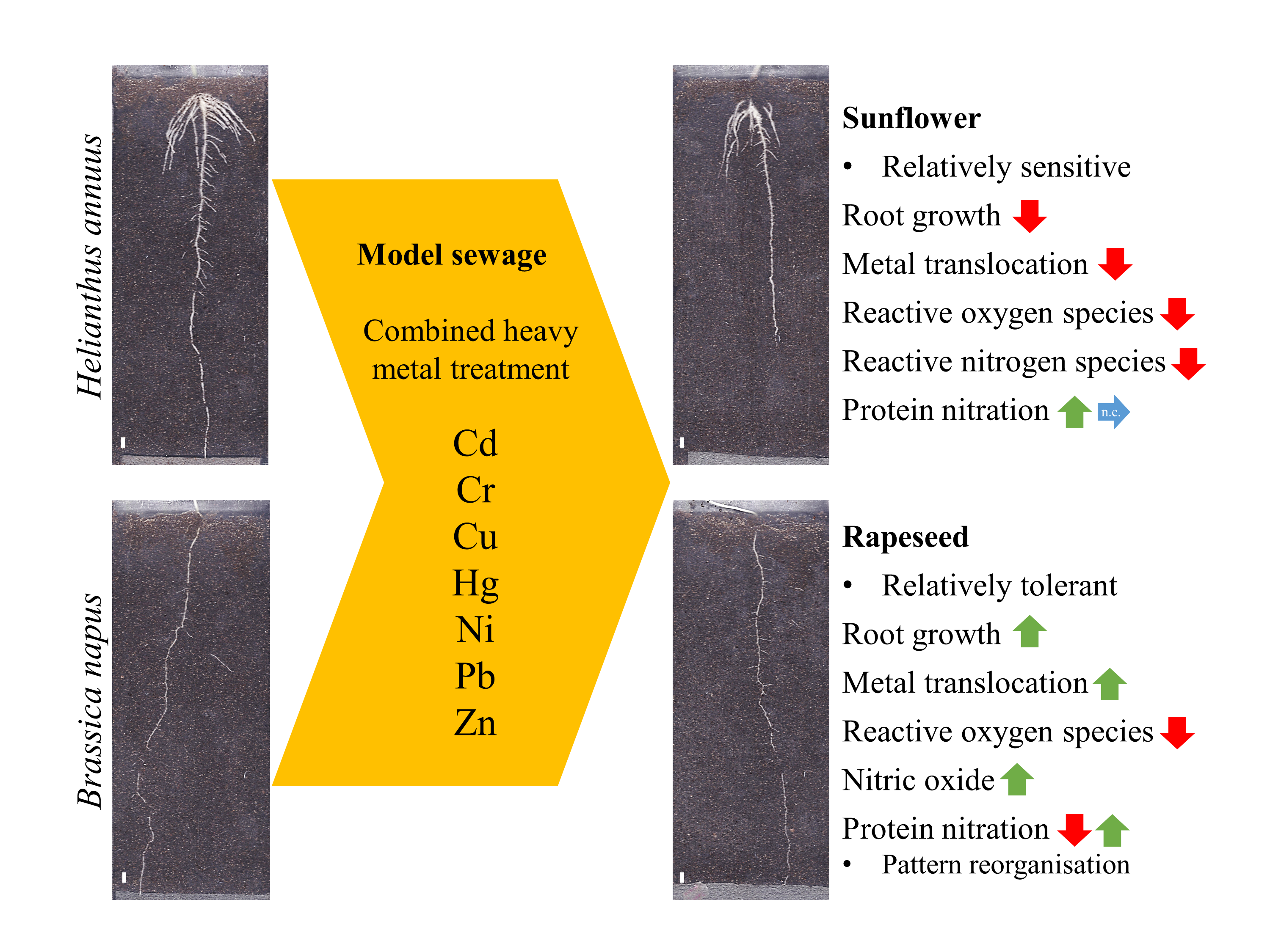
Reorganization of Protein Tyrosine Nitration Pattern Indicates the Relative Tolerance of Brassica napus (L.) over Helianthus annuus (L.) to Combined Heavy Metal Treatment

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
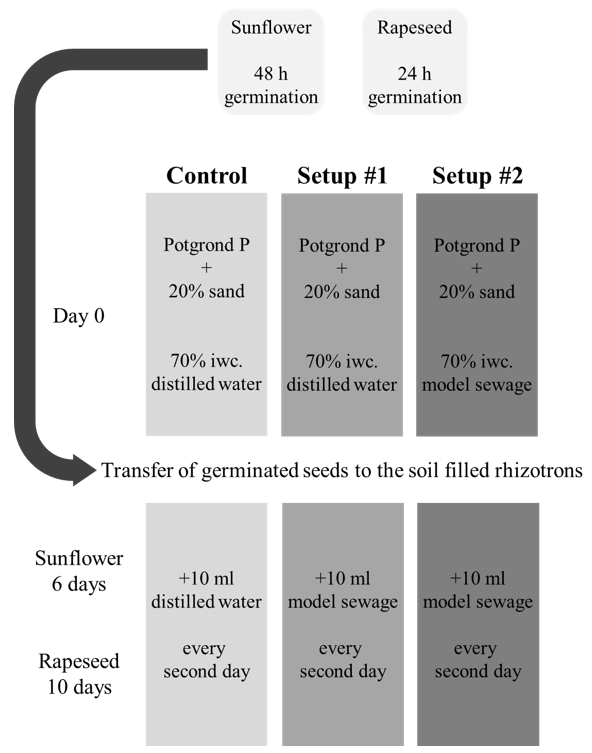
2.1. Rhizotron System, Plant Material, and Growing Conditions
2.2. Morphological Measurements
2.3. Element Content Analysis
2.4. Microscopic Determination of Callose and Pectin Deposition and Lipid Peroxidation in the Root Tissues and the Viability of the Root Apical Meristem
2.5. Detection of ROS and RNS
2.6. Acquirement and Processing of Microscopic Images
2.7. Determination of Soil Catalase Activity
2.8. SDS-PAGE and Western Blotting for Tyrosine Nitration
2.9. Statistical Analysis
3. Results and Discussion
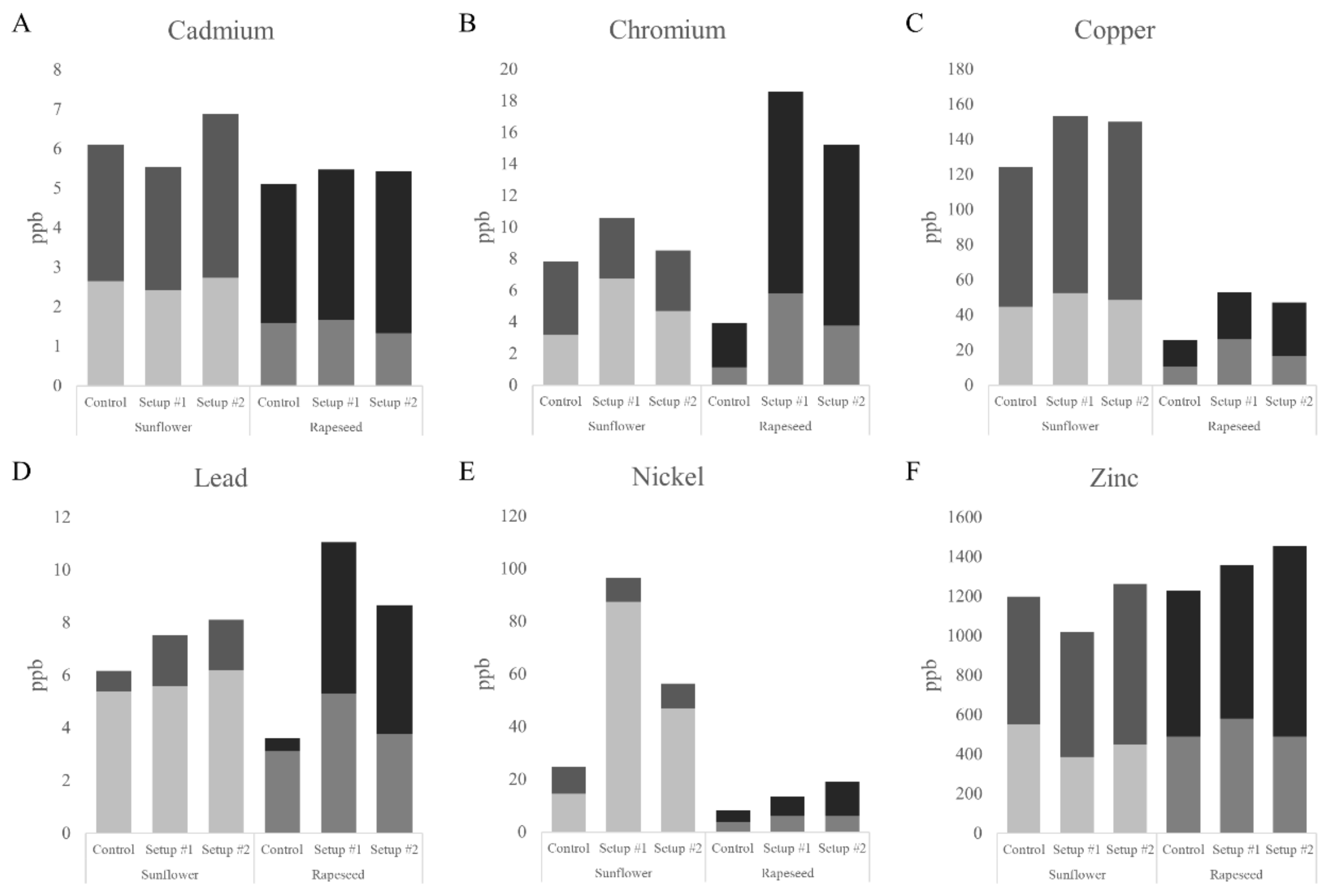
3.1. Heavy Metal Uptake
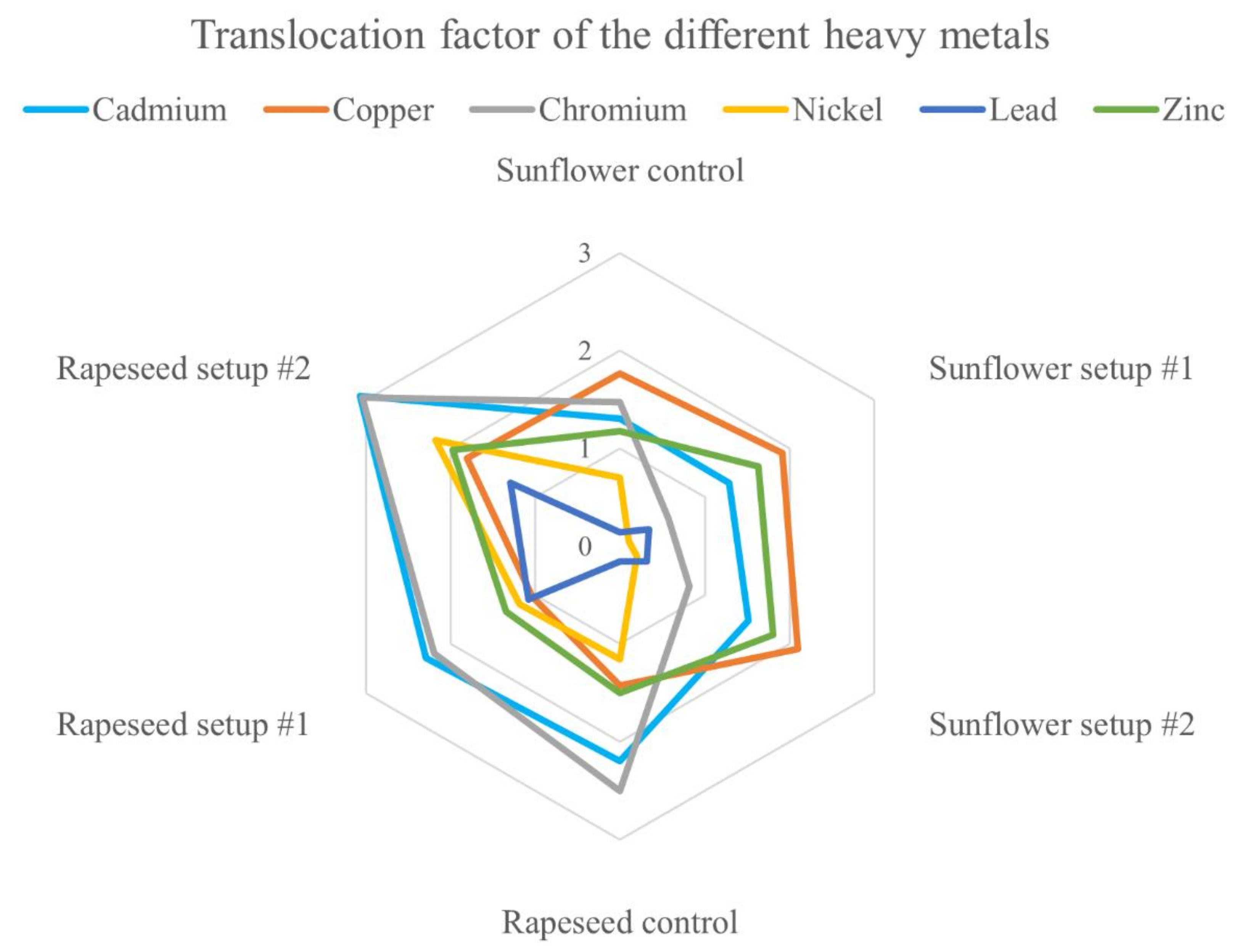
3.2. Translocation of HMs
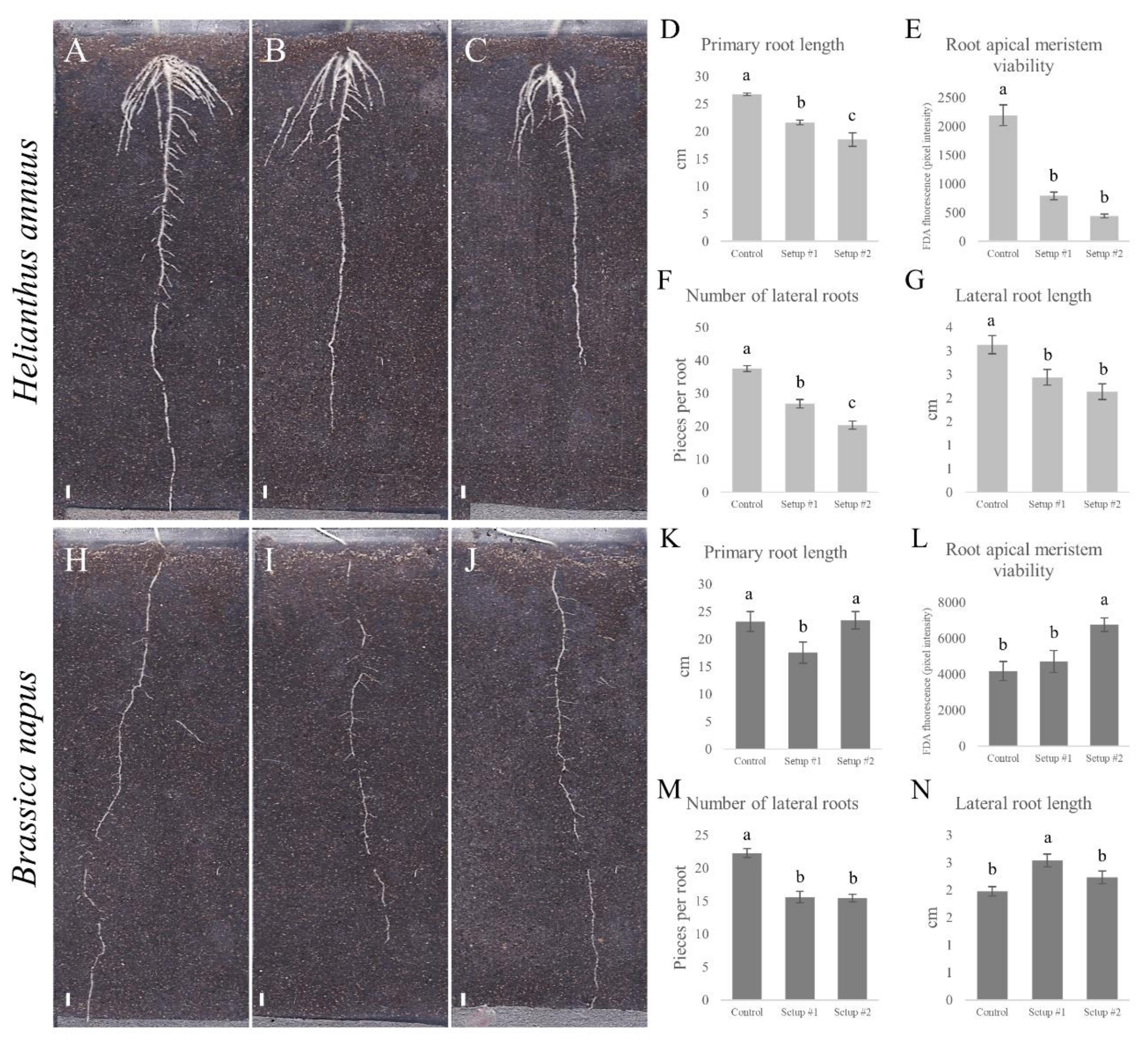
3.3. Changes in Root System Architecture
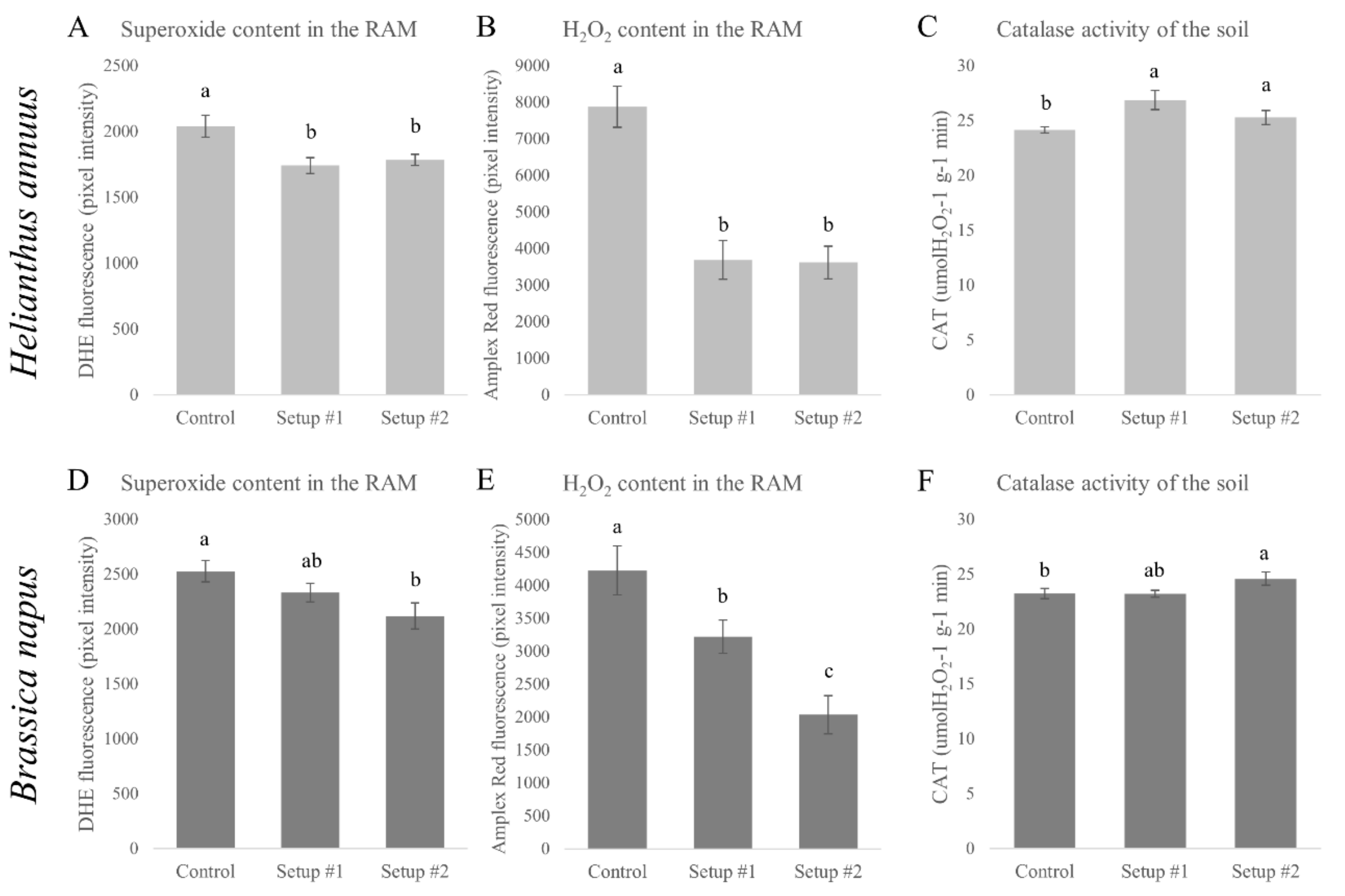
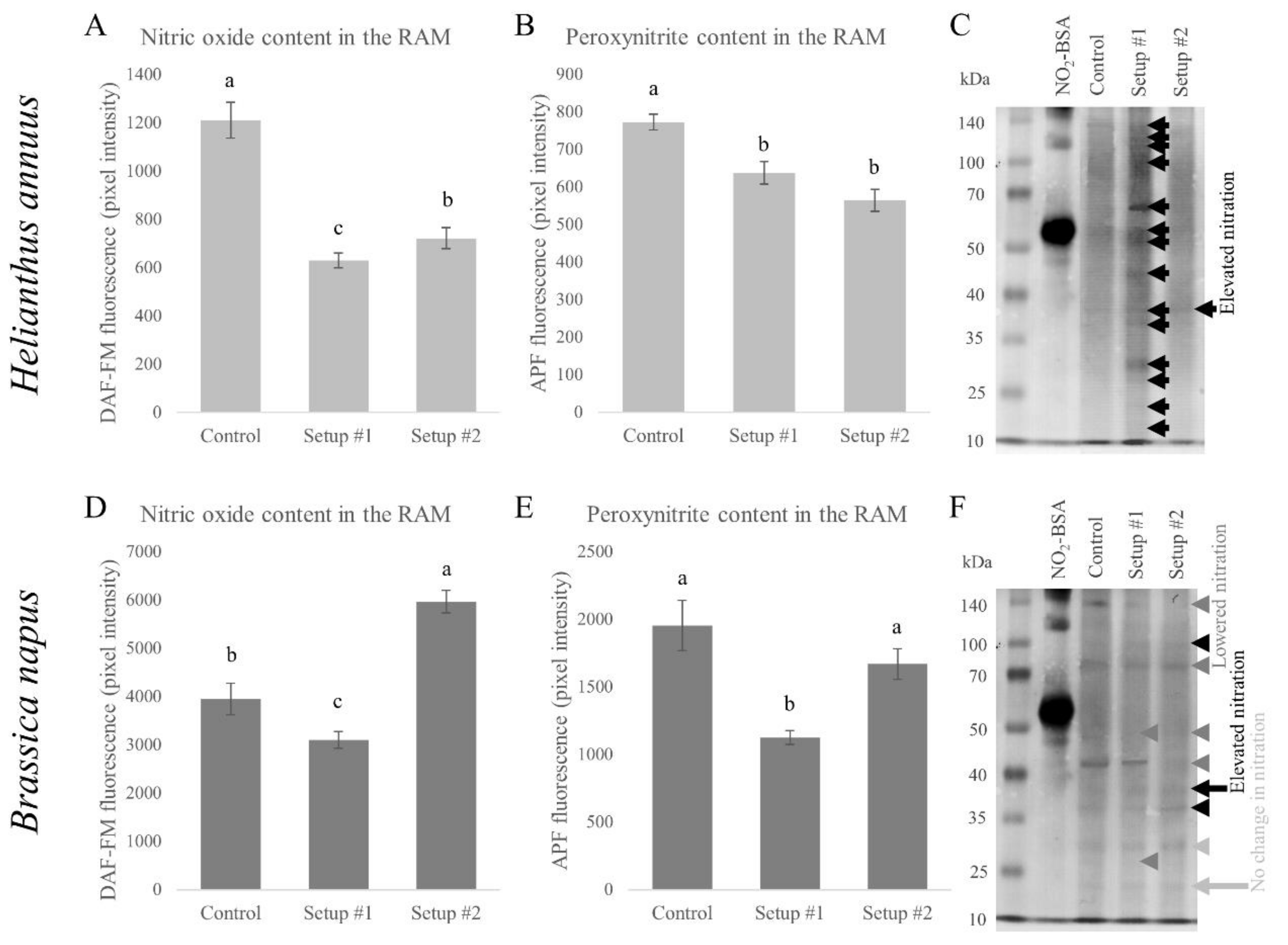
3.4. Changes in the Nitro-Oxidative Status
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Abdulaal, W.; Zeyadi, M.; Baothman, O.; Zamzami, M.; Choudhry, H.; Almulaiky, Y.; Saleh, R.; Mohamed, S. Investigation of antioxidant and detoxifying capacities of some date cultivars (Phoenix dactylifera L.) irrigated with sewage water. RSC Adv. 2017, 7, 12953–12958. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.; Ahmad, K.; Iqbal, S.; Ashfaq, A.; Bashir, H.; Mehmood, N.; Dogan, Y. Evaluation of heavy metals uptake by wheat growing in sewage water irrigated soil. Hum. Ecol. Risk Assess. Int. J. 2018, 24, 1409–1420. [Google Scholar] [CrossRef]
- Fu, F.; Wang, Q. Removal of heavy metal ions from wastewaters: A review. J. Environ. Manag. 2011, 92, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Feigl, G.; Molnár, Á.; Szőllősi, R.; Ördög, A.; Törőcsik, K.; Oláh, D.; Bodor, A.; Perei, K.; Kolbert, Z. Zinc-induced root architectural changes of rhizotron-grown B. napus correlate with a differential nitro-oxidative response. Nitric Oxide 2019, 90, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, P.; Lee, S.; Zhang, M.; Tsang, Y.; Kim, K. Heavy metals in food crops: Health risks, fate, mechanisms, and management. Environ. Int. 2019, 125, 365–385. [Google Scholar] [CrossRef] [PubMed]
- Kolbert, Z.; Oláh, D.; Molnár, Á.; Szőllősi, R.; Erdei, L.; Ördög, A. Distinct redox signalling and nickel tolerance in Brassica juncea and Arabidopsis thaliana. Ecotoxicol. Environ. Saf. 2020, 189, 109989. [Google Scholar] [CrossRef]
- Potters, G.; Pasternak, T.; Guisez, Y.; Palme, K.; Jansen, M. Stress-induced morphogenic responses: Growing out of trouble? Trends Plant Sci. 2007, 12, 98–105. [Google Scholar] [CrossRef]
- Jung, J.; McCouch, S. Getting to the roots of it: Genetic and hormonal control of root architecture. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Lamattina, L.; Spoel, S.; Loake, G. Nitric oxide function in plant biology: A redox cue in deconvolution. New Phytol. 2014, 202, 1142–1156. [Google Scholar] [CrossRef]
- Gill, S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molassiotis, A.; Fotopoulos, V. Oxidative and nitrosative signaling in plants. Plant Signal. Behav. 2011, 6, 210–214. [Google Scholar] [CrossRef] [Green Version]
- Corpas, F.; Barroso, J. Nitro-oxidative stress vs oxidative or nitrosative stress in higher plants. New Phytol. 2013, 199, 633–635. [Google Scholar] [CrossRef]
- Corpas, F.; Palma, J.; del Río, L.; Barroso, J. Protein tyrosine nitration in higher plants grown under natural and stress conditions. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krumins, J.; Goodey, N.; Gallagher, F. Plant–soil interactions in metal contaminated soils. Soil Biol. Biochem. 2015, 80, 224–231. [Google Scholar] [CrossRef]
- Hagmann, D.; Goodey, N.; Mathieu, C.; Evans, J.; Aronson, M.; Gallagher, F.; Krumins, J. Effect of metal contamination on microbial enzymatic activity in soil. Soil Biol. Biochem. 2015, 91, 291–297. [Google Scholar] [CrossRef]
- Pascual, I.; Antolín, M.; García, C.; Polo, A.; Sánchez-Díaz, M. Plant availability of heavy metals in a soil amended with a high dose of sewage sludge under drought conditions. Biol. Fertil. Soils 2004, 40, 291–299. [Google Scholar] [CrossRef]
- Kızılkaya, R.; Heps̨en, S. Effect of biosolid amendment on enzyme activities in earthworm (Lumbricus terrestris) casts. J. Plant Nutr. Soil Sci. 2004, 167, 202–208. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, J.; Wang, H.; Lin, Q.; Chen, X.; Chen, Y. The influence of soil heavy metals pollution on soil microbial biomass, enzyme activity, and community composition near a copper smelter. Ecotoxicol. Environ. Saf. 2007, 67, 75–81. [Google Scholar] [CrossRef]
- Pages, L. Mini-rhizotrons transparents pour l’étude du système racinaire de jeunes plantes. Application à la caractérisation du développement racinaire de jeunes chênes (Quercus robur). Can. J. Bot. 1992, 70, 1840–1847. [Google Scholar] [CrossRef]
- Jordan, M.; Vonarx, C. Les rhizotrons peuvent-ils être utilisés pour l’étude de la ramification des racines primaires nodales du maïs (Zea mays L)? Agronomie 1992, 12, 3–14. [Google Scholar] [CrossRef]
- Devienne-Barret, F.; Richard-Molard, C.; Chelle, M.; Maury, O.; Ney, B. Ara-rhizotron: An effective culture system to study simultaneously root and shoot development of arabidopsis. Plant Soil 2006, 280, 253–266. [Google Scholar] [CrossRef]
- Khan, I.U.; Khan, M.J.; Khan, N.U.; Khan, M.J.; Rahman, H.U.; Bibi, Z.; Ullah, K. Wastewater impact on physiology, biomass and yield of canola (Brassica napus L.). Pak. J. Bot. 2012, 44, 781–785. [Google Scholar]
- Khedkar, D.D.; Dixit, A.J. Effects of wastewater on growth pattern of Spinacea oleracea. Res. J. Env. Pollut. 2003, 2, 45–49. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehotai, N.; Kolbert, Z.; Peto, A.; Feigl, G.; Ordog, A.; Kumar, D.; Tari, I.; Erdei, L. Selenite-induced hormonal and signalling mechanisms during root growth of Arabidopsis thaliana L. J. Exp. Bot. 2012, 63, 5677–5687. [Google Scholar] [CrossRef] [Green Version]
- Rezvani, M.; Zaefarian, F. Bioaccumulation and translocation factors of cadmium and lead in ’Aeluropus littoralis’. Aust. J. Agric. Eng. 2011, 2, 114. [Google Scholar]
- Feigl, G.; Lehotai, N.; Molnár, Á.; Ördög, A.; Rodríguez-Ruiz, M.; Palma, J.; Corpas, F.; Erdei, L.; Kolbert, Z. Zinc induces distinct changes in the metabolism of reactive oxygen and nitrogen species (ROS and RNS) in the roots of two Brassica species with different sensitivity to zinc stress. Ann. Bot. 2014, 116, 613–625. [Google Scholar] [CrossRef] [Green Version]
- Durand, C.; Vicré-Gibouin, M.; Follet-Gueye, M.; Duponchel, L.; Moreau, M.; Lerouge, P.; Driouich, A. The organization pattern of root border-like cells of arabidopsis is dependent on cell wall homogalacturonan. Plant Physiol. 2009, 150, 1411–1421. [Google Scholar] [CrossRef] [Green Version]
- Lehotai, N.; Pető, A.; Bajkán, S.; Erdei, L.; Tari, I.; Kolbert, Z. In vivo and in situ visualization of early physiological events induced by heavy metals in pea root meristem. Acta Physiol. Plant. 2011, 33, 2199–2207. [Google Scholar] [CrossRef]
- Pető, A.; Lehotai, N.; Feigl, G.; Tugyi, N.; Ördög, A.; Gémes, K.; Tari, I.; Erdei, L.; Kolbert, Z. Nitric oxide contributes to copper tolerance by influencing ROS metabolism in Arabidopsis. Plant Cell Rep. 2013, 32, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Kolbert, Z.; Pető, A.; Lehotai, N.; Feigl, G.; Ördög, A.; Erdei, L. In vivo and in vitro studies on fluorophore-specificity. Acta Biol. Szeged. 2012, 56, 37–41. [Google Scholar]
- Chaki, M.; Valderrama, R.; Fernández-Ocaña, A.; Carreras, A.; López-Jaramillo, J.; Luque, F.; Palma, J.; Pedrajas, J.; Begara-Morales, J.; Sánchez-Calvo, B.; et al. Protein targets of tyrosine nitration in sunflower (Helianthus annuus L.) hypocotyls. J. Exp. Bot. 2009, 60, 4221–4234. [Google Scholar] [CrossRef]
- Stpniewska, Z.; Wolińska, A.; Ziomek, J. Response of soil catalase activity to chromium contamination. J. Environ. Sci. 2009, 21, 1142–1147. [Google Scholar] [CrossRef]
- Kolbert, Z.; Molnár, Á.; Szőllősi, R.; Feigl, G.; Erdei, L.; Ördög, A. Nitro-oxidative stress correlates with se tolerance of astragalus species. Plant Cell Physiol. 2018, 59, 1827–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hall, J. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Krzesłowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta Physiol. Plant. 2010, 33, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.; Cao, X.; Zhou, Q.; Ma, L. Accumulation of Pb, Cu, and Zn in native plants growing on a contaminated Florida site. Sci. Total Environ. 2006, 368, 456–464. [Google Scholar] [CrossRef]
- Demirezen, D.; Aksoy, A. Heavy metal levels in vegetables in Turkey are within safe limits for Cu, Zn, Ni and exceeded for Cd and Pb. J. Food Qual. 2006, 29, 252–265. [Google Scholar] [CrossRef]
- Khan, S.; Cao, Q.; Zheng, Y.; Huang, Y.; Zhu, Y. Health risks of heavy metals in contaminated soils and food crops irrigated with wastewater in Beijing, China. Environ. Pollut. 2008, 152, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Parashar, P.; Prasad, F. Study of heavy metal accumulation in sewage irrigated vegetables in different regions of agra district, india. Open J. Soil Sci. 2013, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Orisakwe, O. Evaluation of potential dietary toxicity of heavy metals of vegetables. J. Environ. Anal. Toxicol. 2012, 2. [Google Scholar] [CrossRef]
- Ashfaq, A.; Khan, Z.I.; Bibi, Z.; Ahmad, K.; Ashraf, M.; Mustafa, I.; Aisha, N.; Perveen, R.; Yasmeen, S. Heavy metals uptake by Cucurbita maxima grown in soil contaminated with sewage water and its human health implications in peri-urban areas of Sargodha city. Pak. J. Zool. 2015, 47, 1051–1058. [Google Scholar]
- Ahmad, K.; Ejaz, A.; Azam, M.; Khan, Z.I.; Ashraf, M.; Al-Qurainy, F.; Fardous, A.; Gondal, S.; Bayat, A.R.; Valeem, E.E. Lead, cadmium and chromium contents of canola irrigated with sewage water. Pak. J. Bot. 2011, 43, 1403–1410. [Google Scholar]
- Belhaj, D.; Elloumi, N.; Jerbi, B.; Zouari, M.; Abdallah, F.; Ayadi, H.; Kallel, M. Effects of sewage sludge fertilizer on heavy metal accumulation and consequent responses of sunflower (Helianthus annuus). Environ. Sci. Pollut. Res. 2016, 23, 20168–20177. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef]
- Kolbert, Z.; Feigl, G.; Bordé, Á.; Molnár, Á.; Erdei, L. Protein tyrosine nitration in plants: Present knowledge, computational prediction and future perspectives. Plant Physiol. Biochem. 2017, 113, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Chaki, M.; Begara-Morales, J.; Valderrama, R.; Mata-Pérez, C.; Padilla-Serrano, M.; Barroso, J. Role of nitric oxide–dependent posttranslational modifications of proteins under abiotic stress. Plant Life Chang. Environ. 2020, 793–809. [Google Scholar] [CrossRef]
- Gzyl, J.; Izbiańska, K.; Floryszak-Wieczorek, J.; Jelonek, T.; Arasimowicz-Jelonek, M. Cadmium affects peroxynitrite generation and tyrosine nitration in seedling roots of soybean (Glycine max L.). Environ. Exp. Bot. 2016, 131, 155–163. [Google Scholar] [CrossRef]
- Feigl, G.; Kolbert, Z.; Lehotai, N.; Molnár, Á.; Ördög, A.; Bordé, Á.; Laskay, G.; Erdei, L. Different zinc sensitivity of Brassica organs is accompanied by distinct responses in protein nitration level and pattern. Ecotoxicol. Environ. Saf. 2016, 125, 141–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnár, Á.; Papp, M.; Zoltán Kovács, D.; Bélteky, P.; Oláh, D.; Feigl, G.; Szőllősi, R.; Rázga, Z.; Ördög, A.; Erdei, L.; et al. Nitro-oxidative signalling induced by chemically synthetized zinc oxide nanoparticles (ZnO NPs) in Brassica species. Chemosphere 2020, 251, 126419. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heavy Metal | Allowed Amount (ppm) |
|---|---|
| Cadmium (Cd) | 0.02 |
| Copper (Cu) | 2.0 |
| Chromium (Cr) | 0.5 |
| Mercury (Hg) | 0.01 |
| Nickel (Ni) | 1.0 |
| Lead (Pb) | 1.0 |
| Zinc (Zn) | 5.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feigl, G.; Czifra, Á.; Molnár, Á.; Bodor, A.; Kovács, E.; Perei, K.; Jebet, V.; Kolbert, Z. Reorganization of Protein Tyrosine Nitration Pattern Indicates the Relative Tolerance of Brassica napus (L.) over Helianthus annuus (L.) to Combined Heavy Metal Treatment. Plants 2020, 9, 902. https://doi.org/10.3390/plants9070902
Feigl G, Czifra Á, Molnár Á, Bodor A, Kovács E, Perei K, Jebet V, Kolbert Z. Reorganization of Protein Tyrosine Nitration Pattern Indicates the Relative Tolerance of Brassica napus (L.) over Helianthus annuus (L.) to Combined Heavy Metal Treatment. Plants. 2020; 9(7):902. https://doi.org/10.3390/plants9070902
Chicago/Turabian StyleFeigl, Gábor, Ádám Czifra, Árpád Molnár, Attila Bodor, Etelka Kovács, Katalin Perei, Vivian Jebet, and Zsuzsanna Kolbert. 2020. "Reorganization of Protein Tyrosine Nitration Pattern Indicates the Relative Tolerance of Brassica napus (L.) over Helianthus annuus (L.) to Combined Heavy Metal Treatment" Plants 9, no. 7: 902. https://doi.org/10.3390/plants9070902