Photosynthetic Metabolism and Nitrogen Reshuffling Are Regulated by Reversible Cysteine Thiol Oxidation Following Nitrogen Deprivation in Chlamydomonas


Abstract
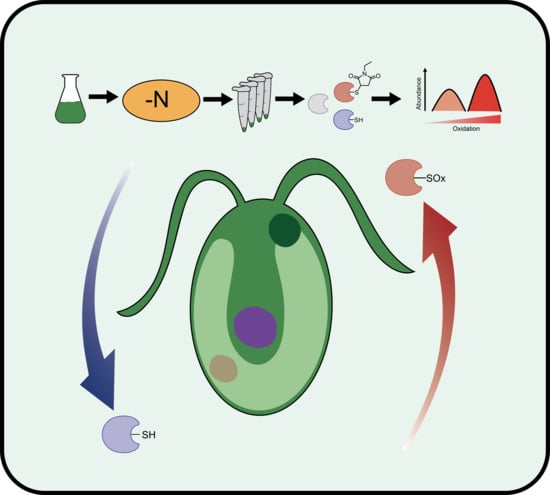
:
1. Introduction
2. Results and Discussion
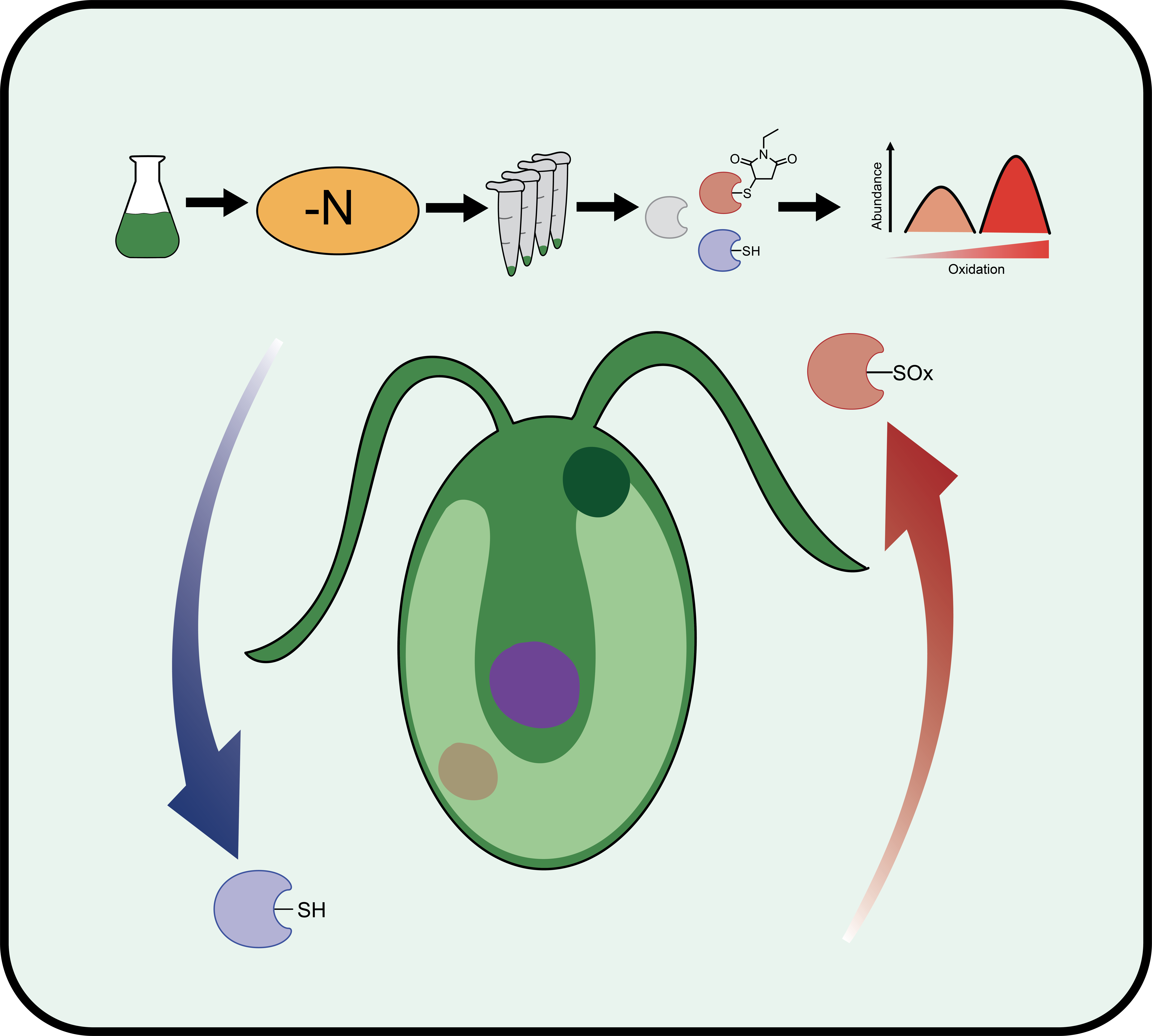
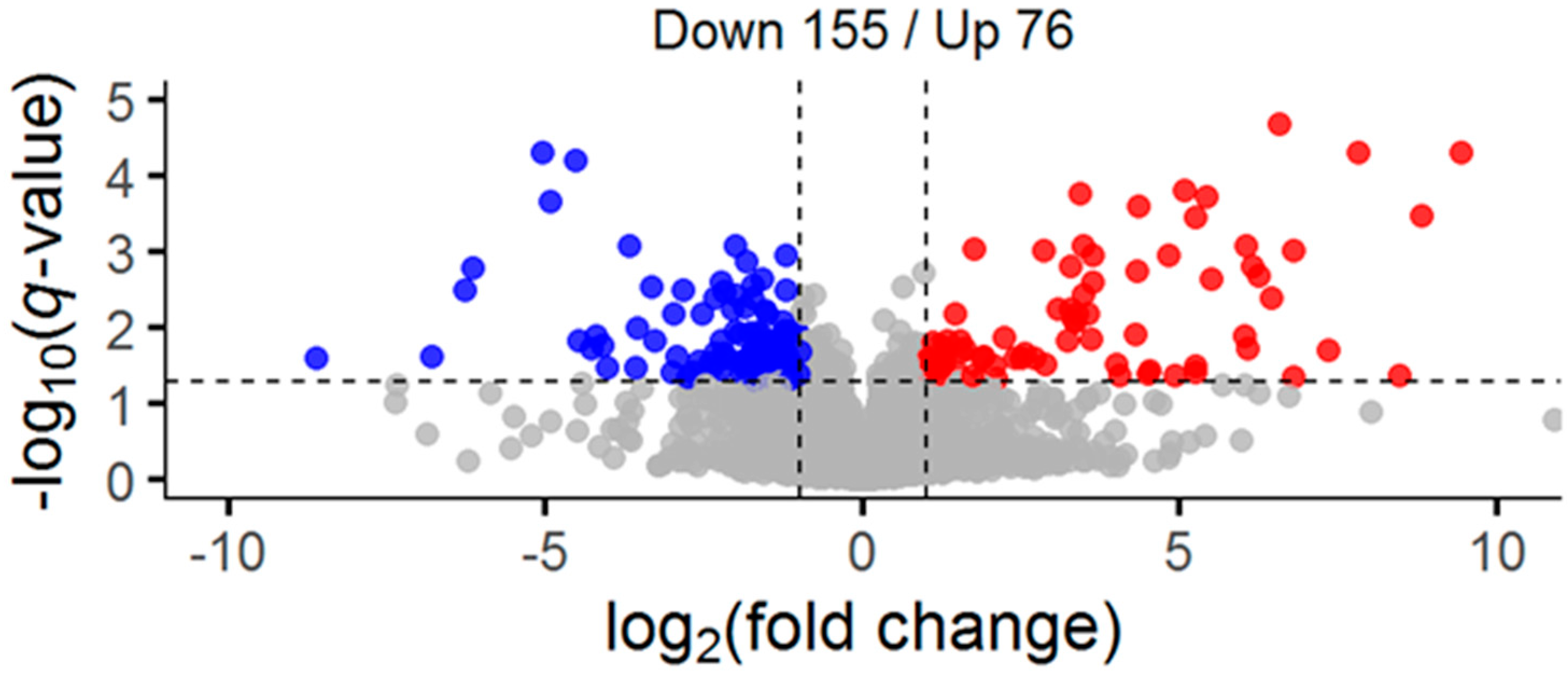
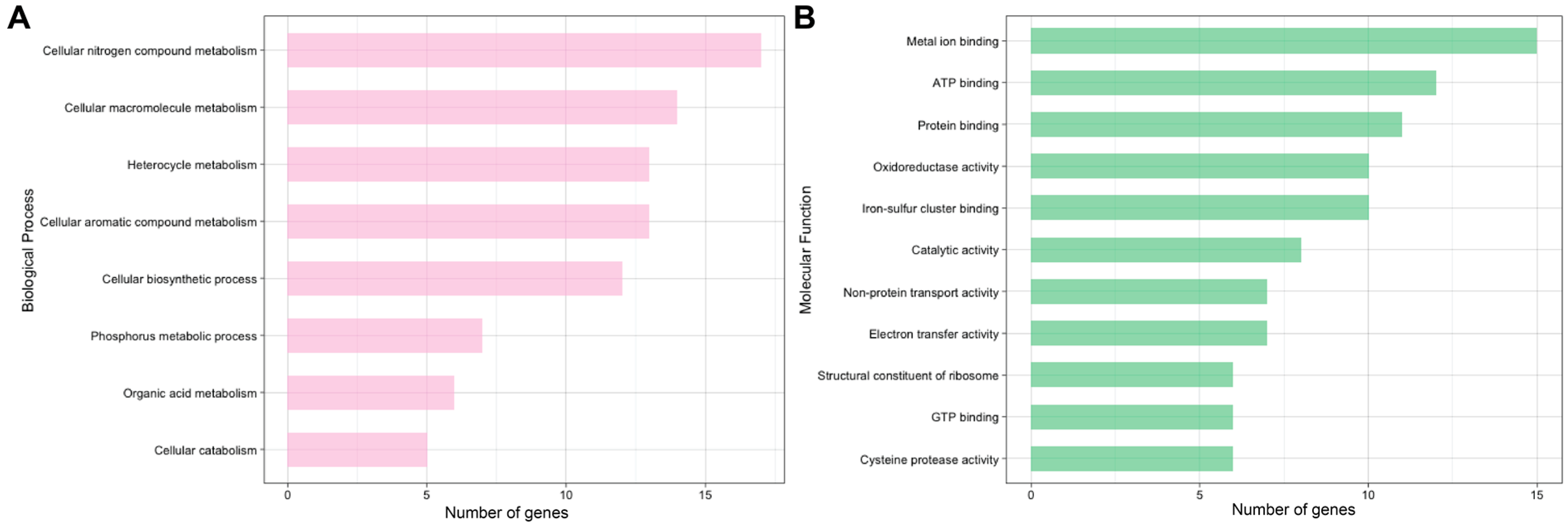
2.1. Global and Redox-Enriched Proteomic Coverage
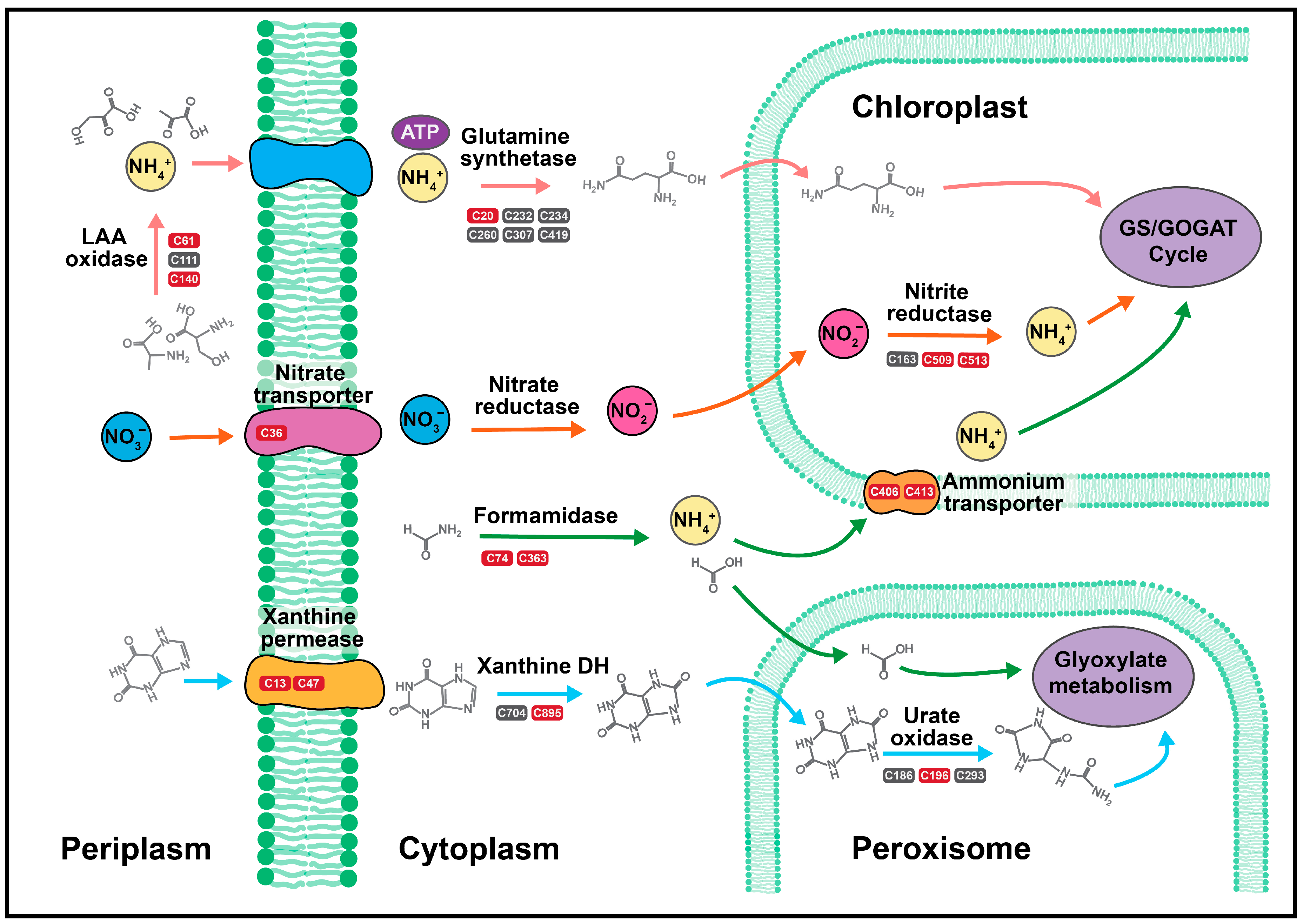
2.2. Nitrogen Assimilation and Allocation
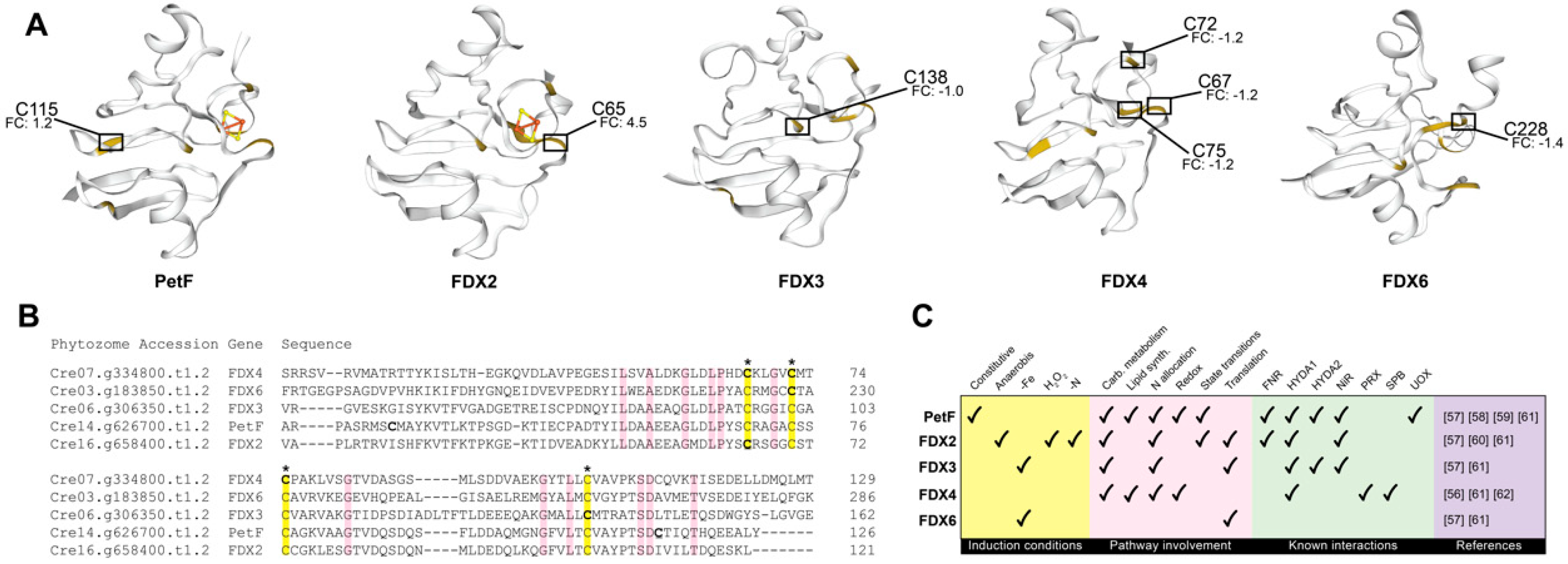
2.3. Photosynthesis and Chloroplastic Regulation
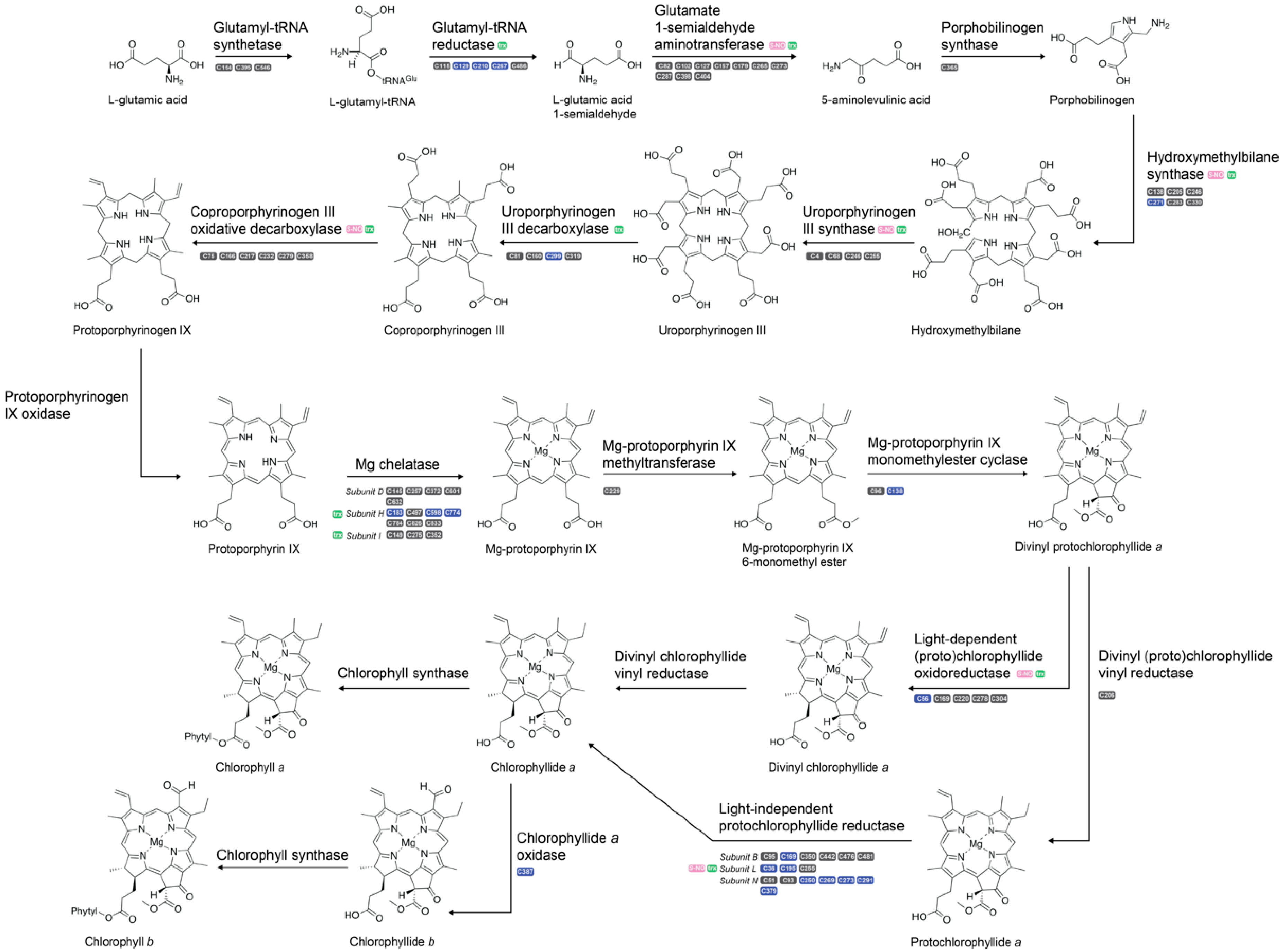
2.4. Chlorophyll Biosynthesis
2.5. Lipid Metabolism
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Cell Growth
3.3. Protein Extraction
3.4. Global Proteomics
3.5. Oxidized Cysteine Enrichment
3.6. Solid-Phase Extraction
3.7. LC–MS/MS Analysis
3.8. Database Searching and Label-Free Quantification
3.9. Data Availability
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pachauri, R.; Meyer, L. IPCC, 2014: Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2015. [Google Scholar]
- Raftery, A.E.; Zimmer, A.; Frierson, D.M.W.; Startz, R.; Liu, P. Less Than 2 °C Warming by 2100 Unlikely. Nat. Clim. Chang. 2017, 7, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Hügler, M.; Sievert, S.M. Beyond the Calvin Cycle: Autotrophic Carbon Fixation in the Ocean. Annu. Rev. Mar. Sci. 2010, 3, 261–289. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.M.; Mills, M.M.; Arrigo, K.R.; Berman-Frank, I.; Bopp, L.; Boyd, P.W.; Galbraith, E.D.; Geider, R.J.; Guieu, C.; Jaccard, S.L.; et al. Processes and patterns of oceanic nutrient limitation. Nat. Geosci. 2013, 6, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Sutherland, S.C.; Sweeney, C.; Poisson, A.; Metzl, N.; Tilbrook, B.; Bates, N.; Wanninkhof, R.; Feely, R.A.; Sabine, C.; et al. Global sea–air CO2 flux based on climatological surface ocean pCO2, and seasonal biological and temperature effects. Deep Sea Res. Part. Ii Top. Stud. Oceanogr. 2002, 49, 1601–1622. [Google Scholar] [CrossRef]
- Buchanan, P.J.; Chase, Z.; Matear, R.J.; Phipps, S.J.; Bindoff, N.L. Marine nitrogen fixers mediate a low latitude pathway for atmospheric CO2 drawdown. Nat. Commun. 2019, 10, 4611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polovina, J.J.; Howell, E.A.; Abecassis, M. Ocean’s least productive waters are expanding. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O.; Bruno, J.F. The Impact of Climate Change on the World’s Marine Ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef]
- Kromdijk, J.; Long, S.P. One crop breeding cycle from starvation? How engineering crop photosynthesis for rising CO2 and temperature could be one important route to alleviation. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152578. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.H. Chlamydomonas as a model organism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 363–406. [Google Scholar] [CrossRef] [Green Version]
- Fischer, B.B.; Wiesendanger, M.; Eggen, R.I.L. Growth Condition-Dependent Sensitivity, Photodamage and Stress Response of Chlamydomonas reinhardtii Exposed to High Light Conditions. Plant Cell Physiol. 2006, 47, 1135–1145. [Google Scholar] [CrossRef]
- Erickson, E.; Wakao, S.; Niyogi, K.K. Light stress and photoprotection in Chlamydomonas reinhardtii. Plant J. 2015, 82, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Allorent, G.; Tokutsu, R.; Roach, T.; Peers, G.; Cardol, P.; Girard-Bascou, J.; Seigneurin-Berny, D.; Petroutsos, D.; Kuntz, M.; Breyton, C.; et al. A dual strategy to cope with high light in Chlamydomonas reinhardtii. Plant Cell 2013, 25, 545–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falk, S.; Samuelsson, G.; Öquist, G. Temperature-dependent photoinhibition and recovery of photosynthesis in the green alga Chlamydomonas reinhardtii acclimated to 12 and 27 °C. Physiol. Plant 1990, 78, 173–180. [Google Scholar] [CrossRef]
- Tanaka, Y.; Nishiyama, Y.; Murata, N. Acclimation of the photosynthetic machinery to high temperature in Chlamydomonas reinhardtii requires synthesis de novo of proteins encoded by the nuclear and chloroplast genomes. Plant Physiol. 2000, 124, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Schmollinger, S.; Muhlhaus, T.; Boyle, N.R.; Blaby, I.K.; Casero, D.; Mettler, T.; Moseley, J.L.; Kropat, J.; Sommer, F.; Strenkert, D.; et al. Nitrogen-Sparing Mechanisms in Chlamydomonas Affect the Transcriptome, the Proteome, and Photosynthetic Metabolism. Plant Cell 2014, 26, 1410–1435. [Google Scholar] [CrossRef] [Green Version]
- Park, J.J.; Wang, H.; Gargouri, M.; Deshpande, R.R.; Skepper, J.N.; Holguin, F.O.; Juergens, M.T.; Shachar-Hill, Y.; Hicks, L.M.; Gang, D.R. The response of Chlamydomonas reinhardtii to nitrogen deprivation: A systems biology analysis. Plant J. Cell Mol. Biol. 2015, 81, 611–624. [Google Scholar] [CrossRef]
- Miller, R.; Wu, G.; Deshpande, R.R.; Vieler, A.; Gärtner, K.; Li, X.; Moellering, E.R.; Zäuner, S.; Cornish, A.J.; Liu, B.; et al. Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen Deprivation Predict Diversion of Metabolism. Plant Physiol. 2010, 154, 1737–1752. [Google Scholar] [CrossRef] [Green Version]
- Longworth, J.; Noirel, J.; Pandhal, J.; Wright, P.C.; Vaidyanathan, S. HILIC- and SCX-Based Quantitative Proteomics of Chlamydomonas reinhardtii during Nitrogen Starvation Induced Lipid and Carbohydrate Accumulation. J. Proteome Res. 2012, 11, 5959–5971. [Google Scholar] [CrossRef]
- Dismukes, G.C.; Carrieri, D.; Bennette, N.; Ananyev, G.M.; Posewitz, M.C. Aquatic phototrophs: Efficient alternatives to land-based crops for biofuels. Curr. Opin. Biotechnol. 2008, 19, 235–240. [Google Scholar] [CrossRef]
- McConnell, E.W.; Werth, E.G.; Hicks, L.M. The phosphorylated redox proteome of Chlamydomonas reinhardtii: Revealing novel means for regulation of protein structure and function. Redox Biol. 2018, 17, 35–46. [Google Scholar] [CrossRef]
- Werth, E.G.; McConnell, E.W.; Gilbert, T.S.; Couso Lianez, I.; Perez, C.A.; Manley, C.K.; Graves, L.M.; Umen, J.G.; Hicks, L.M. Probing the global kinome and phosphoproteome in Chlamydomonas reinhardtii via sequential enrichment and quantitative proteomics. Plant J. Cell Mol. Biol. 2017, 89, 416–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu-Rivet, E.; Scholz, M.; Arias, C.; Dardelle, F.; Schulze, S.; Le Mauff, F.; Teo, G.; Hochmal, A.K.; Blanco-Rivero, A.; Loutelier-Bourhis, C.; et al. Exploring the N-glycosylation pathway in Chlamydomonas reinhardtii unravels novel complex structures. Mol. Cell Proteom. 2013, 12, 3160–3183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.; Eikani, C.K.; Khan, H.; Iavarone, A.T.; Pesavento, J.J. Characterization of Chlamydomonas reinhardtii Core Histones by Top-Down Mass Spectrometry Reveals Unique Algae-Specific Variants and Post-Translational Modifications. J. Proteome Res. 2018, 17, 23–32. [Google Scholar] [CrossRef]
- Lemeille, S.; Turkina, M.V.; Vener, A.V.; Rochaix, J.-D. Stt7-dependent Phosphorylation during State Transitions in the Green Alga Chlamydomonas reinhardtii. Mol. Cell. Proteom. 2010, 9, 1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaffagnini, M.; Bedhomme, M.; Groni, H.; Marchand, C.H.; Puppo, C.; Gontero, B.; Cassier-Chauvat, C.; Decottignies, P.; Lemaire, S.D. Glutathionylation in the photosynthetic model organism Chlamydomonas reinhardtii: A proteomic survey. Mol. Cell Proteom. 2012, 11. [Google Scholar] [CrossRef] [Green Version]
- Morisse, S.; Zaffagnini, M.; Gao, X.-H.; Lemaire, S.D.; Marchand, C.H. Insight into protein S-nitrosylation in Chlamydomonas reinhardtii. Antioxid. Redox Signal. 2014, 21, 1271–1284. [Google Scholar] [CrossRef] [Green Version]
- Perez-Perez, M.E.; Mauries, A.; Maes, A.; Tourasse, N.J.; Hamon, M.; Lemaire, S.D.; Marchand, C.H. The Deep Thioredoxome in Chlamydomonas reinhardtii: New Insights into Redox Regulation. Mol. Plant 2017, 10, 1107–1125. [Google Scholar] [CrossRef]
- Slade, W.; Werth, E.; McConnell, E.; Alvarez, S.; Hicks, L. Quantifying Reversible Oxidation of Protein Thiols in Photosynthetic Organisms. J. Am. Soc. Mass Spectrom. 2015, 26, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Grabsztunowicz, M.; Koskela, M.M.; Mulo, P. Post-translational Modifications in Regulation of Chloroplast Function: Recent Advances. Front. Plant Sci. 2017, 8, 240. [Google Scholar] [CrossRef] [Green Version]
- Klomsiri, C.; Karplus, P.A.; Poole, L.B. Cysteine-based redox switches in enzymes. Antioxid. Redox Signal. 2011, 14, 1065–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, M.M.; Smythers, A.L.; McConnell, E.W.; Lowery, S.C.; Kolling, D.R.J.; Hicks, L.M. Inhibition of TOR in Chlamydomonas reinhardtii Leads to Rapid Cysteine Oxidation Reflecting Sustained Physiological Changes. Cells 2019, 8, 1171. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, S.; Lu, Y.; Alvarez, S.; Hicks, L.M.; Ge, X.; Xia, Y. Proteomic Analysis of Early-Responsive Redox-Sensitive Proteins in Arabidopsis. J. Proteome Res. 2012, 11, 412–424. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, H.; Wang, H.; Xia, Y. Identification of redox-sensitive cysteines in the Arabidopsis proteome using OxiTRAQ, a quantitative redox proteomics method. Proteomics 2014, 14, 750–762. [Google Scholar] [CrossRef] [PubMed]
- Strand, D.D.; Fisher, N.; Davis, G.A.; Kramer, D.M. Redox regulation of the antimycin A sensitive pathway of cyclic electron flow around photosystem I in higher plant thylakoids. Biochim. Biophys. Acta 2016, 1857, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelet, L.; Zaffagnini, M.; Vanacker, H.; Le Marechal, P.; Marchand, C.; Schroda, M.; Lemaire, S.D.; Decottignies, P. In vivo targets of S-thiolation in Chlamydomonas reinhardtii. J. Biol. Chem. 2008, 283, 21571–21578. [Google Scholar] [CrossRef] [PubMed]
- Muthuramalingam, M.; Matros, A.; Scheibe, R.; Mock, H.P.; Dietz, K.J. The hydrogen peroxide-sensitive proteome of the chloroplast in vitro and in vivo. Front. Plant Sci. 2013, 4, 54. [Google Scholar] [CrossRef] [Green Version]
- McDonagh, B. Detection of ROS Induced Proteomic Signatures by Mass Spectrometry. Front. Physiol. 2017, 8, 470. [Google Scholar] [CrossRef]
- Lemaire, S.D.; Guillon, B.; Le Maréchal, P.; Keryer, E.; Miginiac-Maslow, M.; Decottignies, P. New thioredoxin targets in the unicellular photosynthetic eukaryote Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2004, 101, 7475–7480. [Google Scholar] [CrossRef] [Green Version]
- Gargouri, M.; Park, J.-J.; Holguin, F.O.; Kim, M.-J.; Wang, H.; Deshpande, R.R.; Shachar-Hill, Y.; Hicks, L.M.; Gang, D.R. Identification of regulatory network hubs that control lipid metabolism in Chlamydomonas reinhardtii. J. Exp. Bot. 2015, 66, 4551–4566. [Google Scholar] [CrossRef] [Green Version]
- Vallon, O.; BultÉ, L.; Kuras, R.; Olive, J.; Wollman, F.-A. Extensive accumulation of an extracellular l-amino-acid oxidase during gametogenesis of Chlamydomonas reinhardtii. Eur. J. Biochem. 1993, 215, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Piedras, P.; Pineda, M.; Muñoz, J.; Cárdenas, J. Purification and characterization of an L-amino-acid oxidase from Chlamydomonas reinhardtii. Planta 1992, 188, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Luque, E.; Ocaña-Calahorro, F.; Llamas, A.; Galvan, A.; Fernandez, E. Nitric oxide controls nitrate and ammonium assimilation in Chlamydomonas reinhardtii. J. Exp. Bot. 2013, 64, 3373–3383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.; Derrien, B.; Gautier, A.; Houille-Vernes, L.; Boulouis, A.; Saint-Marcoux, D.; Malnoë, A.; Rappaport, F.; de Vitry, C.; Vallon, O.; et al. Nitric Oxide–Triggered Remodeling of Chloroplast Bioenergetics and Thylakoid Proteins upon Nitrogen Starvation in Chlamydomonas reinhardtii. Plant Cell 2014, 26, 353–372. [Google Scholar] [CrossRef] [Green Version]
- Juergens, M.T.; Deshpande, R.R.; Lucker, B.F.; Park, J.-J.; Wang, H.; Gargouri, M.; Holguin, F.O.; Disbrow, B.; Schaub, T.; Skepper, J.N.; et al. The regulation of photosynthetic structure and function during nitrogen deprivation in Chlamydomonas reinhardtii. Plant Physiol. 2015, 167, 558–573. [Google Scholar] [CrossRef] [Green Version]
- Melo, P.M.; Silva, L.S.; Ribeiro, I.; Seabra, A.R.; Carvalho, H.G. Glutamine synthetase is a molecular target of nitric oxide in root nodules of Medicago truncatula and is regulated by tyrosine nitration. Plant Physiol. 2011, 157, 1505–1517. [Google Scholar] [CrossRef] [Green Version]
- Noctor, G.; Reichheld, J.-P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.X.; Mabbitt, P.D.; Phua, S.Y.; Mueller, J.W.; Nisar, N.; Gigolashvili, T.; Stroeher, E.; Grassl, J.; Arlt, W.; Estavillo, G.M.; et al. Sensing and signaling of oxidative stress in chloroplasts by inactivation of the SAL1 phosphoadenosine phosphatase. Proc. Natl. Acad. Sci. USA 2016, 113, E4567–E4576. [Google Scholar] [CrossRef] [Green Version]
- Zaffagnini, M.; De Mia, M.; Morisse, S.; Di Giacinto, N.; Marchand, C.H.; Maes, A.; Lemaire, S.D.; Trost, P. Protein S-nitrosylation in photosynthetic organisms: A comprehensive overview with future perspectives. Biochim. Biophys. Acta 2016, 1864, 952–966. [Google Scholar] [CrossRef]
- Ballottari, M.; Alcocer, M.J.P.; D’Andrea, C.; Viola, D.; Ahn, T.K.; Petrozza, A.; Polli, D.; Fleming, G.R.; Cerullo, G.; Bassi, R. Regulation of photosystem I light harvesting by zeaxanthin. Proc. Natl. Acad. Sci. USA 2014, 111, E2431–E2438. [Google Scholar] [CrossRef] [Green Version]
- Couso, I.; Vila, M.; Vigara, J.; Cordero, B.F.; Vargas, M.Á.; Rodríguez, H.; León, R. Synthesis of carotenoids and regulation of the carotenoid biosynthesis pathway in response to high light stress in the unicellular microalga Chlamydomonas reinhardtii. Eur. J. Phycol. 2012, 47, 223–232. [Google Scholar] [CrossRef]
- Schmitter, J.M.; Jacquot, J.P.; de Lamotte-Guéry, F.; Beauvallet, C.; Dutka, S.; Gadal, P.; Decottignies, P. Purification, properties and complete amino acid sequence of the ferredoxin from a green alga, Chlamydomonas reinhardtii. Eur. J. Biochem. 1988, 172, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.m.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, A.; Winkler, M. Evolution of Chlamydomonas reinhardtii ferredoxins and their interactions with [FeFe]-hydrogenases. Photosynth. Res. 2017, 134, 307–316. [Google Scholar] [CrossRef]
- Terauchi, A.M.; Lu, S.-F.; Zaffagnini, M.; Tappa, S.; Hirasawa, M.; Tripathy, J.N.; Knaff, D.B.; Farmer, P.J.; Lemaire, S.D.; Hase, T.; et al. Pattern of Expression and Substrate Specificity of Chloroplast Ferredoxins from Chlamydomonas reinhardtii. J. Biol. Chem. 2009, 284, 25867–25878. [Google Scholar] [CrossRef] [Green Version]
- Philipps, G.; Happe, T.; Hemschemeier, A. Nitrogen deprivation results in photosynthetic hydrogen production in Chlamydomonas reinhardtii. Planta 2012, 235, 729–745. [Google Scholar] [CrossRef]
- Noth, J.; Krawietz, D.; Hemschemeier, A.; Happe, T. Pyruvate:ferredoxin oxidoreductase is coupled to light-independent hydrogen production in Chlamydomonas reinhardtii. J. Biol. Chem. 2013, 288, 4368–4377. [Google Scholar] [CrossRef] [Green Version]
- Mus, F.; Dubini, A.; Seibert, M.; Posewitz, M.C.; Grossman, A.R. Anaerobic Acclimation in Chlamydomonas reinhardtii: Anoxic Gene Expression, Hydrogenase Induction, and Metabolic Pathways. J. Biol. Chem. 2007, 282, 25475–25486. [Google Scholar] [CrossRef] [Green Version]
- Peden, E.A.; Boehm, M.; Mulder, D.W.; Davis, R.; Old, W.M.; King, P.W.; Ghirardi, M.L.; Dubini, A. Identification of Global Ferredoxin Interaction Networks in Chlamydomonas reinhardtii. J. Biol. Chem. 2013, 288, 35192–35209. [Google Scholar] [CrossRef] [Green Version]
- Baier, M.; Dietz, K.J. Protective function of chloroplast 2-cysteine peroxiredoxin in photosynthesis. Evidence from transgenic Arabidopsis. Plant Physiol 1999, 119, 1407–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, N.R.; Page, M.D.; Liu, B.; Blaby, I.K.; Casero, D.; Kropat, J.; Cokus, S.J.; Hong-Hermesdorf, A.; Shaw, J.; Karpowicz, S.J.; et al. Three acyltransferases and nitrogen-responsive regulator are implicated in nitrogen starvation-induced triacylglycerol accumulation in Chlamydomonas. J. Biol. Chem. 2012, 287, 15811–15825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Msanne, J.; Xu, D.; Konda, A.R.; Casas-Mollano, J.A.; Awada, T.; Cahoon, E.B.; Cerutti, H. Metabolic and gene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgae Chlamydomonas reinhardtii and Coccomyxa sp. C-169. Phytochemistry 2012, 75, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Kamalanathan, M.; Pierangelini, M.; Shearman, L.A.; Gleadow, R.; Beardall, J. Impacts of nitrogen and phosphorus starvation on the physiology of Chlamydomonas reinhardtii. J. Appl. Phycol. 2016, 28, 1509–1520. [Google Scholar] [CrossRef]
- Wase, N.; Black, P.N.; Stanley, B.A.; DiRusso, C.C. Integrated Quantitative Analysis of Nitrogen Stress Response in Chlamydomonas reinhardtii Using Metabolite and Protein Profiling. J. Proteome Res. 2014, 13, 1373–1396. [Google Scholar] [CrossRef] [PubMed]
- Nogaj, L.A.; Srivastava, A.; van Lis, R.; Beale, S.I. Cellular Levels of Glutamyl-tRNA Reductase and Glutamate-1-Semialdehyde Aminotransferase Do Not Control Chlorophyll Synthesis in Chlamydomonas reinhardtii. Plant Physiol. 2005, 139, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.D.; Wang, W.Y. Chlorophyll biosynthesis in Chlamydomonas starts with the formation of glutamyl-tRNA. J. Biol. Chem. 1986, 261, 13451–13455. [Google Scholar]
- Grossman, A.R.; Lohr, M.; Im, C.S. Chlamydomonas reinhardtii in the Landscape of Pigments. Annu. Rev. Genet. 2004, 38, 119–173. [Google Scholar] [CrossRef]
- Richter, A.; Grimm, B. Thiol-based redox control of enzymes involved in the tetrapyrrole biosynthesis pathway in plants. Front. Plant Sci. 2013, 4, 371. [Google Scholar] [CrossRef] [Green Version]
- Nogaj, L.A.; Beale, S.I. Physical and kinetic interactions between glutamyl-tRNA reductase and glutamate-1-semialdehyde aminotransferase of Chlamydomonas reinhardtii. J. Biol. Chem. 2005, 280, 24301–24307. [Google Scholar] [CrossRef] [Green Version]
- Richter, A.S.; Peter, E.; Rothbart, M.; Schlicke, H.; Toivola, J.; Rintamäki, E.; Grimm, B. NADPH-Dependent Thioredoxin Reductase C. Plant Physiol. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, R.; Tanaka, A. Tetrapyrrole Biosynthesis in Higher Plants. Annu. Rev. Plant Biol. 2007, 58, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, N.; Tanaka, R.; Grimm, B.; Masuda, T.; Moulin, M.; Smith, A.G.; Tanaka, A.; Terry, M.J. The cell biology of tetrapyrroles: A life and death struggle. Trends Plant Sci. 2010, 15, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liang, F.-C.; Wittmann, D.; Siegel, A.; Shan, S.-o.; Grimm, B. Chloroplast SRP43 acts as a chaperone for glutamyl-tRNA reductase, the rate-limiting enzyme in tetrapyrrole biosynthesis. Proc. Natl. Acad. Sci. USA 2018, 115, E3588–E3596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czarnecki, O.; Hedtke, B.; Melzer, M.; Rothbart, M.; Richter, A.; Schröter, Y.; Pfannschmidt, T.; Grimm, B. An Arabidopsis GluTR binding protein mediates spatial separation of 5-aminolevulinic acid synthesis in chloroplasts. Plant Cell 2011, 23, 4476–4491. [Google Scholar] [CrossRef] [Green Version]
- Willows, R.D. Biosynthesis of chlorophylls from protoporphyrin IX. Nat. Prod. Rep. 2003, 20, 327–341. [Google Scholar] [CrossRef]
- Jensen, P.E.; Reid, J.D.; Hunter, C.N. Modification of cysteine residues in the ChlI and ChlH subunits of magnesium chelatase results in enzyme inactivation. Biochem J. 2000, 352(Pt. 2), 435–441. [Google Scholar] [CrossRef]
- Fuesler, T.P.; Wong, Y.-S.; Castelfranco, P.A. Localization of Mg-chelatase and Mg-protoporphyrin IX monomethyl ester (oxidative) cyclase activities within isolated, developing cucumber chloroplasts. Plant Physiol. 1984, 75, 662–664. [Google Scholar] [CrossRef] [Green Version]
- Lankford, K.D.; Gaston, K.A.; Grovenstein, P.B.; Perera, S. Growth Phenotype and Biochemical Characterization of a Chlamydomonas reinhardtii (Green Micro-Alga) Pigment Deficient Mutant F3 (14). In Proceedings of the National Conference On Undergraduate Research (NCUR) 2012 Weber State University, Ogden, UT, USA, 29–31 March 2012. [Google Scholar]
- Fujita, Y.; Bauer, C.E. Reconstitution of light-independent protochlorophyllide reductase from purified BchL and BchN-BchB subunits in vitro confirmation of nitrogenase-like features of a bacteriochlorophyll biosynthesis enzyme. J. Biol. Chem. 2000, 275, 23583–23588. [Google Scholar] [CrossRef] [Green Version]
- Nomata, J.; Swem, L.R.; Bauer, C.E.; Fujita, Y. Overexpression and characterization of dark-operative protochlorophyllide reductase from Rhodobacter capsulatus. Biochim. Biophys. Acta (BBA) Bioenerg. 2005, 1708, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, T.; Benning, C. Nitrogen-dependent coordination of cell cycle, quiescence and TAG accumulation in Chlamydomonas. Biotechnol. Biofuels 2019, 12, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merchant, S.S.; Kropat, J.; Liu, B.; Shaw, J.; Warakanont, J. TAG, You’re it! Chlamydomonas as a reference organism for understanding algal triacylglycerol accumulation. Curr. Opin. Biotechnol. 2012, 23, 352–363. [Google Scholar] [PubMed]
- Rengel, R.; Smith, R.T.; Haslam, R.P.; Sayanova, O.; Vila, M.; León, R. Overexpression of acetyl-CoA synthetase (ACS) enhances the biosynthesis of neutral lipids and starch in the green microalga Chlamydomonas reinhardtii. Algal Res. 2018, 31, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Ramanan, R.; Kim, B.-H.; Cho, D.-H.; Ko, S.-R.; Oh, H.-M.; Kim, H.-S. Lipid droplet synthesis is limited by acetate availability in starchless mutant of Chlamydomonas reinhardtii. FEBS Lett. 2013, 587, 370–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodenough, U.; Blaby, I.; Casero, D.; Gallaher, S.D.; Goodson, C.; Johnson, S.; Lee, J.-H.; Merchant, S.S.; Pellegrini, M.; Roth, R. The path to triacylglyceride obesity in the sta6 strain of Chlamydomonas reinhardtii. Eukaryot. Cell 2014, 13, 591–613. [Google Scholar] [CrossRef] [Green Version]
- Crespo, J.L.; Díaz-Troya, S.; Florencio, F.J. Inhibition of target of rapamycin signaling by rapamycin in the unicellular green alga Chlamydomonas reinhardtii. Plant Physiol. 2005, 139, 1736–1749. [Google Scholar] [CrossRef] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas Genome Reveals the Evolution of Key Animal and Plant Functions. Science 2007, 318, 245. [Google Scholar] [CrossRef] [Green Version]
- Kall, L.; Canterbury, J.D.; Weston, J.; Noble, W.S.; MacCoss, M.J. Semi-supervised learning for peptide identification from shotgun proteomics datasets. Nat. Methods 2007, 4, 923–925. [Google Scholar] [CrossRef]
- Vizcaino, J.A.; Deutsch, E.W.; Wang, R.; Csordas, A.; Reisinger, F.; Rios, D.; Dianes, J.A.; Sun, Z.; Farrah, T.; Bandeira, N.; et al. ProteomeXchange provides globally coordinated proteomics data submission and dissemination. Nat. Biotechnol. 2014, 32, 223–226. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | UniProt ID | Gene | Protein | Cys Site(s) | log2FC |
|---|---|---|---|---|---|
| Cre02.g113200.t1.1 | A0A2K3E350 | GLN1 | Glutamine synthetase | C20 | 5.2 |
| Cre09.g410750.t1.2 | Q9ZR67 | NII1 | Nitrite Reductase | C509, C513 | 4.3 |
| Cre09.g410850.t1.2 | A0A2K3DFM1 | NRT2.1 | Nitrate transporter | C36 | 6.0 |
| Cre10.g442800.t1.1 | A8II65 | XUV6 | Xanthine/uracil permease | C13 | 5.5 |
| C47 | 8.5 | ||||
| Cre12.g504950.t1.2 | A0A2K3D311 | UOX1 | Urate oxidase | C196 | 1.3 |
| Cre12.g545101.t1.1 | A0A2K3D6K5 | XDH1 | Xanthine dehydrogenase | C895 | 4.3 |
| Cre12.g551352.t1.1 | A0A2K3D647 | LAO1 | L-amino acid oxidase | C140 | 6.4 |
| Cre12.g551353.t1.1 | A0A2K3D651 | LAO1 | L-amino acid oxidase | C61 | 6.1 |
| Cre13.g569850.t1.2 | A8HSA2 | AMT4 | Ammonium transporter | C406 | 6.8 |
| C413 | 4.3 | ||||
| Cre16.g691750.t1.2 | A8JBG4 | AMI1 | Formamidase | C74 | 7.8 |
| C363 | 3.4 |
| Accession | UniProt ID | Gene | Protein | Cys Site(s) | log2FC |
|---|---|---|---|---|---|
| Cre01.g052250.t1.2 | Q84XR9 | TRXx | Thioredoxin x | C24 | −4.3 |
| Cre02.g082550.t1.2 | Q84U73 | ZEP1 | Zeaxanthin epoxidase | C250 | −1.3 |
| Cre03.g183850.t1.2 | Q2HZ21 | FDX6 | Ferredoxin | C228 | −1.4 |
| Cre03.g199800.t1.1 | Q9FYU1 | HYDA1 | Fe-hydrogenase | C88 | −1.6 |
| C225 | −2.2 | ||||
| C238 | −2.6 | ||||
| Cre06.g272650.t1.2 | Q75VY7 | LHCA8 | Chlorophyll a-b binding protein | C24 | −1.3 |
| Cre06.g306350.t1.2 | Q2HZ24 | FDX3 | Ferredoxin | C138 | −1.0 |
| Cre07.g334800.t1.2 | Q2HZ23 | FDX4 | Ferredoxin | C67, C72, C75 | −1.2 |
| Cre14.g626700.t1.2 | A8IV40 | PetF | Ferredoxin | C30 | −1.6 |
| C115 | 1.2 | ||||
| Cre16.g658400.t1.2 | A0A2K3CTD8 | FDX2 | Ferredoxin | C65 | 4.5 |
| Accession | UniProt ID | Gene | Protein | Cys Site(s) | log2FC |
|---|---|---|---|---|---|
| Cre01.g015350.t1.1 | Q39617 | POR1 | Light-dependent protochlorophyllide reductase | C56 | −4.5 |
| Cre01.g015500.t1.2 | A8HPJ5 | CGL91 | Predicted signaling protein | C10 | 1.3 |
| Cre01.g043350.t1.2 | Q9ZWM5 | CAO1 | Chlorophyll a oxygenase | C387 | −2.3 |
| Cre01.g050950.t1.2 | A8HNE8 | Unknown | Geranylgeranyl reductase | C115, C118, C125 | −1.3 |
| Cre02.g142266.t1.1 | A5YU14 | Unknown | CYP97A5 | C231 | −2.9 |
| Cre07.g325500.t1.1 | A8I7P5 | CHLH1 | Magnesium chelatase subunit H | C183 | −1.8 |
| C598 | −1.7 | ||||
| C774 | −1.8 | ||||
| Cre07.g342150.t1.2 | Q9FPR7 | HEMA1 | Glutamyl-tRNA reductase | C129 | −1.7 |
| C210 | −1.8 | ||||
| C267 | −1.2 | ||||
| Cre07.g346050.t1.2 | Q9LD46 | CRD1 | Magnesium-protoporphyrin IX monomethyl ester cyclase | C138 | −2.7 |
| Cre11.g467700.t1.1 | A8JC21 | UROD1 | Uroporphyrinogen III decarboxylase | C299 | −1.0 |
| Cre12.g488350.t1.1 | A0A2K3D2A6 | Unknown | Uncharacterized protein | C578 | −1.1 |
| Cre16.g652000.t1.1 | A0A2K3CSX1 | Unknown | Carotenoid isomerase | C42 | −1.5 |
| Cre16.g663900.t1.2 | A8JFB1 | PBGD1 | Porphobilinogen deaminase | C271 | −1.7 |
| NP_958412.2 | P29683 | chlN | Light-independent protochlorophyllide reductase subunit N | C250 | −1.2 |
| C269, C273 | −1.3 | ||||
| C273 | −1.2 | ||||
| C291 | −1.0 | ||||
| C379 | −1.4 | ||||
| NP_958360.1 | P36437 | chlB | Light-independent protochlorophyllide reductase subunit B | C169 | −1.0 |
| NP_958366.1 | Q00469 | chlL | Light-independent protochlorophyllide reductase subunit L | C36 | −1.3 |
| C195 | −1.9 |
| Accession | UniProt ID | Gene | Protein | Cys Site(s) | log2FC |
|---|---|---|---|---|---|
| Cre01.g038600.t1.2 | A1E5M5 | FAD7 | Chloroplastic glycerolipid omega-3-fatty acid desaturase | C75 | −1.6 |
| C230 | −1.6 | ||||
| Cre01.g055408.t1.1 | A8JCQ8 | ACS2 | Acetyl-CoA synthetase | C164 | −1.0 |
| C594 | −1.1 | ||||
| Cre02.g105200.t1.2 | A8I478 | Unknown | Saposin B domain-containing protein | C103, C109 | 1.6 |
| Cre03.g182750.t1.2 | A0A2K3DXV3 | Unknown | Phospholipase B-like protein | C653 | 3.3 |
| Cre03.g210513.t1.1 | A8JBW0 | Unknown | 12-oxophytodienoate reductase 2 | C23 | −4.0 |
| Cre05.g235700.t1.2 | A8J8E3 | Unknown | Saposin related protein | C213, C224 | 2.9 |
| Cre13.g585301.t1.1 | H1AFJ8 | CPLD55 | Monogalactosyldiacylglycerol synthase | C272 | −1.0 |
| Cre13.g590500.t1.1 | O48663 | FAD6 | Omega-6-fatty acid desaturase | C387 | −1.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smythers, A.L.; McConnell, E.W.; Lewis, H.C.; Mubarek, S.N.; Hicks, L.M. Photosynthetic Metabolism and Nitrogen Reshuffling Are Regulated by Reversible Cysteine Thiol Oxidation Following Nitrogen Deprivation in Chlamydomonas. Plants 2020, 9, 784. https://doi.org/10.3390/plants9060784
Smythers AL, McConnell EW, Lewis HC, Mubarek SN, Hicks LM. Photosynthetic Metabolism and Nitrogen Reshuffling Are Regulated by Reversible Cysteine Thiol Oxidation Following Nitrogen Deprivation in Chlamydomonas. Plants. 2020; 9(6):784. https://doi.org/10.3390/plants9060784
Chicago/Turabian StyleSmythers, Amanda L., Evan W. McConnell, Hailey C. Lewis, Saher N. Mubarek, and Leslie M. Hicks. 2020. "Photosynthetic Metabolism and Nitrogen Reshuffling Are Regulated by Reversible Cysteine Thiol Oxidation Following Nitrogen Deprivation in Chlamydomonas" Plants 9, no. 6: 784. https://doi.org/10.3390/plants9060784