GmNMHC5, A Neoteric Positive Transcription Factor of Flowering and Maturity in Soybean

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Overexpression of GmNMHC5 Significantly Promoted Flowering in Soybeans
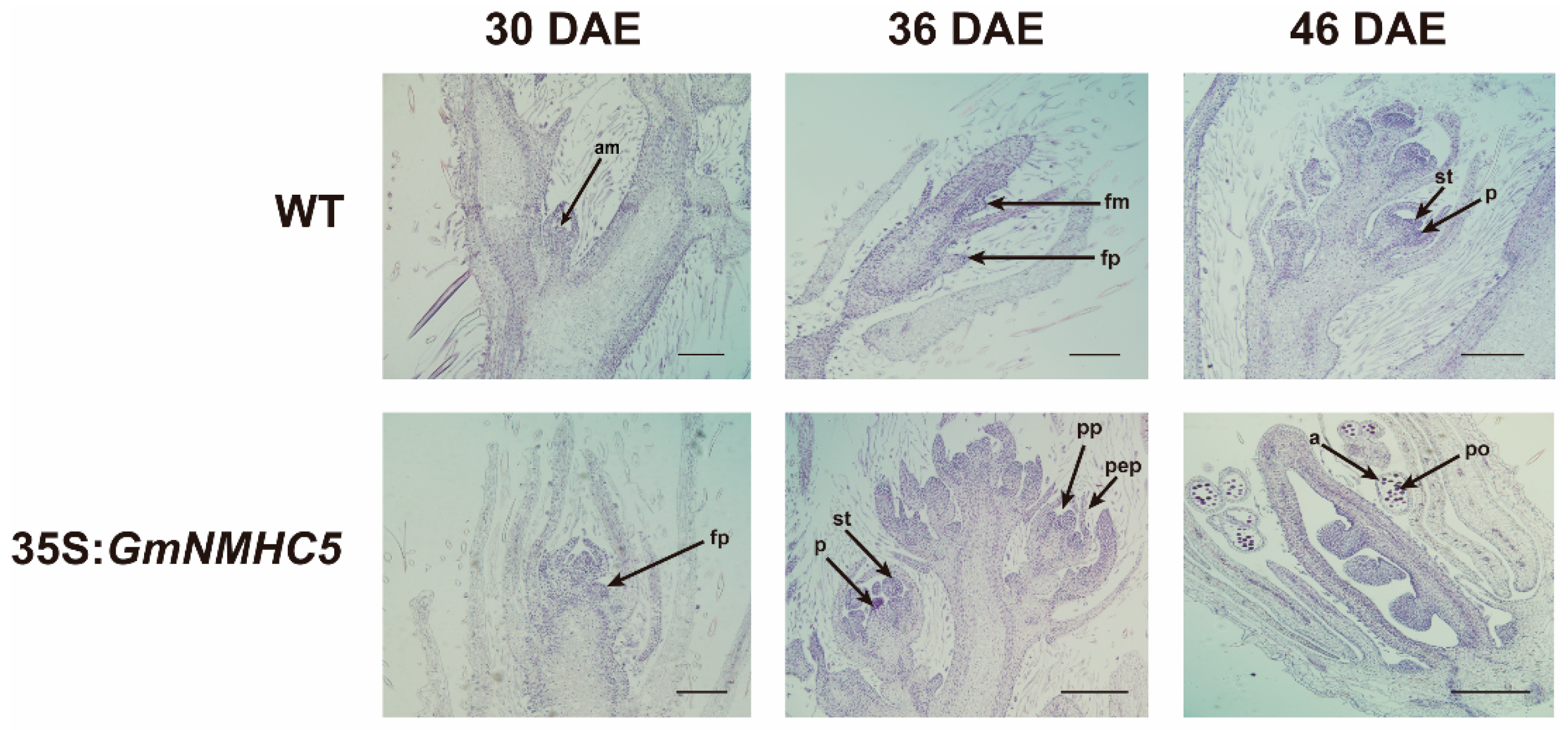
2.2. Flower Development of the 35S:GmNMHC5 Lines Occurred Earlier than That of the Wild-Type Lines
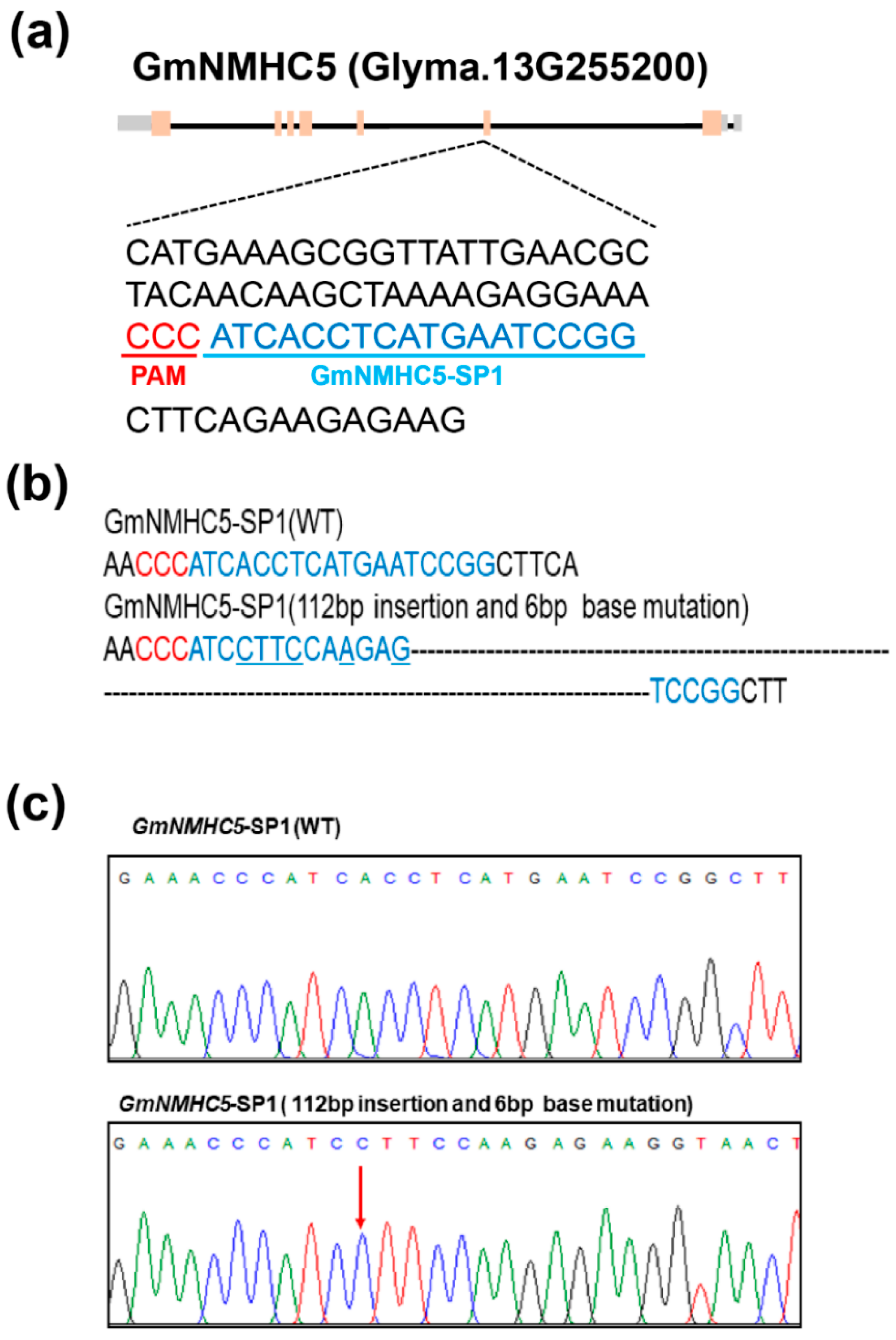
2.3. Targeted Mutagenesis of Gmnmhc5 Induced by CRISPR-Cas9
2.4. Gmnmhc5 Showed Obviously Late Flowering in Soybean
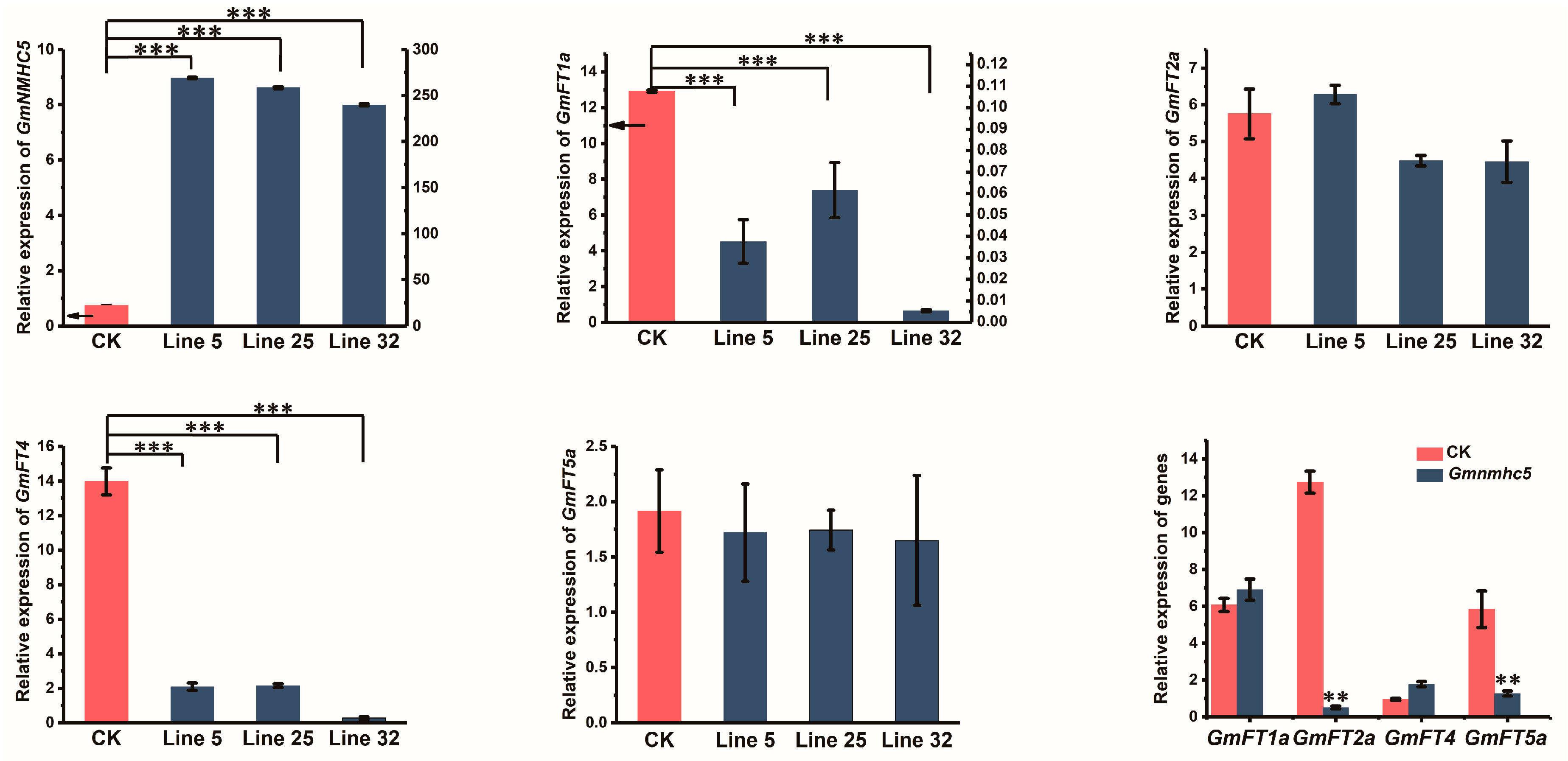
2.5. Overexpression of GmNMHC5 Inhibited the Expression of FT1a and FT4
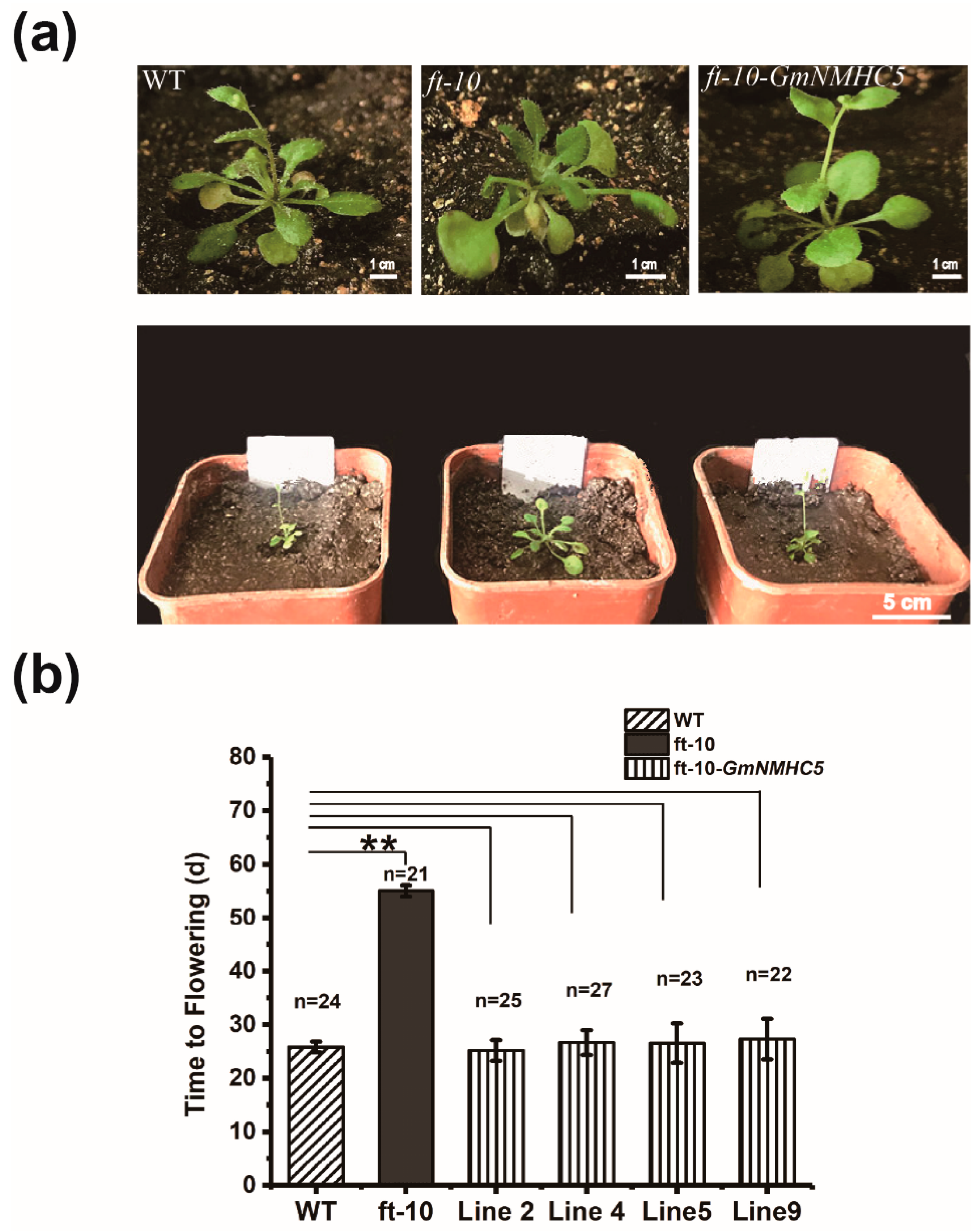
2.6. GmNMHC5 Could Rescue the Extremely Late Flowering of ft-10 in Arabidopsis
3. Discussion
4. Material and Methods
4.1. Plant Materials and Growth Conditions
4.2. Time Measurements and Statistical Analyses
4.3. Paraffin Sectioning
4.4. sgRNA Design and Construction of the CRISPR-Cas9 Expression Vector
4.5. Transformation of CRISPR-Cas9 in Soybeans and Screening for Mutations by Sequencing Analysis
4.6. Transcriptome Analysis and Gene Functional Annotation
4.7. Gene Expression Validation by qRT-PCR
4.8. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pnueli, L.; Abu-Abeid, M.; Zamir, D.; Nacken, W.; Schwarz-Sommer, Z.; Lifschitz, E. The MADS box gene family in tomato: Temporal expression during floral development, conserved secondary structures and homology with homeotic genes from Antirrhinum and Arabidopsis. Plant J. 1991, 1, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Rounsley, S.D.; Ditta, G.S.; Yanofsky, M.F. Diverse roles for MADS-box genes in Arabidopsis development. Plant Cell 1995, 7, 1259–1269. [Google Scholar] [PubMed] [Green Version]
- Ma, H.; Yanofsky, M.F.; Meyerowitz, E.M. AGL1-AGL6, an Arabidopsis gene family with similarity to floral homeotic and transcription factor genes. Genes Dev. 1991, 5, 484–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotoda, N.; Wada, M.; Kusaba, S.; Kano-Murakami, Y.; Masuda, T.; Soejima, J. Overexpression of MdMADS5, an APETALA1-like gene of apple, causes early flowering in transgenic Arabidopsis. Plant Sci. 2002, 162, 679–687. [Google Scholar] [CrossRef]
- Lee, H.; Suh, S.-S.; Park, E.; Cho, E.; Ahn, J.H.; Kim, S.-G.; Lee, J.S.; Kwon, Y.M.; Lee, I. The AGAMOUS-LIKE 20 MADS domain protein integrates floral inductive pathways in Arabidopsis. Genome Res. 2000, 14, 2366–2376. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.K.; Lee, J.S.; Ahn, J.H. Overexpression of AGAMOUS-LIKE 28 (AGL28) promotes flowering by upregulating expression of floral promoters within the autonomous pathway. Biochem. Biophys. Res. Commun. 2006, 348, 929–936. [Google Scholar] [CrossRef]
- Seo, E.; Lee, H.; Jeon, J.; Park, H.; Kim, J.; Noh, Y.-S.; Lee, I. Crosstalk between Cold Response and Flowering in Arabidopsis Is Mediated through the Flowering-Time Gene SOC1 and Its Upstream Negative Regulator FLC. Plant Cell 2009, 21, 3185–3197. [Google Scholar] [CrossRef] [Green Version]
- Adamczyk, B.J.; Fernandez, D.E.; Lehti-Shiu, M.D. The MADS domain factors AGL15 and AGL18 act redundantly as repressors of the floral transition in Arabidopsis. Plant J. 2007, 50, 1007–1019. [Google Scholar] [CrossRef]
- Ma, W.-Y.; Liu, W.; Hou, W.-S.; Sun, S.; Jiang, B.-J.; Han, T.-F.; Feng, Y.-J.; Wu, C. GmNMH7, a MADS-box transcription factor, inhibits root development and nodulation of soybean (Glycine max [L.] Merr.). J. Integr. Agric. 2019, 18, 553–562. [Google Scholar] [CrossRef]
- Gan, Y.; Filleur, S.; Rahman, A.; Gotensparre, S.; Forde, B. Nutritional regulation of ANR1 and other root-expressed MADS-box genes in Arabidopsis thaliana. Planta 2005, 222, 730–742. [Google Scholar] [CrossRef]
- Han, P.; García-Ponce, B.; Fonseca-Salazar, G.; Alvarez-Buylla, E.R.; Yu, H. AGAMOUS-LIKE 17, a novel flowering promoter, acts in a FT-independent photoperiod pathway. Plant J. 2008, 55, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Heard, J.; Caspi, M.; Dunn, K. Evolutionary Diversity of Symbiotically Induced Nodule MADS Box Genes: Characterization ofnmhC5, a Member of a Novel Subfamily. Mol. Plant Microbe Interact. 1997, 10, 665–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Han, X.; Zhan, G.; Zhao, Z.; Feng, Y.; Wu, C. A Novel Sucrose-Regulatory MADS-Box Transcription Factor GmNMHC5 Promotes Root Development and Nodulation in Soybean (Glycine max [L.] Merr.). Int. J. Mol. Sci. 2015, 16, 20657–20673. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.G.; Dean, C. Arabidopsis, the rosette stone of flowering time? Science 2002, 296, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.X.; Ma, Q.B.; Yam, K.M.; Cheung, M.Y.; Xu, Y.Y.; Han, T.F.; Lam, H.M.; Chong, K. In situ expression of the GmNMH7 gene is photoperiod-dependent in a unique soybean (Glycine max L. Merr.) flowering reversion system. Planta 2005, 223, 725–735. [Google Scholar] [CrossRef]
- Zhao, C.; Takeshima, R.; Zhu, J.; Xu, M.; Sato, M.; Watanabe, S.; Kanazawa, A.; Liu, B.; Kong, F.; Yamada, T.; et al. A recessive allele for delayed flowering at the soybean maturity locus E9 is a leaky allele of FT2a, a FLOWERING LOCUS T ortholog. BMC Plant Boil. 2016, 16, 20. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.; Nan, H.; Cao, D.; Li, Y.; Wu, F.; Wang, J.; Lu, S.; Yuan, X.; Cober, E.; Abe, J.; et al. A New Dominant Gene E9 Conditions Early Flowering and Maturity in Soybean. Crop. Sci. 2014, 54, 2529–2535. [Google Scholar] [CrossRef]
- Taoka, K.-I.; Ohki, I.; Tsuji, H.; Kojima, C.; Shimamoto, K. Structure and function of florigen and the receptor complex. Trends Plant Sci. 2013, 18, 287–294. [Google Scholar] [CrossRef]
- Corbesier, L.; Vincent, C.; Jang, S.; Fornara, F.; Fan, Q.; Searle, I.; Giakountis, A.; Farrona, S.; Gissot, L.; Turnbull, C.; et al. FT Protein Movement Contributes to Long-Distance Signaling in Floral Induction of Arabidopsis. Science 2007, 316, 1030–1033. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.; Liu, B.; Xia, Z.; Sato, S.; Kim, B.M.; Watanabe, S.; Abe, J.; Yamada, T.; Tabata, S.; Kanazawa, A.; et al. Two coordinately regulated homologues of FLOWERING LOCUS T are involved in the control of photoperiodic flowering in soybean. Plant Phys. 2010, 154, 1220–1231. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Jia, Z.; Cao, D.; Jiang, B.; Wu, C.; Hou, W.; Liu, Y.; Fei, Z.; Zhao, D.; Han, T. GmFT2a, a Soybean Homolog of FLOWERING LOCUS T, Is Involved in Flowering Transition and Maintenance. PLoS ONE 2011, 6, e29238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.P.; Chen, L.; Liu, X.J.; Guo, C.; Sun, S.; Wu, C.X.; Jiang, B.J.; Han, T.F.; Hou, W.S. CRISPR-Cas-9-mediated targeted mutagenesis of GmFT2a delays flowering time in soybean. Plant Biotechn. J. 2017, 16, 176–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, H.; Lu, S.; Liang, S.; Wu, H.; Zhang, X.; Liu, B.; Kong, F.; Yuan, X.; Li, J.; Xia, Z. GmFT4, a Homolog of FLOWERING LOCUS T, Is Positively Regulated by E1 and Functions as a Flowering Repressor in Soybean. PLoS ONE 2014, 9, e89030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Jiang, B.; Ma, L.; Zhang, S.; Zhai, H.; Xu, X.; Hou, W.; Xia, Z.; Wu, C.; Sun, S.; et al. Functional diversification of Flowering Locus T homologs in soybean: GmFT1a and GmFT2a/5a have opposite roles in controlling flowering and maturation. N. Phytol. 2017, 217, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.H.; Ito, S.; Imaizumi, T. Flowering time regulation: Photoperiod- and temperature-sensing in leaves. Trends Plant Sci. 2013, 18, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Abe, M.; Kobayashi, Y.; Yamamoto, S.; Daimon, Y.; Yamaguchi, A.; Ikeda, Y.; Ichinoki, H.; Notaguchi, M.; Goto, K.; Araki, T. FD, a bZIP Protein Mediating Signals from the Floral Pathway Integrator FT at the Shoot Apex. Science 2005, 309, 1052–1056. [Google Scholar] [CrossRef]
- Weigel, D.; Meyerowitz, E.M. The ABCs of floral homeotic genes. Cell 1994, 78, 203–209. [Google Scholar] [CrossRef]
- Chi, Y.; Huang, F.; Liu, H.; Yang, S.; Yu, D. An APETALA1-like gene of soybean regulates flowering time and specifies floral organs. J. Plant Phys. 2011, 168, 2251–2259. [Google Scholar] [CrossRef]
- Preston, J.C.; Hileman, L.C. SQUAMOSA-PROMOTER BINDING PROTEIN 1 initiates flowering in Antirrhinum majus through the activation of meristem identity genes. Plant J. 2010, 62, 704–712. [Google Scholar] [CrossRef]
- Jack, T.; Brockman, L.L.; Meyerowitz, E.M. The homeotic gene APETALA3 of Arabidopsis thaliana encodes a MADS box and is expressed in petals and stamens. Cell 1992, 68, 683–697. [Google Scholar] [CrossRef]
- Gu, Q.; Ferrándiz, C.; Yanofsky, M.F.; Martienssen, R. The FRUITFULL MADS-box gene mediates cell differentiation during Arabidopsis fruit development. Development 1998, 125, 1509–1517. [Google Scholar] [PubMed]
- Alvarez-Buylla, E.R.; Liljegren, S.J.; Pelaz, S.; Gold, S.E.; Burgeff, C.; Ditta, G.S.; Vergara-Silva, F.; Yanofsky, M.F. MADS-box gene evolution beyond flowers: Expression in pollen, endosperm, guard cells, roots and trichomes. Plant J. 2000, 24, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Zachgo, S.; Saedler, H.; Schwarz-Sommer, Z. Pollen-specific expression of DEFH125, a MADS-box transcription factor in Antirrhinum with unusual features. Plant J. 1997, 11, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Pinyopich, A.; Ditta, G.S.; Savidge, B.; Liljegren, S.J.; Baumann, E.; Wisman, E.; Yanofsky, M.F. Assessing the redundancy of MADS-box genes during carpel and ovule development. Nature 2003, 424, 85–88. [Google Scholar] [CrossRef]
- Moreno-Risueno, M.A.; Van Norman, J.M.; Moreno, A.; Zhang, J.; Ahnert, S.E.; Benfey, P.N. Oscillating Gene Expression Determines Competence for Periodic Arabidopsis Root Branching. Science 2010, 329, 1306–1311. [Google Scholar] [CrossRef] [Green Version]
- Burgeff, C.; Liljegren, S.J.; Tapia-López, R.; Yanofsky, M.F.; Alvarez-Buylla, E.R. MADS-box gene expression in lateral primordia, meristems and differentiated tissues of Arabidopsis thaliana roots. Planta 2002, 214, 365–372. [Google Scholar] [CrossRef]
- Tapia-López, R.; García-Ponce, B.; Dubrovsky, J.G.; Garay, A.; Pérez-Ruíz, R.V.; Kim, S.-H.; Acevedo, F.; Pelaz, S.; Alvarez-Buylla, E.R. An AGAMOUS-Related MADS-Box Gene, XAL1 (AGL12), Regulates Root Meristem Cell Proliferation and Flowering Transition in Arabidopsis1[W][OA]. Plant Physiol. 2008, 146, 1182–1192. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.; Suh, S.-S.; Lee, H.; Choi, K.-R.; Hong, C.B.; Paek, N.-C.; Kim, S.-G.; Lee, I. TheSOC1MADS-box gene integrates vernalization and gibberellin signals for flowering inArabidopsis. Plant J. 2003, 35, 613–623. [Google Scholar] [CrossRef]
- Borner, R.; Kampmann, G.; Wisman, E.; Apel, K.; Chandler, J.; Gleißner, R.; Melzer, S. A MADS domain gene involved in the transition to flowering in Arabidopsis. Plant J. 2000, 24, 591–599. [Google Scholar] [CrossRef]
- Turck, F.; Fornara, F.; Coupland, G. Regulation and Identity of Florigen: FLOWERING LOCUS T Moves Center Stage. Annu. Rev. Plant Boil. 2008, 59, 573–594. [Google Scholar] [CrossRef] [Green Version]
- Pin, P.A.; Benlloch, R.; Bonnet, M.; Wremerth-Weich, E.; Kraft, T.; Gielen, J.J.L.; Nilsson, O. An Antagonistic Pair of FT Homologs Mediates the Control of Flowering Time in Sugar Beet. Science 2010, 330, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Kobayashi, Y.; Goto, K.; Abe, M.; Araki, T. TWIN SISTER OF FT (TSF) Acts as a Floral Pathway Integrator Redundantly with FT. Plant Cell Physiol. 2005, 46, 1175–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clough, S.J.; Bent, A. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of Development Descriptions for Soybeans, Glycine Max (L.) Merrill 1. Crop. Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Jiang, Y.; Wu, C.; Hu, P.; Hou, W.; Han, T. Han, Morphological and anatomical characteristics of apical inflorescence development of soybean varieties with different pod bearing habits. Acta Agronom. Sin. 2014, 40, 1117–1124. [Google Scholar] [CrossRef]
- Lei, Y.; Lu, L.; Liu, H.; Li, S.; Xing, F.; Chen, L.-L. CRISPR-P: A Web Tool for Synthetic Single-Guide RNA Design of CRISPR-System in Plants. Mol. Plant 2014, 7, 1494–1496. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Hou, W.; Godo, I.; Wu, C.; Yu, Y.; Matityahu, I.; Hacham, Y.; Sun, S.; Han, T.; Amir, R. Soybean seeds expressing feedback-insensitive cystathionine ?-synthase exhibit a higher content of methionine. J. Exp. Bot. 2013, 64, 1917–1926. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Wang, Z.; Hou, W.; Chen, L.; Jiang, B.; Liu, W.; Feng, Y.; Wu, C. GmNMHC5, A Neoteric Positive Transcription Factor of Flowering and Maturity in Soybean. Plants 2020, 9, 792. https://doi.org/10.3390/plants9060792
Wang W, Wang Z, Hou W, Chen L, Jiang B, Liu W, Feng Y, Wu C. GmNMHC5, A Neoteric Positive Transcription Factor of Flowering and Maturity in Soybean. Plants. 2020; 9(6):792. https://doi.org/10.3390/plants9060792
Chicago/Turabian StyleWang, Wenting, Zhili Wang, Wensheng Hou, Li Chen, Bingjun Jiang, Wei Liu, Yongjun Feng, and Cunxiang Wu. 2020. "GmNMHC5, A Neoteric Positive Transcription Factor of Flowering and Maturity in Soybean" Plants 9, no. 6: 792. https://doi.org/10.3390/plants9060792