Physiological Basis of Salt Stress Tolerance in a Landrace and a Commercial Variety of Sweet Pepper (Capsicum annuum L.)

 , ,
, ,
Abstract
:1. Introduction
2. Results
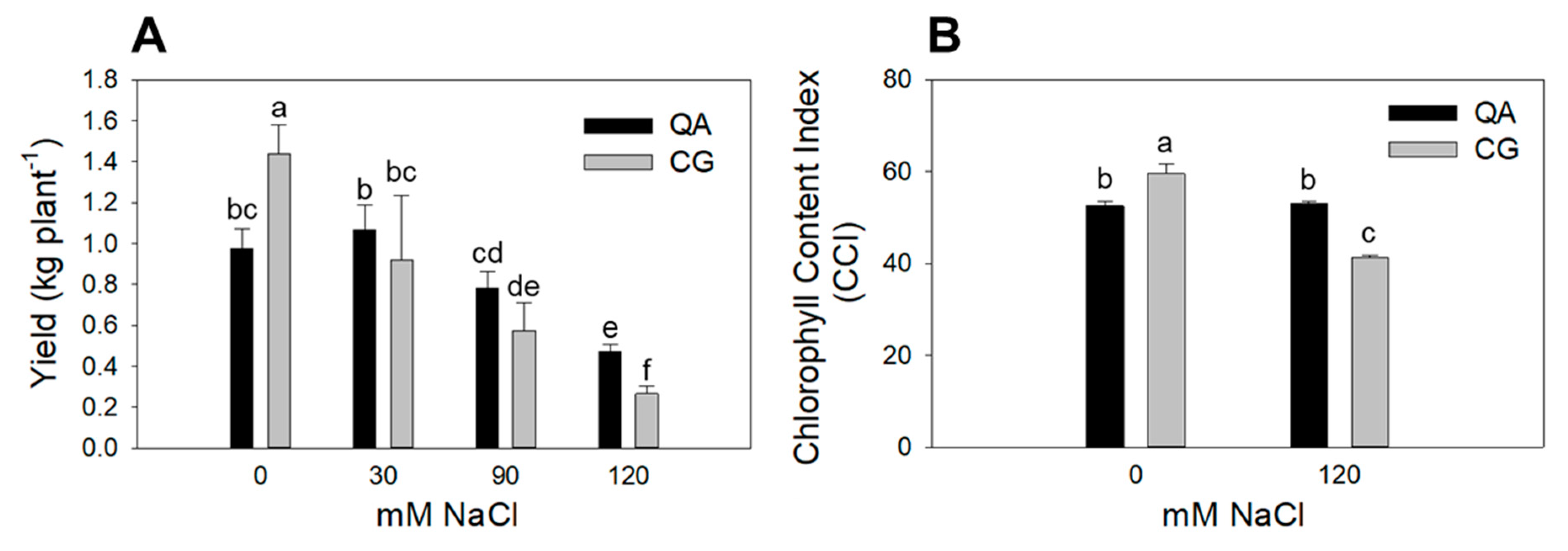
2.1. Plant Leaf Area, Yield and Chlorophyll Content Index
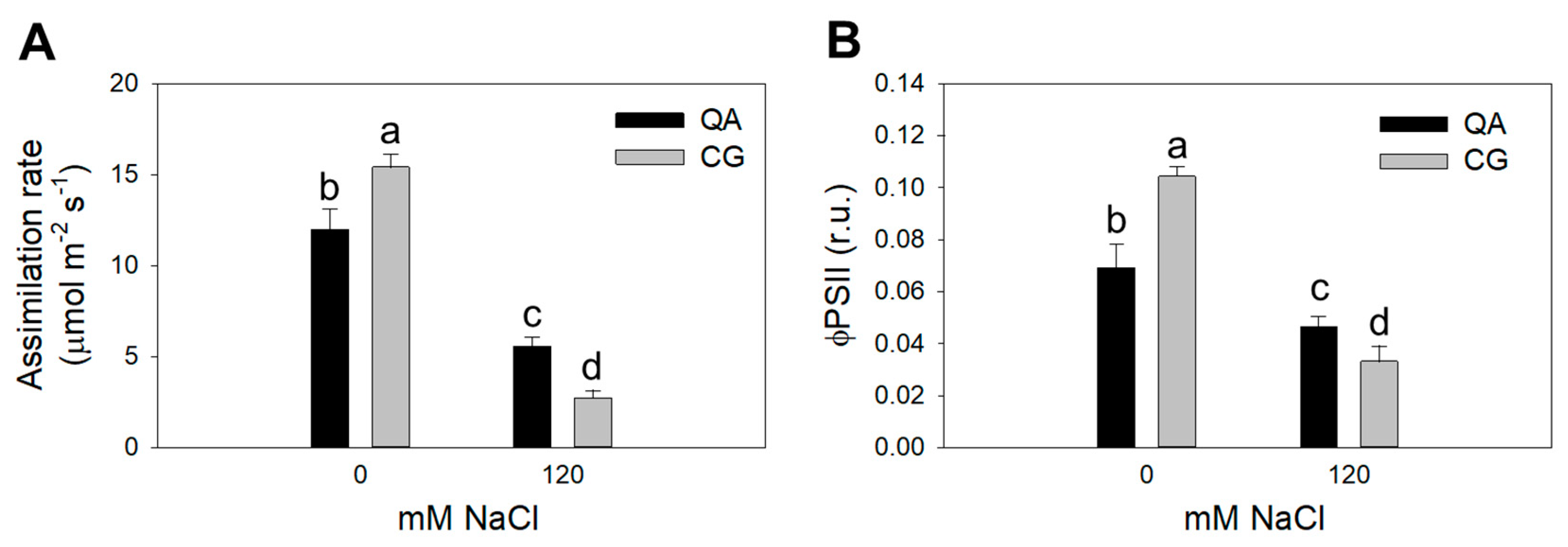
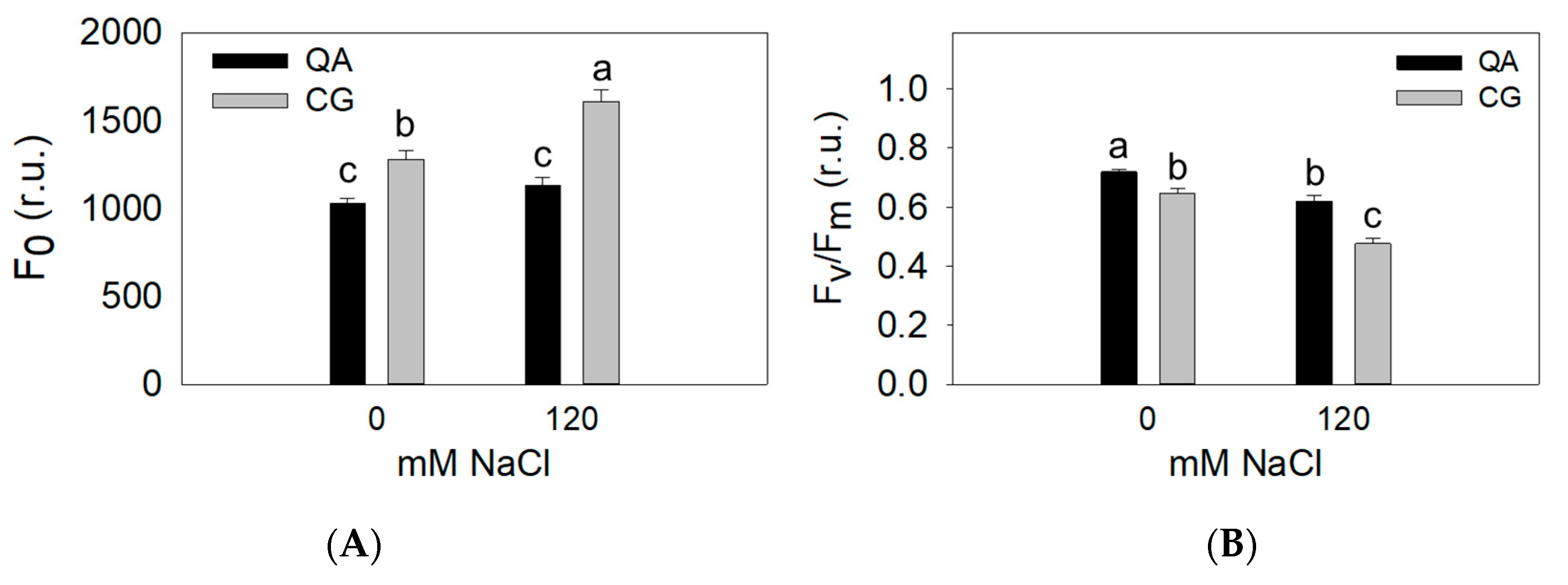
2.2. Physiological Parameters
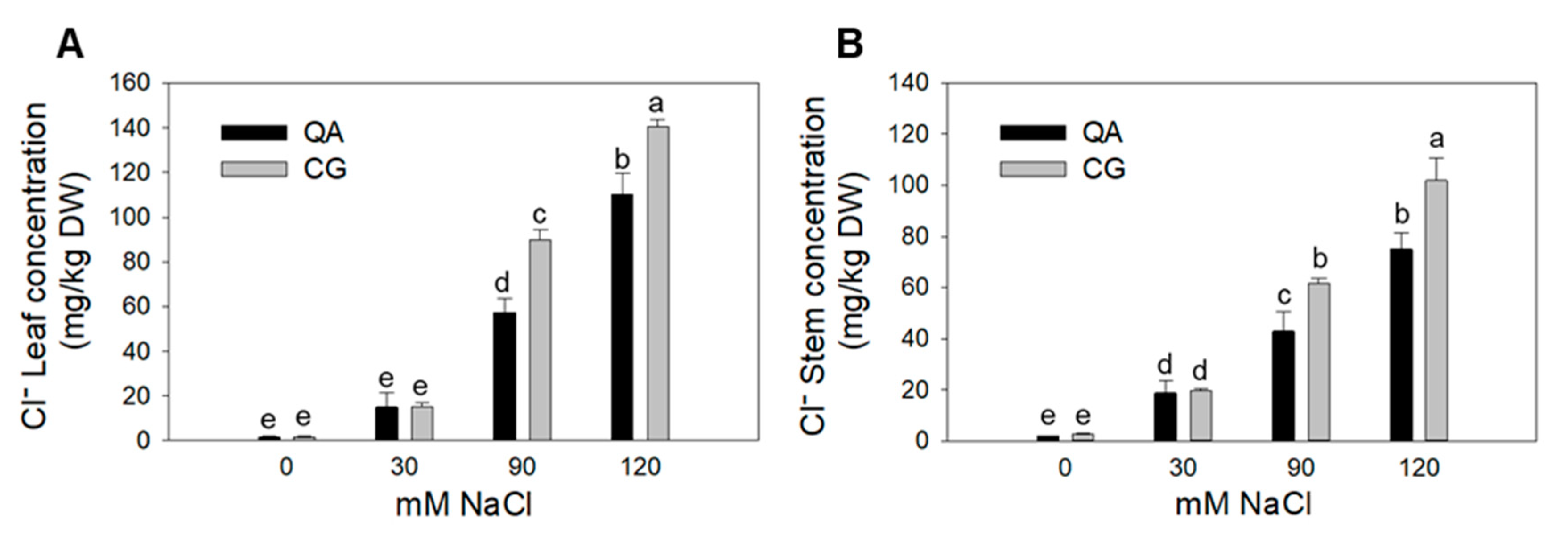
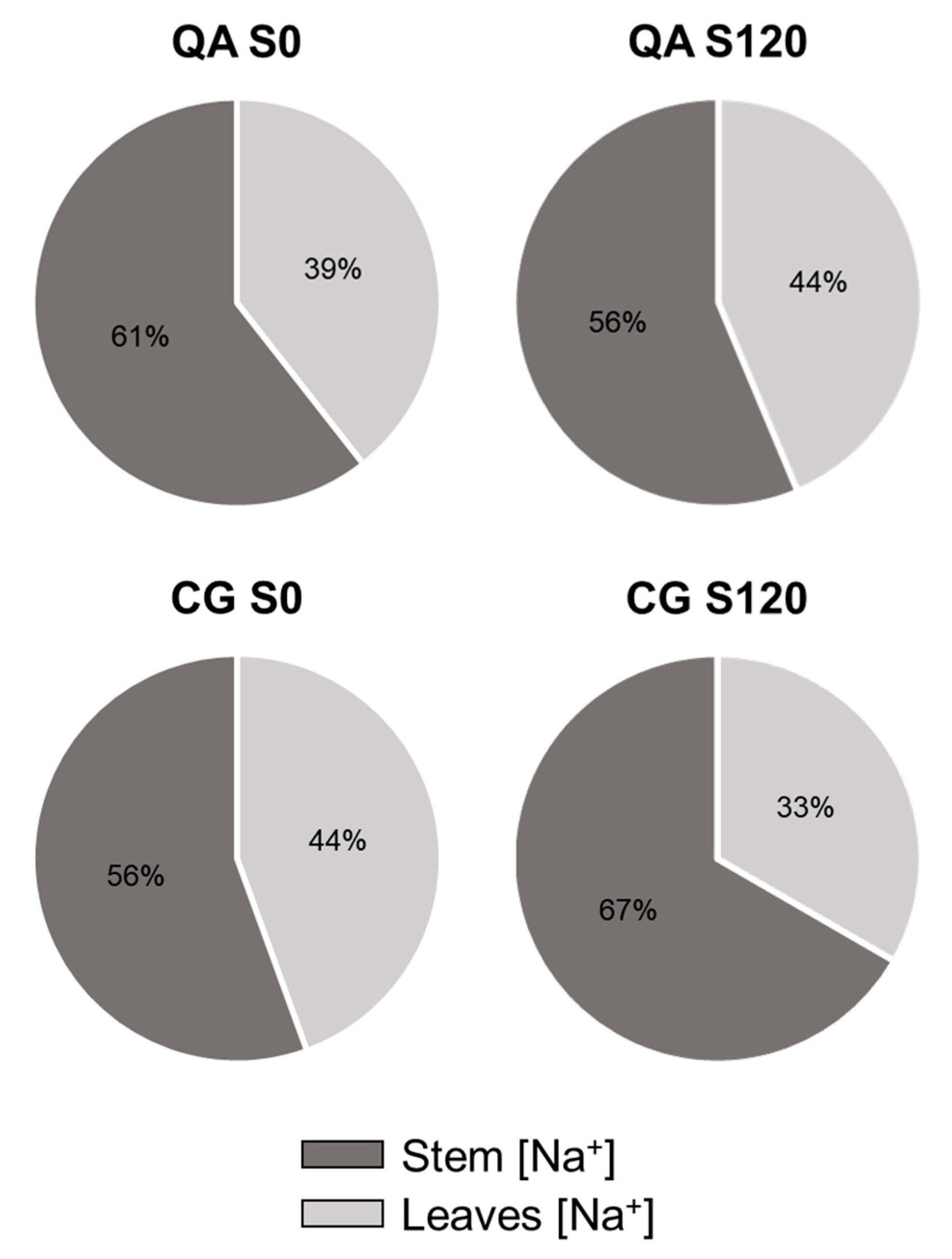
2.3. Leaf and Stem Ion Contents
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material, Growth Conditions and Salt Treatments
5.2. Growth Measurements and Yield
5.3. Plant Water Status
5.4. Leaf Gas Exchanges and Modulated Chl a Fluorescence
5.5. Transient Chl a Fluorescence Emission
5.6. Chlorophyll Content Index (CCI)
5.7. Leaf and Stem Ion Content
5.8. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Rengasamy, P. World salinization with emphasis on Australia. J. Exp. Bot. 2006, 57, 1017–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.K.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M.; Bohnert, H.J. Salt and crops: Salinity tolerance. NewsCAST News Counc. Agric. Sci. Technol. 2005, 32, 13–16. [Google Scholar]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H. Plant salt tolerance and Na+ sensing and transport. Crop J. 2018, 6, 215–225. [Google Scholar] [CrossRef]
- Niu, M.; Xie, J.; Chen, C.; Cao, H.; Sun, J.; Kong, Q.; Shabala, S.; Shabala, L.; Huang, Y.; Bie, Z. An early ABA-induced stomatal closure, Na+ sequestration in leaf vein and K+ retention in mesophyll confer salt tissue tolerance in Cucurbita species. J. Exp. Bot. 2018, 69, 4945–4960. [Google Scholar] [CrossRef] [Green Version]
- Rana, M.M.; Takamatsu, T.; Baslam, M.; Kaneko, K.; Itoh, K.; Harada, N.; Sugiyama, T.; Ohnishi, T.; Kinoshita, T.; Takagi, H.; et al. Salt tolerance improvement in rice through efficient SNP marker-assisted selection coupled with speed-breeding. Int. J. Mol. Sci. 2019, 20, 2585. [Google Scholar] [CrossRef] [Green Version]
- Teakle, N.L.; Tyerman, S.D. Mechanisms of Cl− transport contributing to salt tolerance. Plant Cell Environ. 2010, 33, 566–589. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice 2012, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuartero, J.; Bolarín, M.C.; Asíns, M.J.; Moreno, V. Increasing salt tolerance in the tomato. J. Exp. Bot. 2006, 57, 1045–1058. [Google Scholar] [CrossRef] [PubMed]
- Fita, A.; Rodríguez-Burruezo, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Breeding and domesticating crops adapted to drought and salinity: A new paradigm for increasing food production. Front. Plant Sci. 2015, 6, 978. [Google Scholar] [CrossRef] [Green Version]
- Cirillo, V.; Masin, R.; Maggio, A.; Zanin, G. Crop-weed interactions in saline environments. Eur. J. Agron. 2018, 99, 51–61. [Google Scholar] [CrossRef]
- Morton, M.J.L.; Awlia, M.; Al-Tamimi, N.; Saade, S.; Pailles, Y.; Negrão, S.; Tester, M. Salt stress under the scalpel—Dissecting the genetics of salt tolerance. Plant J. 2019, 97, 148–163. [Google Scholar] [CrossRef] [Green Version]
- Massaretto, I.L.; Albaladejo, I.; Purgatto, E.; Flores, F.B.; Plasencia, F.; Egea-Fernández, J.M.; Bolarin, M.C.; Egea, I. Recovering tomato landraces to simultaneously improve fruit yield and nutritional quality against salt stress. Front. Plant Sci. 2018, 9, 1778. [Google Scholar] [CrossRef] [Green Version]
- Razzaque, S.; Elias, S.M.; Haque, T.; Biswas, S.; Jewel, G.M.N.A.; Rahman, S.; Weng, X.; Ismail, A.M.; Walia, H.; Juenger, T.E.; et al. Gene Expression analysis associated with salt stress in a reciprocally crossed rice population. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Chartzoulakis, K.; Klapaki, G. Response of two greenhouse pepper hybrids to NaCl salinity during different growth stages. Sci. Hortic. (Amst.) 2000, 86, 247–260. [Google Scholar] [CrossRef]
- Aktas, H.; Abak, K.; Cakmak, I. Genotypic variation in the response of pepper to salinity. Sci. Hortic. (Amst.) 2006, 110, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Ruggiero, A.; Landi, S.; Punzo, P.; Possenti, M.; Van Oosten, M.J.; Costa, A.; Morelli, G.; Maggio, A.; Grillo, S.; Batelli, G. Salinity and ABA seed responses in pepper: Expression and interaction of ABA core signaling components. Front. Plant Sci. 2019, 10, 304. [Google Scholar] [CrossRef] [Green Version]
- De Pascale, S.; Orsini, F.; Caputo, R.; Palermo, M.A.; Barbieri, G.; Maggio, A. Seasonal and multiannual effects of salinisation on tomato yield and fruit quality. Funct. Plant Biol. 2012, 39, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.A. Salinity tolerance in plants: Trends and perspectives. Int. J. Mol. Sci. 2019, 20, 2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggieri, V.; Calafiore, R.; Schettini, C.; Rigano, M.M.; Olivieri, F.; Frusciante, L.; Barone, A. Exploiting genetic and genomic resources to enhance heat-tolerance in tomatoes. Agronomy 2019, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Ruggieri, V.; Francese, G.; Sacco, A.; D’Alessandro, A.; Rigano, M.M.; Parisi, M.; Milone, M.; Cardi, T.; Mennella, G.; Barone, A. An association mapping approach to identify favourable alleles for tomato fruit quality breeding. BMC Plant Biol. 2014, 14, 337. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, R. The Importance of Crop Wild Relatives, Diversity, and Genetic Potential for Adaptation to Abiotic Stress-Prone Environments. In Crop Wild Relatives and Climate Change; Wiley-Blackwell: Hoboken, NJ, USA, 2015; ISBN 9781118854396. [Google Scholar]
- Tanji, K.K.; Kielen, N.C. Crop salt tolerance data. Agric. Drain. Water Manag. Arid Semi-Arid Areas. 2002. [Google Scholar]
- Flowers, T.J.; Flowers, S.A. Why does salinity pose such a difficult problem for plant breeders? Agric. Water Manag. 2005, 78, 15–24. [Google Scholar] [CrossRef]
- Gharsallah, C.; Fakhfakh, H.; Grubb, D.; Gorsane, F. Effect of salt stress on ion concentration, proline content, antioxidant enzyme activities and gene expression in tomato cultivars. AoB Plants 2016, 8, plw055. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.P.; Dalal, R.C.; Routley, R.; Schwenke, G.D.; Daniells, I. Subsoil constraints to grain production in the cropping soils of the north-eastern region of Australia: An overview. Aust. J. Exp. Agric. 2006, 46, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Tavakkoli, E.; Fatehi, F.; Coventry, S.; Rengasamy, P.; McDonald, G.K. Additive effects of Na+ and Cl− ions on barley growth under salinity stress. J. Exp. Bot. 2011, 62, 2189–2203. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Woodrow, P.; Raimondi, G.; El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Colla, G.; Mori, M.; Giordano, M.; De Pascale, S.; et al. Omeprazole promotes chloride exclusion and induces salt tolerance in greenhouse basil. Agronomy 2019, 9, 355. [Google Scholar] [CrossRef] [Green Version]
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High concentrations of Na+ and Cl− ions in soil solution have simultaneous detrimental effects on growth of faba bean under salinity stress. J. Exp. Bot. 2010, 61, 4449–4459. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Tester, M.; Gilliham, M. Chloride on the Move. Trends Plant Sci. 2017, 22, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Orsini, F.; Alnayef, M.; Bona, S.; Maggio, A.; Gianquinto, G. Low stomatal density and reduced transpiration facilitate strawberry adaptation to salinity. Environ. Exp. Bot. 2012, 81, 1–10. [Google Scholar] [CrossRef]
- Peng, Z.; He, S.; Sun, J.; Pan, Z.; Gong, W.; Lu, Y.; Du, X. Na+ compartmentalization related to salinity stress tolerance in upland cotton (Gossypium hirsutum) seedlings. Sci. Rep. 2016, 6, 34548. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R. Chloride in soils and its uptake and movement within the plant: A review. Ann. Bot. 2001, 88, 967–988. [Google Scholar] [CrossRef] [Green Version]
- Jaarsma, R.; De Vries, R.S.M.; De Boer, A.H. Effect of Salt Stress on Growth, Na+ Accumulation and Proline Metabolism in Potato (Solanum tuberosum) Cultivars. PLoS ONE 2013, 8, e60183. [Google Scholar] [CrossRef]
- Ali, A.; Maggio, A.; Bressan, R.A.; Yun, D.J. Role and functional differences of HKT1-type transporters in plants under salt stress. Int. J. Mol. Sci. 2019, 20, 1059. [Google Scholar] [CrossRef] [Green Version]
- Britto, D.T.; Kronzucker, H.J. Sodium efflux in plant roots: What do we really know? J. Plant Physiol. 2015, 186, 1–12. [Google Scholar] [CrossRef]
- Mayrose, M.; Kane, N.C.; Mayrose, I.; Dlugosch, K.M.; Rieseberg, L.H. Increased growth in sunflower correlates with reduced defences and altered gene expression in response to biotic and abiotic stress. Mol. Ecol. 2011, 20, 4683–4694. [Google Scholar] [CrossRef]
- Weih, M. Trade-offs in plants and the prospects for breeding using modern biotechnology. New Phytol. 2003, 158, 7–9. [Google Scholar] [CrossRef]
- Venezia, A.; Tonini, A.; Piro, F.; Di Cesare, C.; Schiavi, M. Water and nutrient use efficiency of tomato soilless culture as affected by irrigation method and water quality. In Proceedings of the ISHS Acta Horticulturae 609: International Symposium on Managing Greenhouse Crops in Saline Environment, Pisa, Italy, 9–12 July 2003. [Google Scholar]
- Von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Kitajima, M.; Butler, W.L. Quenching of chlorophyll fluorescence and primary photochemistry in chloroplasts by dibromothymoquinone. BBA-Bioenerg. 1975, 376, 105–115. [Google Scholar] [CrossRef]
- Netto, A.T.; Campostrini, E.; De Oliveira, J.G.; Bressan-Smith, R.E. Photosynthetic pigments, nitrogen, chlorophyll a fluorescence and SPAD-502 readings in coffee leaves. Sci. Hortic. (Amst.) 2005, 104, 199–209. [Google Scholar] [CrossRef]
- Cirillo, V.; Van Oosten, M.J.; Izzo, M.; Maggio, A. Omeprazole treatment elicits contrasting responses to salt stress in two basil genotypes. Ann. Appl. Biol. 2019, 174, 329–338. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | LA | Height | Yield | CCI |
|---|---|---|---|---|
| (cm2) | (m) | (kg Plant−1) | (r.u.) | |
| Genotype (G) | ** | ns | ns | ns |
| QA | 4067 a | 0.76 | 0.82 | 52.9 |
| CG | 2462 b | 0.79 | 0.80 | 50.5 |
| Salinity (S) | *** | *** | *** | *** |
| S0 | 5648 a | 0.92 a | 1.20 a | 56.1 a |
| S30 | 4949 a | 1.00 a | 0.99 b | - |
| S90 | 1565 b | 0.64 b | 0.68 c | - |
| S120 | 896 b | 0.53 c | 0.37 c | 47.2 b |
| G × S | ns | ns | *** | *** |
| QA S0 | 6889 | 0.85 | 0.97 bc | 52.6 b |
| CG S0 | 4408 | 1.00 | 1.44 a | 59.6 a |
| QA S30 | 6163 | 1.00 | 1.06 b | - |
| CG S30 | 3734 | 1.01 | 0.92 bc | - |
| QA S90 | 1867 | 0.64 | 0.782 cd | - |
| CG S90 | 1263 | 0.64 | 0.575 de | - |
| QA S120 | 1347 | 0.56 | 0.473 ef | 53.1 b |
| CG S120 | 444 | 0.51 | 0.266 f | 41.3 c |
| Factors | Ψl | gs | A | ΦPSII |
|---|---|---|---|---|
| (MPa) | (mol m−2 s−1) | (µmol m−2 s−1) | (r.u.) | |
| Genotype (G) | ns | ns | ns | ns |
| QA | −0.91 | 0.19 | 8.78 | 0.060 |
| CG | −0.92 | 0.16 | 8.38 | 0.063 |
| Salinity (S) | *** | *** | *** | *** |
| 0 mM | −0.62 a | 0.37 a | 13.42 a | 0.084 a |
| 120 mM | −1.25 b | 0.05 b | 4.15 b | 0.040 b |
| G × S | ns | ns | *** | ** |
| QA S0 | −0.58 | 0.36 | 12.00 b | 0.069 b |
| CG S0 | −0.65 | 0.38 | 15.41 a | 0.104 a |
| QA S120 | −1.27 | 0.06 | 5.57 c | 0.046 c |
| CG S120 | −1.23 | 0.04 | 2.72 d | 0.033 d |
| Factors | F0 | Fv/Fm |
|---|---|---|
| (r.u.) | (r.u.) | |
| Genotype (G) | *** | *** |
| QA | 1083 b | 0.67 a |
| CG | 1445 a | 0.56 b |
| Salinity (S) | *** | *** |
| 0 mM | 1156 b | 0.68 a |
| 120 mM | 1371 a | 0.55 b |
| G × S | * | * |
| QA S0 | 1031 c | 0.72 a |
| CG S0 | 1281 b | 0.65 b |
| QA S120 | 1134 c | 0.62 b |
| CG S120 | 1269 a | 0.48 c |
| Factors | Leaves | Stem | ||
|---|---|---|---|---|
| Na+ | Cl− | Na+ | Cl− | |
| (mg kg−1 DW) | ||||
| Genotype (G) | ns | *** | ns | ** |
| QA | 6.1 | 46.0 b | 9.1 | 34.6 b |
| CG | 4.9 | 61.8 a | 11.9 | 46.5 a |
| Salinity (S) | *** | *** | *** | *** |
| 0 mM | 0.4 b | 1.7 d | 0.6 c | 2.3 d |
| 30 mM | 3.7 b | 15.0 c | 9.0 b | 19.2 c |
| 90 mM | 6.1 b | 73.5 b | 13.6 ab | 52.3 b |
| 120 mM | 11.7 a | 125.4 a | 19.0 a | 88.4 a |
| G × S | ns | * | ns | * |
| QA S0 | 0.39 | 1.6 e | 0.6 | 1.9 e |
| CG S0 | 0.48 | 1.7 e | 0.6 | 2.7 e |
| QA S30 | 3.6 | 15.0 e | 10.4 | 18.7 d |
| CG S30 | 3.8 | 15.0 e | 7.5 | 19.7 d |
| QA S90 | 8.0 | 57.3 d | 9.6 | 42.9 c |
| CG S90 | 4.2 | 89.7 c | 17.7 | 61.7 b |
| QA S120 | 12.4 | 110.1 b | 16.0 | 75.0 b |
| CG S120 | 11.0 | 140.6 a | 22.0 | 101.9 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giorio, P.; Cirillo, V.; Caramante, M.; Oliva, M.; Guida, G.; Venezia, A.; Grillo, S.; Maggio, A.; Albrizio, R. Physiological Basis of Salt Stress Tolerance in a Landrace and a Commercial Variety of Sweet Pepper (Capsicum annuum L.). Plants 2020, 9, 795. https://doi.org/10.3390/plants9060795
Giorio P, Cirillo V, Caramante M, Oliva M, Guida G, Venezia A, Grillo S, Maggio A, Albrizio R. Physiological Basis of Salt Stress Tolerance in a Landrace and a Commercial Variety of Sweet Pepper (Capsicum annuum L.). Plants. 2020; 9(6):795. https://doi.org/10.3390/plants9060795
Chicago/Turabian StyleGiorio, Pasquale, Valerio Cirillo, Martina Caramante, Marco Oliva, Gianpiero Guida, Accursio Venezia, Stefania Grillo, Albino Maggio, and Rossella Albrizio. 2020. "Physiological Basis of Salt Stress Tolerance in a Landrace and a Commercial Variety of Sweet Pepper (Capsicum annuum L.)" Plants 9, no. 6: 795. https://doi.org/10.3390/plants9060795