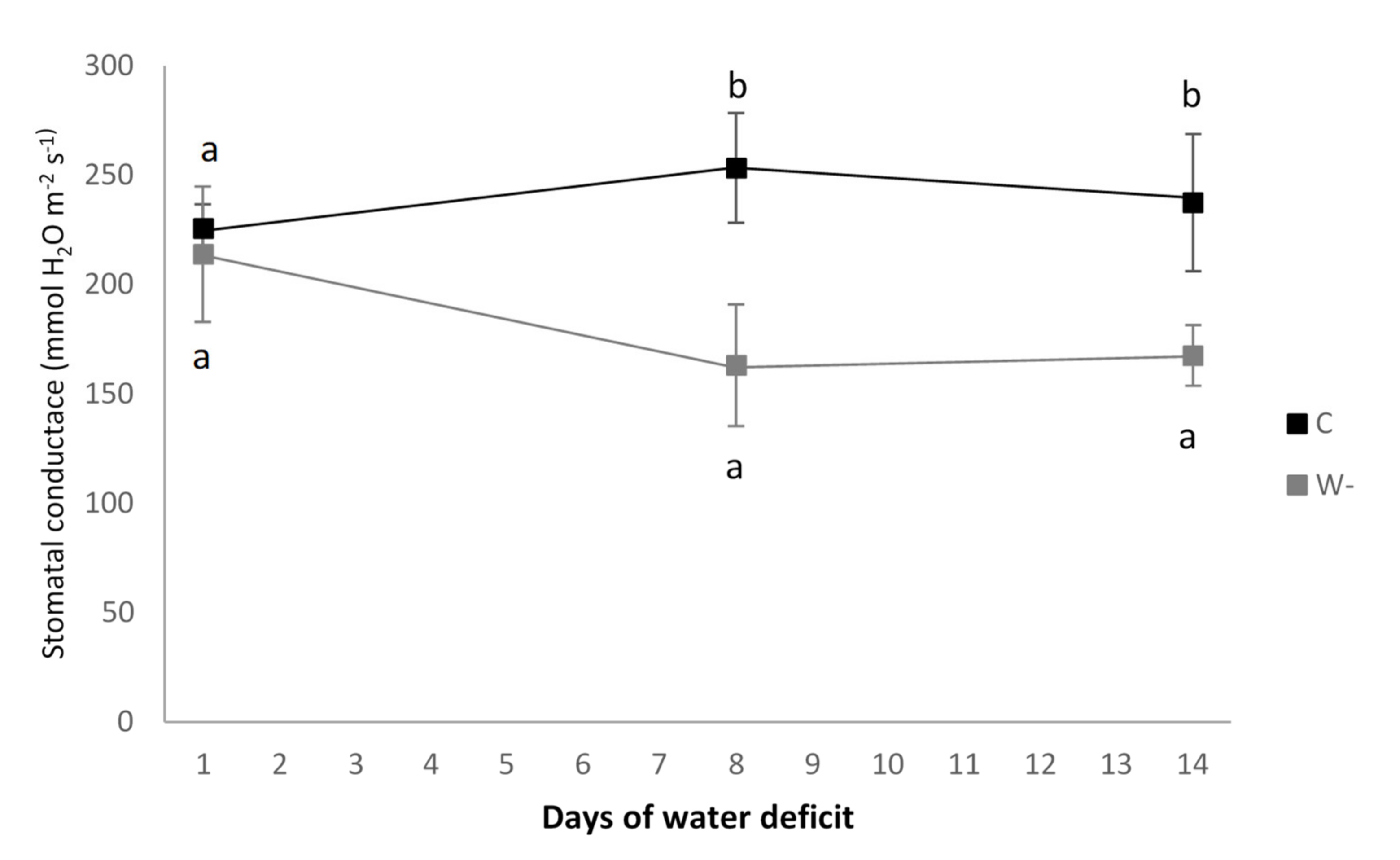
3.1. Growth Conditions and Leaf Traits
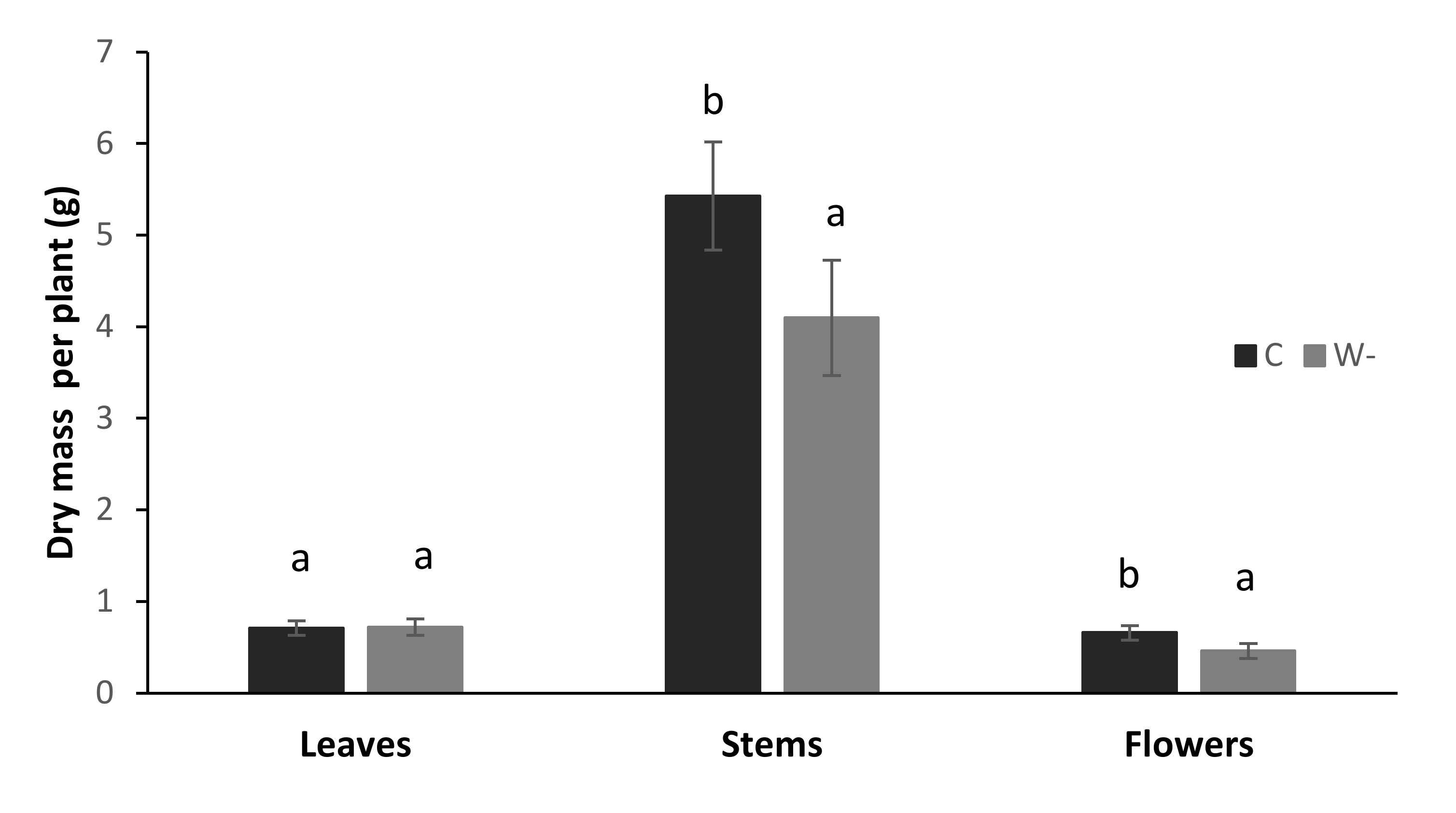
3.2. Plant Biomass
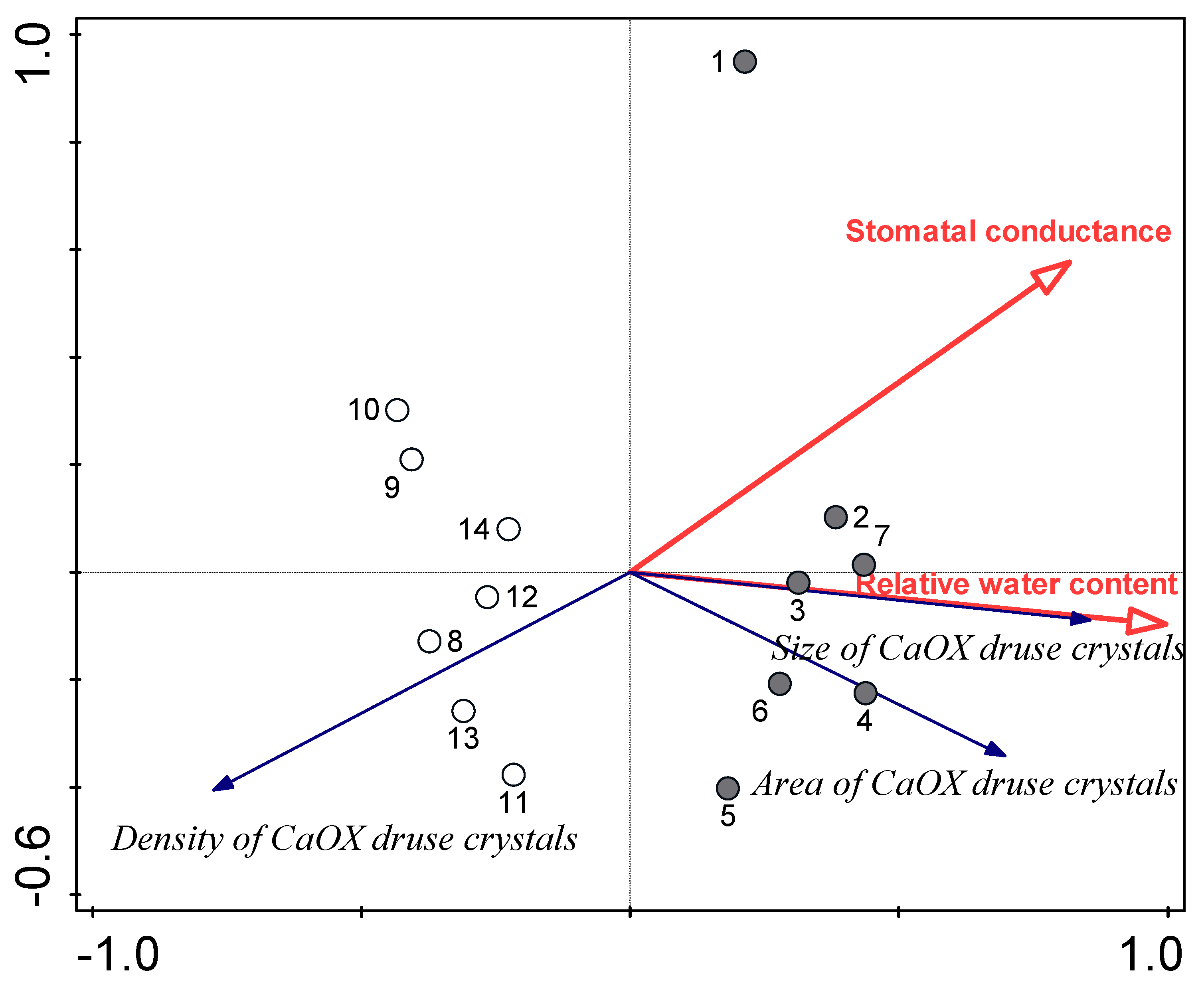
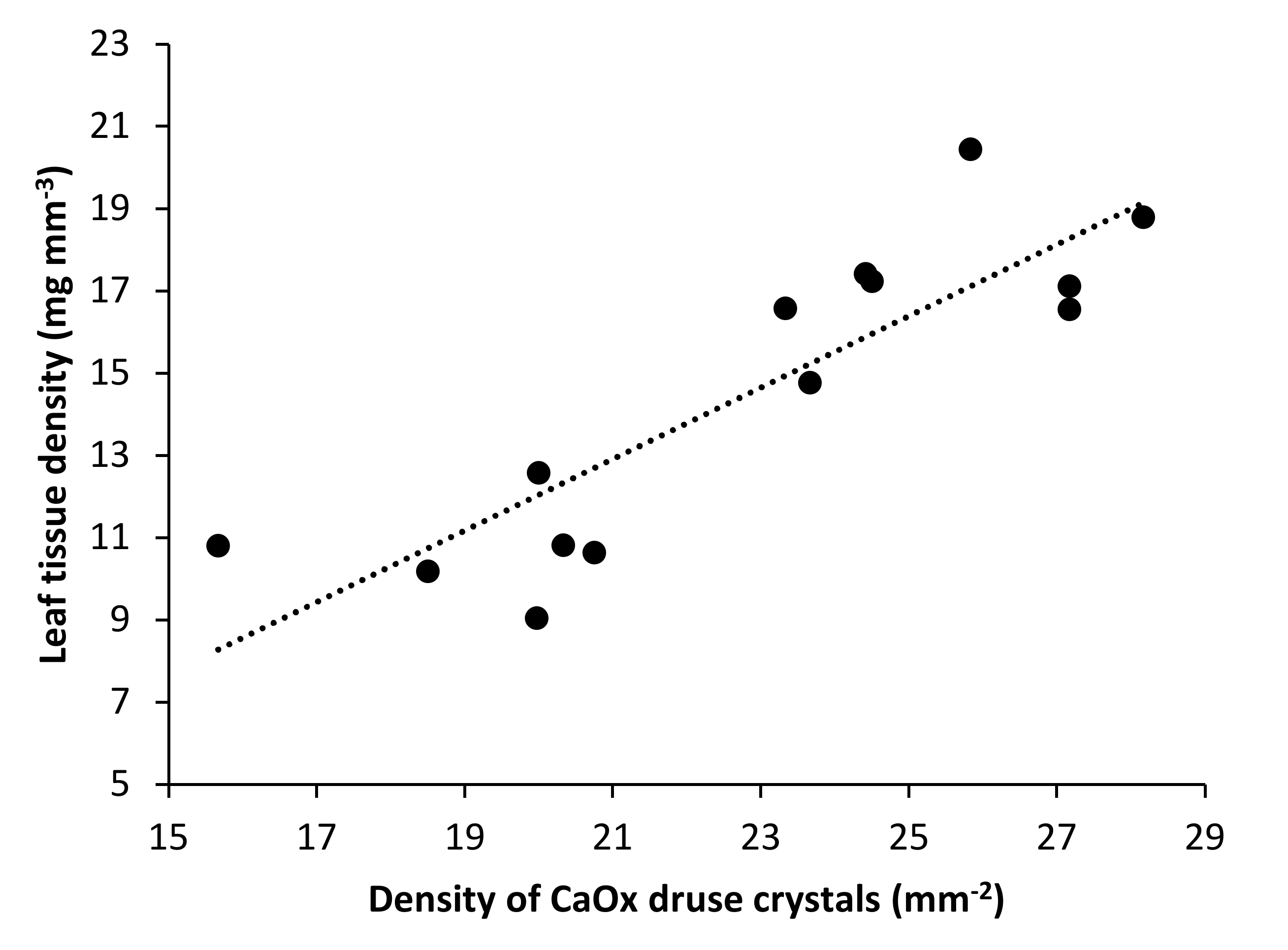
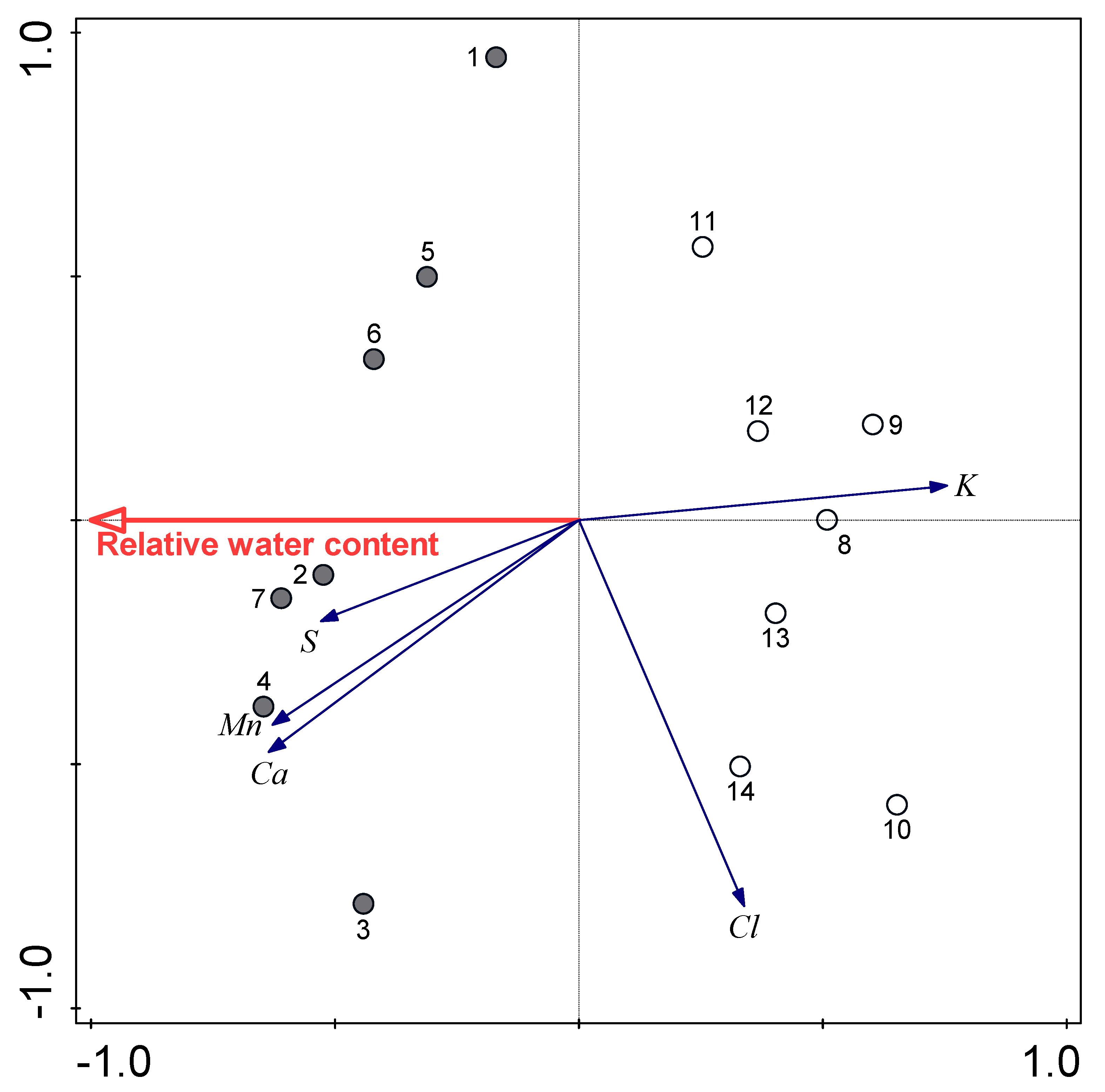
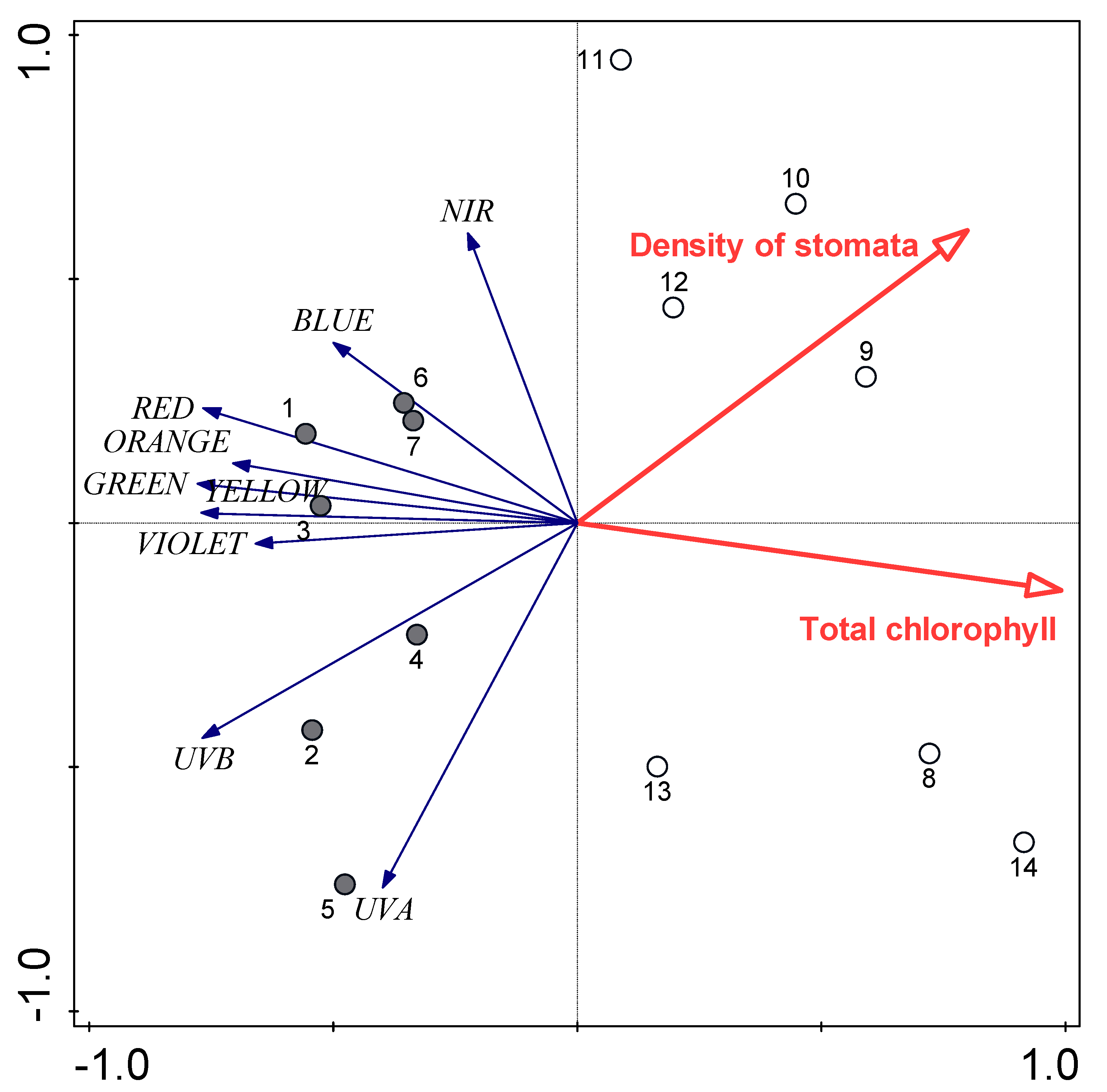
3.3. Relationships between Water Availability Indicators and Certain Leaf Traits
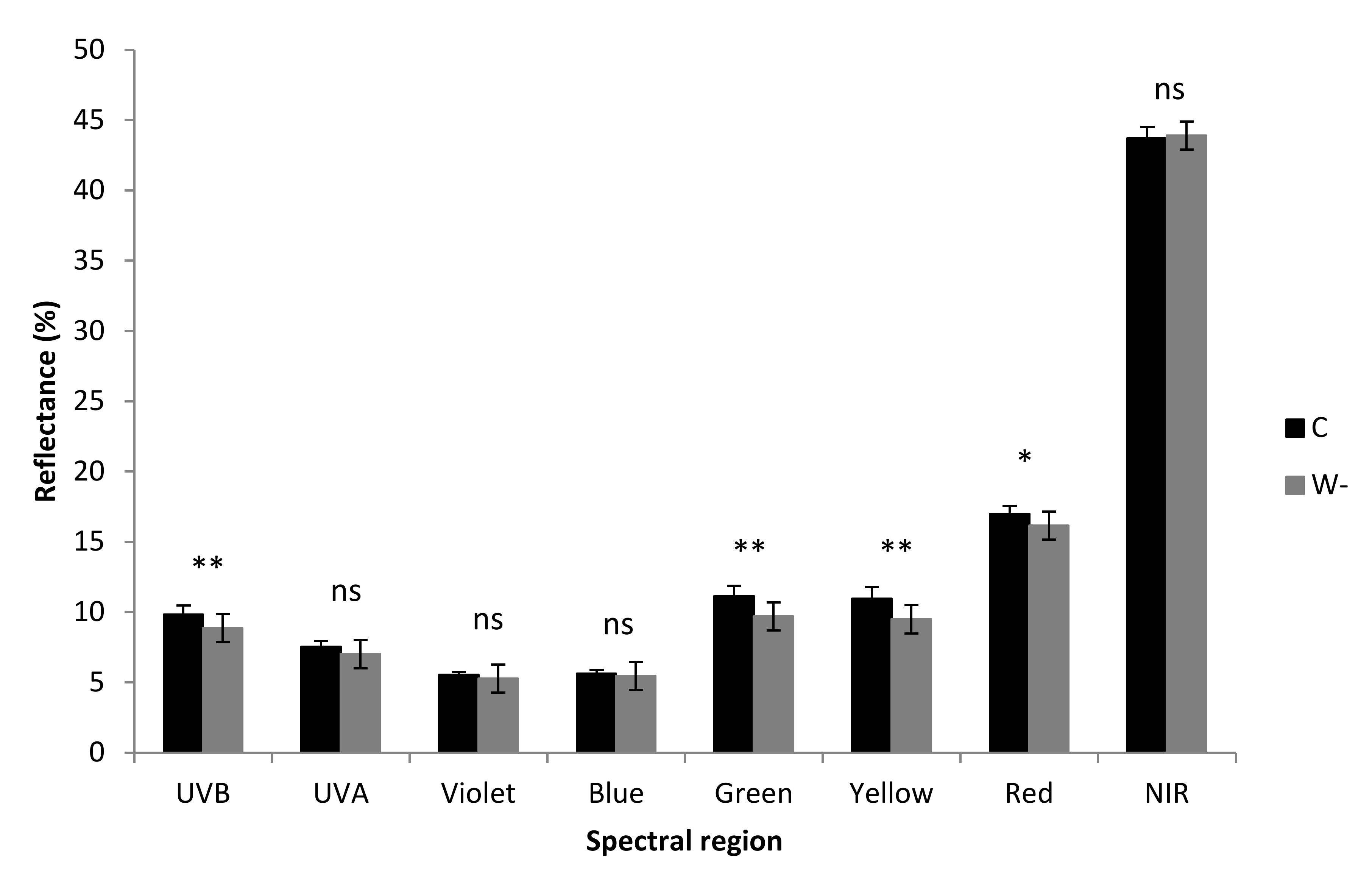
3.4. Optical Properties
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crop adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Ye, W.; Wang, M.; Yan, X. Climate change and drought: A risk assessment of crop-yield impacts. Clim. Res. 2009, 39, 31–46. [Google Scholar] [CrossRef]
- Venuprasad, R.; Lafitte, H.; Atlin, G. Response to direct selection for grain yield under drought stress in rice. Crop. Sci. 2007, 47, 285–293. [Google Scholar] [CrossRef]
- Aghaie, P.; Tafreshi, S.A.H.; Ebrahimi, M.A.; Haerinasaba, M. Tolerance evaluation and clustering of fourteen tomato cultivars grown under mild and severe drought conditions. Sci. Hortic. 2018, 232, 1–12. [Google Scholar] [CrossRef]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups; Springer: Berlin, Germany, 2003. [Google Scholar]
- Kasajima, S.; Hatae, K.; Morishita, T. Seed-setting habit of a semi-dwarf common buckwheat line. Fagopyrum 2017, 34, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; Didio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.A.; Zhukov, V.; Tikhonovich, I.; Tarakhovskaya, E.; et al. Methodology of drought stress research: Experimental set-up and physiological characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Yin, L.; Shan, L.; Deng, X. Genotypic variation in growth and physiological response to drought stress and re-watering reveals the critical role of recovery in drought adaptation in maize seedlings. Front. Plant Sci. 2016, 6, 1241. [Google Scholar] [CrossRef] [Green Version]
- Gaberščik, A.; Vončina, M.; Trošt Sedej, T.; Germ, M.; Björn, L.O. Growth and production of buckwheat (Fagopyrum esculentum) treated with reduced, ambient, and enhanced UV-B radiation. J. Photochem. Photobiol. B Biol. 2002, 66, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Breznik, B.; Germ, M.; Gaberščik, A.; Kreft, I. Combined effects of elevated UV-B radiation and the addition of selenium on common (Fagopyrum esculentum Moench) and tartary (Fagopyrum tataricum (L.) Gaertn.) buckwheat. Photosyntetica 2005, 43, 583–589. [Google Scholar] [CrossRef]
- Dolinar, N.; Germ, M.; Kreft, I.; Breznik, B.; Gaberščik, A. Effects of water deficit and selenium on common buckwheat (Fagopyrum esculentum Moench.) plants. Photosyntetica 2007, 45, 472–476. [Google Scholar]
- Germ, M.; Breznik, B.; Dolinar, N.; Kreft, I.; Gaberščik, A. The combined effect of water limitation and UV-B radiation on common and tartary buckwheat. Cereal Res. Commun. 2013, 41, 97–105. [Google Scholar] [CrossRef]
- Golob, A.; Kugovnik, A.; Kreft, I.; Gaberščik, A.; Germ, M. The interactions between UV radiation, drought and selenium in different buckwheat species. Acta Biol. Slov. 2019, 62, 57–66. [Google Scholar]
- Musilová, J.; Lachman, J.; Bystrická, J.; Vollmannová, A.; Čičová, I.; Timoracká, M. Cultivar and growth phases–the factors affecting antioxidant activity of buckwheat (Fagopyrum esculentum Moench.). Acta Agric. Slov. 2013, 101, 201–208. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Golob, A.; Stibilj, V.; Nečemer, M.; Kump, P.; Kreft, I.; Hočevar, A.; Gaberščik, A.; Germ, M. Calcium oxalate druses affect leaf optical properties in selenium-treated Fagopyrum tataricum. J. Photochem. Photobiol. B Biol. 2018, 180, 51–55. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Horner, H.T. Calcium oxalate crystals in plants. Bot. Rev. 1980, 46, 361–427. [Google Scholar] [CrossRef]
- Webb, M.A. Cell-mediated crystallization of calcium oxalate in plants. Plant Cell. 1999, 11, 751–761. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, L.P.; Mansfield, T.A. A postulated role for calcium oxalate in the regulation of calcium ions in the vicinity of stomatal guard cells. New Phytol. 1994, 127, 473–481. [Google Scholar] [CrossRef]
- Kreft, I.; Zhou, M.; Golob, A.; Germ, M.; Likar, M.; Dziedzic, K.; Luthar, Z. Breeding buckwheat for nutritional quality. Breed. Sci. 2019, 70, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Grašič, M.; Malovrh, U.; Golob, A.; Vogel-Mikuš, K.; Gaberščik, A. Effects of water availability and UV radiation on silicon accumulation in the C4 crop proso millet. Photochem. Photobiol. Sci. 2019, 18, 375–386. [Google Scholar] [CrossRef]
- Grašič, M.; Dobravc, M.; Golob, A.; Vogel-Mikuš, K.; Gaberščik, A. Water shortage reduces silicon uptake in barley leaves. Agric. Water Manag. 2019, 217, 47–56. [Google Scholar] [CrossRef]
- Grašič, M.; Golob, A.; Vogel-Mikuš, K.; Gaberščik, A. Severe water deficiency during the mid-vegetative and reproductive phase has little effect on proso millet performance. Water 2019, 11, 2155. [Google Scholar] [CrossRef] [Green Version]
- Martinčič, A.; Wraber, T.; Jogan, N.; Podobnik, A.; Turk, B.; Vreš, B.; Ravnik, V.; Frajman, B.; Strgulc Krajšek, S.; Trčak, B.; et al. Mala flora Slovenije–Ključ za Določanje Praprotnic in Semenk; 4. Izdaja; Tehniška založba Slovenije: Ljubljana, Slovenia, 2007; 967p. [Google Scholar]
- Halbrecq, B.; Romedenne, P.; Ledent, J.F. Evolution of flowering, ripening and seed set in buckwheat (Fagopyrum esculentum Moench): Quantitative analysis. Eur. J. Agron. 2005, 23, 209–224. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Extraction of photosynthetic tissues: Chlorophylls and carotenoids. Curr. Protoc. Food Anal. Chem. 2001, 1, 165–170. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, 171–178. [Google Scholar] [CrossRef]
- Drumm, H.; Mohr, H. The mode of interaction between blue (UV) light photoreceptor and phytochrome in anthocyanin formation of the Sorghum seedling. Photochem. Photobiol. 1978, 27, 241–248. [Google Scholar] [CrossRef]
- Caldwell, M.M. Solar ultraviolet radiation as an ecological factor for alpine plants. Ecol. Monogr. 1968, 38, 243–268. [Google Scholar] [CrossRef]
- Schreiber, U.; Kühl, M.; Klimant, I.; Reising, H. Measurement of chlorophyll fluorescence within leaves using a modified PAM fluorometer with a fiber-optic microprobe. Photosynth. Res. 1996, 47, 103–109. [Google Scholar] [CrossRef]
- Packard, T.T. The measurement of respiratory electron transport activity in marine phytoplankton. J. Mar. Res. 1971, 29, 215–244. [Google Scholar]
- Kenner, A.A.; Ahmed, S.I. Measurements of electron transport activities in marine phytoplankton. Mar. Biol. 1975, 3, 19–127. [Google Scholar] [CrossRef]
- Klančnik, K.; Vogel-Mikuš, K.; Gaberščik, A. Silicified structures affect leaf optical properties in grasses and sedge. J. Photochem. Photobiol. B 2014, 130, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bondada, B.R. An array of simple, fast, and safe approaches to visualizing fine cellular structures in free-hand sections of stem, leaf, and fruit using optical microscopy. Curr. Bot. 2012, 3, 11–22. [Google Scholar]
- Nečemer, M.; Kump, P.; Ščančar, J.; Jaćimović, R.; Simčič, J.; Pelicon, P.; Budnar, M.; Jeran, Z.; Pongrac, P.; Regvar, M.; et al. Application of X-ray fluorescence analytical techniques in phytoremediation and plant biology studies. Spectrochim. Acta Part B At. Spectrosc. 2008, 63, 1240–1247. [Google Scholar] [CrossRef]
- Vekemans, B.; Janssens, K.; Vincze, L.; Adams, F.; Vanespen, P. Analysis of X-ray spectra by iterative least squares (AXIL): New developments. X-ray Spectrom. 1994, 23, 278–285. [Google Scholar] [CrossRef]
- Kump, P.; Nečemer, M.; Rupnik, Z.; Pelicon, P.; Ponikvar, D.; Vogel-Mikuš, K.; Regvar, M.; Pongrac, P. Improvement of the XRF quantification and enhancement of the combined applications by EDXRF and micro-PIXE. In Integration of Nuclear Spectrometry Methods as a New Approach to Material Research; IAEA: Vienna, Austria, 2011; pp. 101–109. [Google Scholar]
- ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows Userʹs Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Björkman, O.; Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef]
- Pfündel, E. Estimating the contribution of photosystem I to total leaf chlorophyll fluorescence. Photosyn. Res. 1998, 56, 185–195. [Google Scholar] [CrossRef]
- Urban, L.; Aarrouf, J.; Bidel, L.P.R. Assessing the effects of water deficit on photosynthesis using parameters derived from measurements of leaf gas exchange and of chlorophyll a fluorescence. Front. Plant Sci. 2017, 8, 2068. [Google Scholar] [CrossRef] [Green Version]
- Tourneux, C.; Peltier, G. Effect of water deficit on photosynthetic oxygen exchange measured using 18O2 and mass spectrometry in Solanum tuberosum L. leaf discs. Planta 1995, 195, 570–577. [Google Scholar] [CrossRef]
- Germ, M.; Kreft, I.; Stibilj, V.; Urbanc-Berčič, O. Combined effects of selenium and drought on photosynthesis and mitochondrial respiration in potato. Plant Physiol. Biochem. 2007, 45, 162–167. [Google Scholar] [CrossRef]
- Miyashita, K.; Tanakamaru, S.; Maitani, T.; Kimura, K. Recovery responses of photosynthesis, transpiration, and stomatal conductance in kidney bean following drought stress. Environ. Exp. Bot. 2005, 53, 205–214. [Google Scholar] [CrossRef]
- Pankovic, D.; Sakac, Z.; Kevres, S.; Plesnicar, M. Acclimation to long-term water deficit in the leaves of two sunflower hybrids: Photosynthesis, electron transport and carbon metabolism. J. Exp. Bot. 1999, 330, 127–138. [Google Scholar] [CrossRef]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.P.; Rodrigues, M.L.; Picardo, C.P.P.; Osorio, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How plants cope with water stress in the field. Photosynthesis and growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, P.; Wahbi, S.; Tsonev, T.; Haworth, M.; Liu, S.; Centritto, M. On the use of leaf spectral indices to assess water status and photosynthetic limitations in Olea europaea L. during water stress and recovery. PLoS ONE 2014, 9, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, W.M. Effects of water deficit on photosynthetic capacity. Physiol. Plant. 1987, 71, 1. [Google Scholar] [CrossRef]
- de la Riva, E.G.; Olmo, M.; Poorter, H.; Ubera, J.L.; Villar, R. Leaf mass per area (LMA) and its relationship with leaf structure and anatomy in 34 Mediterranean woody species along a water availability gradient. PLoS ONE 2016, 11, e0148788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farquharson, K.L. Fine-tuning plant growth in the face of drought. Plant Cell 2017, 29, 4. [Google Scholar] [CrossRef] [Green Version]
- Koch, G.; Rolland, G.; Dauzat, M.; Bédiée, A.; Baldazzi, V.; Bertin, N.; Guédon, Y.; Granier, C. Leaf production and expansion: A generalized response to drought stresses from cells to whole leaf biomass—A case study in the tomato compound leaf. Plants 2019, 8, 409. [Google Scholar] [CrossRef] [Green Version]
- Lehmeier, C.; Pajor, R.; Lundgren, M.R.; Mathers, A.; Sloan, J.; Bauch, M.; Mitchell, A.; Bellasio, C.; Green, A.; Bouyer, D.; et al. Cell density and airspace patterning in the leaf can be manipulated to increase leaf photosynthetic capacity. Plant. J. 2017, 92, 981–994. [Google Scholar] [CrossRef]
- Lee, L.R.; Bergmann, D.C. The plant stomatal lineage at a glance. J. Cell Sci. 2019, 132, jcs228551. [Google Scholar] [CrossRef] [Green Version]
- Rozema, J.; Björn, L.O.; Bornman, J.F.; Gaberščik, A.; Häder, D.P.; Trošt, T.; Germ, M.; Klisch, M.; Gröniger, A.; Sinha, R.P.; et al. The role of UV-B radiation in aquatic and terrestrial ecosystems-an experimental and functional analysis of the evolution of UV-absorbing compounds. J. Photochem. Photobiol. B Biol. 2002, 66, 2–12. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Front. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [Green Version]
- Mathesius, U. Flavonoid functions in plants and their interactions with other organisms. Plants 2018, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Basahi, M.J.; Ismail, I.M.; Hassan, I.A. Effects of enhanced UV-B radiation and drought stress on photosynthetic performance of lettuce (Lactuca sativa L. Romaine) plants. Annu. Res. Rev. Biol. 2014, 4, 1739–1756. [Google Scholar] [CrossRef]
- Nakabayashi, R.; Mori, T.; Saito, K. Alternation of flavonoid accumulation under drought stress in Arabidopsis thaliana. Plant Signal. Behav. 2014, 9, e29518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Farooq, S.; Hasan, W.; Ul-Allah, S.; Tanveer, M.; Farooqh, M.; Nawaz, A. Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agric. Water Manag. 2018, 201, 152–166. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Elsevier: London, UK, 1995; p. 889. [Google Scholar]
- Franceschi, V.R.; Nakata, P.A. Calcium oxalate in plants: Formation and function. Annu. Rev. Plant Biol. 2005, 56, 41–71. [Google Scholar] [CrossRef] [PubMed]
- Ustin, S.L.; Jacquemoud, S.; Govaerts, Y. Simulation of photon transport in a three-dimensional leaf: Implications for photosynthesis. Plant Cell Environ. 2001, 24, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Klančnik, K.; Mlinar, M.; Gaberščik, A. Heterophylly results in a variety of “spectral signatures” in aquatic plant species. Aquat. Bot. 2012, 98, 20–26. [Google Scholar] [CrossRef]
- Brotherton, S.J.; Joyce, C.B. Extreme climate events and wet grasslands: Plant traits for ecological resilience. Hydrobiologia 2015, 750, 229–243. [Google Scholar] [CrossRef] [Green Version]
- Gutschick, V.P.; BassiriRad, H. Extreme events as shaping physiology, ecology, and evolution of plants: Toward a unified definition and evaluation of their consequences. New Phytol. 2003, 160, 21. [Google Scholar] [CrossRef]
- Volk, G.M.; Lynch-Holm, V.J.; Kostman, T.A.; Goss, L.J.; Franceschi, V.R. The role of druse and raphide calcium oxalate crystals in tissue calcium regulation in Pistia stratiotes leaves. Plant Biol. 2002, 4, 34–45. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.-S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Trait | Treatment | p | |
|---|---|---|---|
| Control | Water Shortage | ||
| Anatomical and morphological | |||
| Relative water content (%) | 79.5 ± 2.33 b | 67.1 ± 2.04 a | <0.001 |
| Specific leaf area (cm2 mg−1 DM) | 0.34 ± 0.04 b | 0.30 ± 0.2 a | 0.049 |
| Tissue density (mg mm−3) | 11.3 ± 1.9 a | 17.7 ± 1.4 b | <0.001 |
| Leaf thickness (µm) | 275.4 ± 31.0 b | 191.8 ± 6.3 a | <0.001 |
| Palisade mesophyll (µm) | 136.1 ± 22.0 b | 88.4 ± 5.5 a | <0.001 |
| Spongy mesophyll (µm) | 85.9 ± 10.7 b | 59.5 ± 3.5 a | <0.001 |
| Density of CaOx druse crystals (mm−2) | 19.8 ± 2.4 a | 25.8 ± 1.9 b | <0.001 |
| Diameter of CaOx druse crystals (µm) | 33.3 ± 4.3 b | 25.2 ± 1.4 a | <0.001 |
| Area of CaOx druse crystals (% transection area) | 1.78 ± 0.46 b | 1.29 ± 0.17 a | 0.021 |
| Adaxial leaf surface | |||
| Epidermis thickness (µm) | 29.6 ± 3.0 a | 27.5 ± 5.2 a | 0.115 |
| Stomata density (mm−2) | 20.9 ± 3.4 a | 31.7 ± 4.0 b | <0.001 |
| Stomata length (µm) | 35.8 ± 2.0 b | 30.4 ± 1.0 a | <0.001 |
| Stomata width (µm) | 23.0 ± 0.8 b | 21.7 ± 1.2 a | 0.033 |
| Abaxial surface | |||
| Epidermis thickness (µm) | 22.9 ± 2.2 a | 21.7 ± 1.2 a | 0.209 |
| Stomata density (mm−2) | 50.0 ± 13.6 a | 94.7 ± 9.7 b | <0.001 |
| Stomata length (µm) | 34.2 ± 2.2 b | 28.3 ± 1.2 a | <0.001 |
| Stomata width (µm) | 21.9 ± 0.7 b | 20.1 ± 1.1 a | 0.003 |
| Biochemical | |||
| Total chlorophyll (mg dm−2) | 3.16 ± 0.26 a | 5.37 ± 0.95 b | <0.001 |
| Carotenoids (mg dm−2) | 0.49 ± 0.10 a | 0.74 ± 0.09 b | <0.001 |
| Anthocyanins (rel. units cm−2) | 0.76 ± 0.06 a | 0.70 ± 0.13 a | 0.357 |
| UV-B–absorbing compounds (rel. units cm−2) | 160 ± 23 a | 232 ± 27 b | <0.001 |
| UV-A–absorbing compounds (rel. units cm−2) | 314 ± 60 a | 495 ± 60 b | <0.001 |
| Physiological | |||
| Stomatal conductance (mmol H2O m−2 s−1) | 253 ± 25 b | 163 ± 28 a | <0.001 |
| Fv/Fm | 0.8 ± 0.02 a | 0.79 ± 0.01 a | 0.780 |
| ∆Fv/Fm’ | 0.68 ± 0.02 a | 0.67 ± 0.02 a | 0.414 |
| ETS (µL O2 mg−1 DM h−1) | 2.55 ± 0.44 a | 2.65 ± 0.25 a | 0.605 |
| Element | Element Contents (mg g−1 DM) according to Treatment | p | |
|---|---|---|---|
| Control | Water Shortage | ||
| Phosphorus | 2.34 ± 0.38 a | 2.50 ± 0.49 a | 0.513 |
| Sulphur | 0.51 ± 0.15 b | 0.38 ± 0.07 a | 0.049 |
| Chlorine | 9.42 ± 4.15 a | 12.94 ± 2.71 a | 0.084 |
| Potassium | 12.86 ± 2.13 a | 21.80 ± 4.79 b | 0.001 |
| Calcium | 38.98 ± 3.56 b | 32.22 ± 4.71 a | 0.011 |
| Manganese | 0.13 ± 0.03 b | 0.10 ± 0.02 a | 0.048 |
| Iron | 0.27 ± 0.02 a | 0.28 ± 0.03 a | 0.707 |
| Copper | 0.016 ± 0.004 a | 0.017 ± 0.004 a | 0.620 |
| Zinc | 0.067 ± 0.01 a | 0.078 ± 0.016 a | 0.181 |
| Spectral Region | Leaf Traits | Explained Variance (%) | p |
|---|---|---|---|
| UV | Size of CaOx druse crystals | 46.2 | 0.004 |
| Violet, blue | Total chlorophyll | 35.9 | 0.017 |
| Calcium content | 18.4 | 0.034 | |
| Green, yellow | Total chlorophyll | 58.6 | 0.002 |
| Red, near-infrared | Total chlorophyll | 35.9 | 0.002 |
| Calcium content | 24.6 | 0.005 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaberščik, A.; Grašič, M.; Vogel-Mikuš, K.; Germ, M.; Golob, A. Water Shortage Strongly Alters Formation of Calcium Oxalate Druse Crystals and Leaf Traits in Fagopyrum esculentum. Plants 2020, 9, 917. https://doi.org/10.3390/plants9070917
Gaberščik A, Grašič M, Vogel-Mikuš K, Germ M, Golob A. Water Shortage Strongly Alters Formation of Calcium Oxalate Druse Crystals and Leaf Traits in Fagopyrum esculentum. Plants. 2020; 9(7):917. https://doi.org/10.3390/plants9070917
Chicago/Turabian StyleGaberščik, Alenka, Mateja Grašič, Katarina Vogel-Mikuš, Mateja Germ, and Aleksandra Golob. 2020. "Water Shortage Strongly Alters Formation of Calcium Oxalate Druse Crystals and Leaf Traits in Fagopyrum esculentum" Plants 9, no. 7: 917. https://doi.org/10.3390/plants9070917