The Recovery of Soybean Plants after Short-Term Cadmium Stress

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
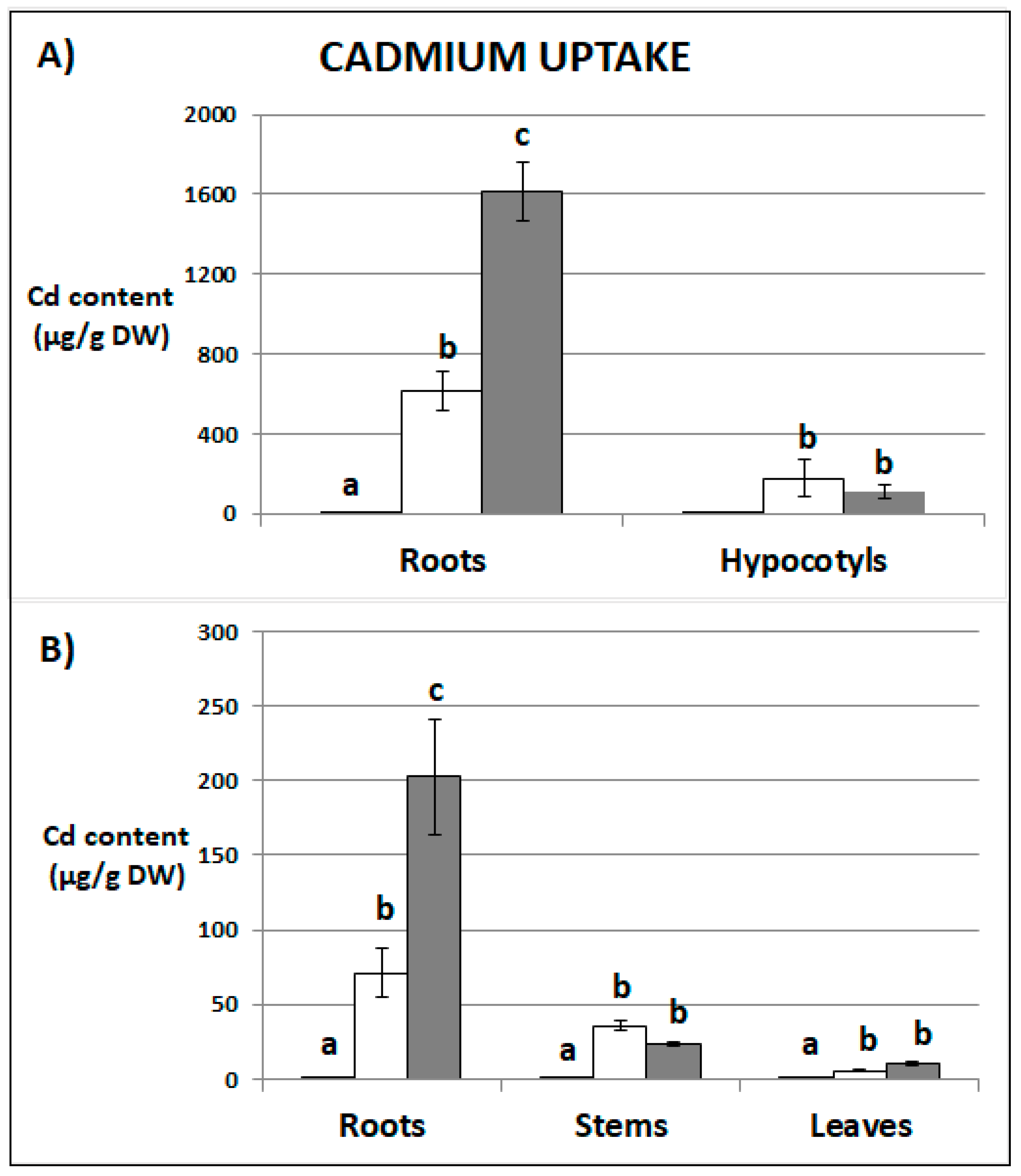
2.1. Cadmium Content
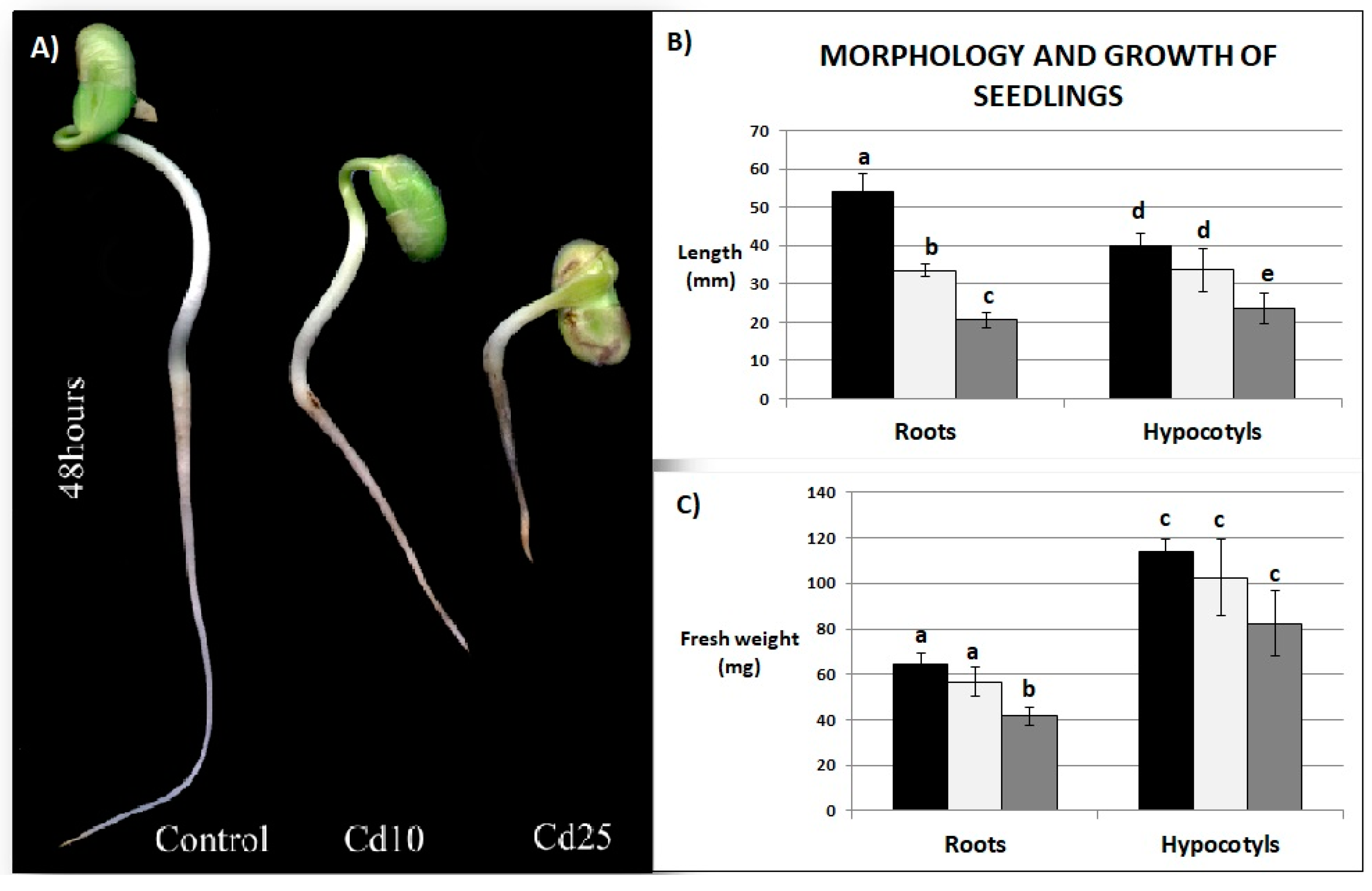
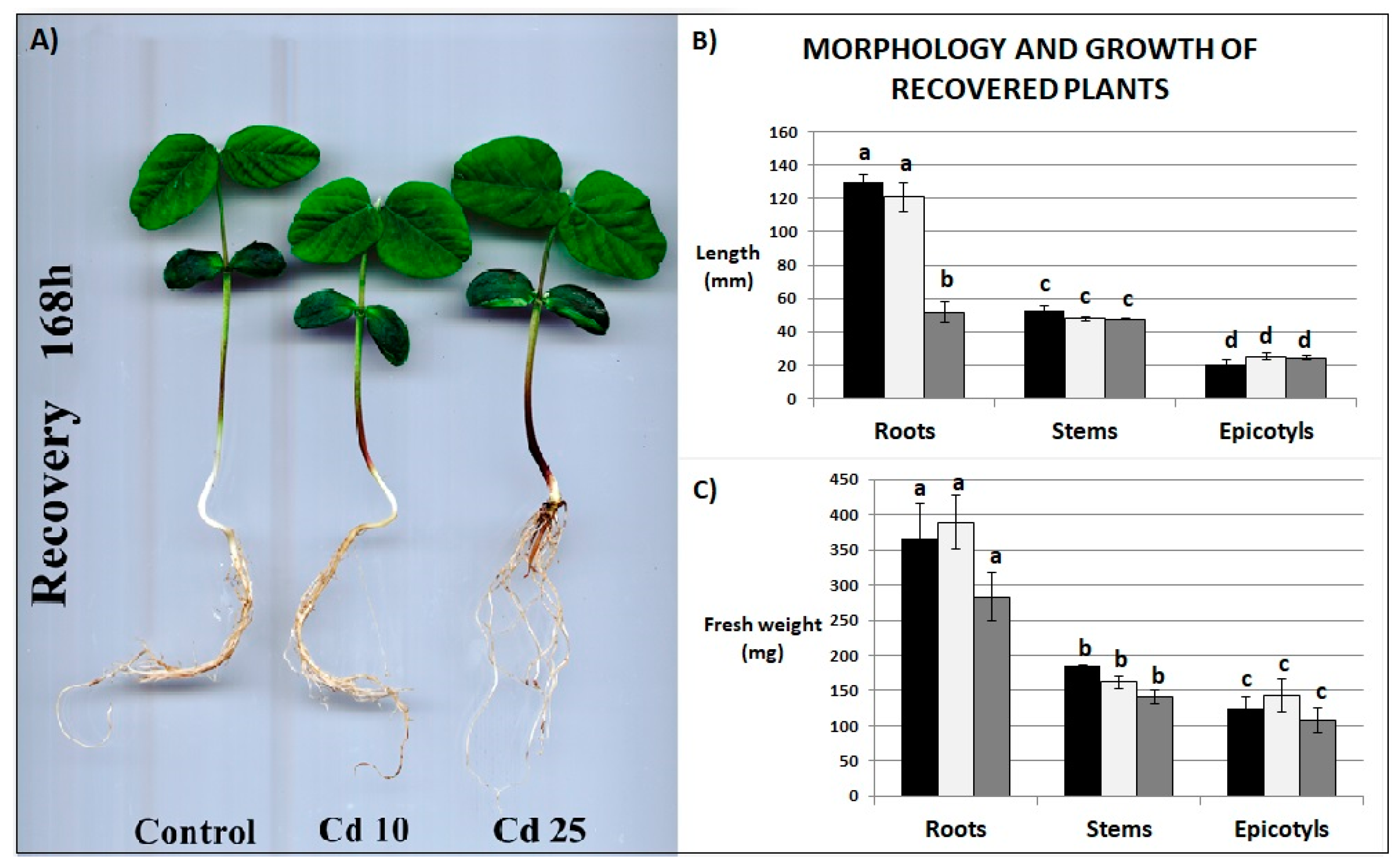
2.2. Growth Parameters
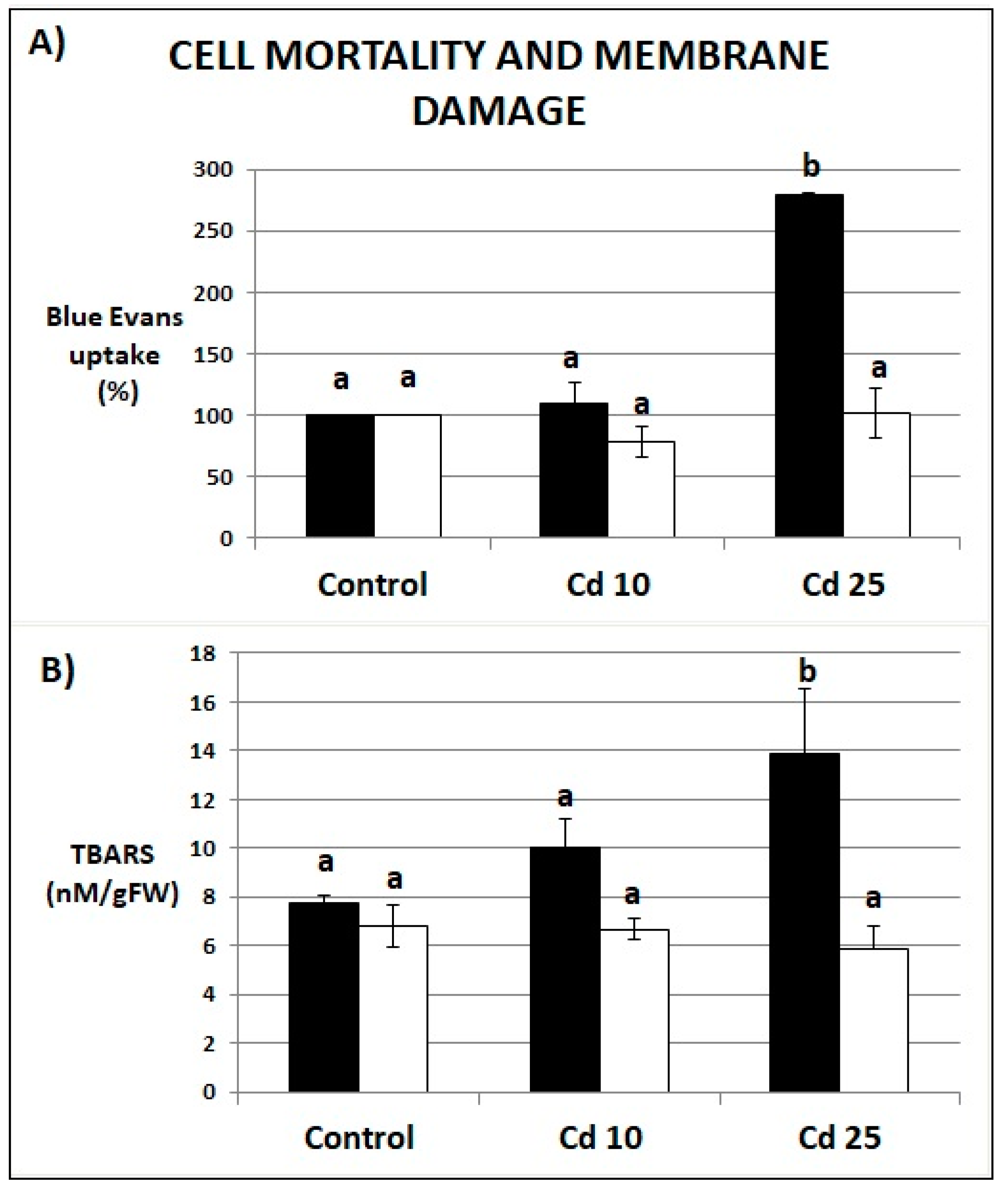
2.3. Cell Death and Membrane Integrity
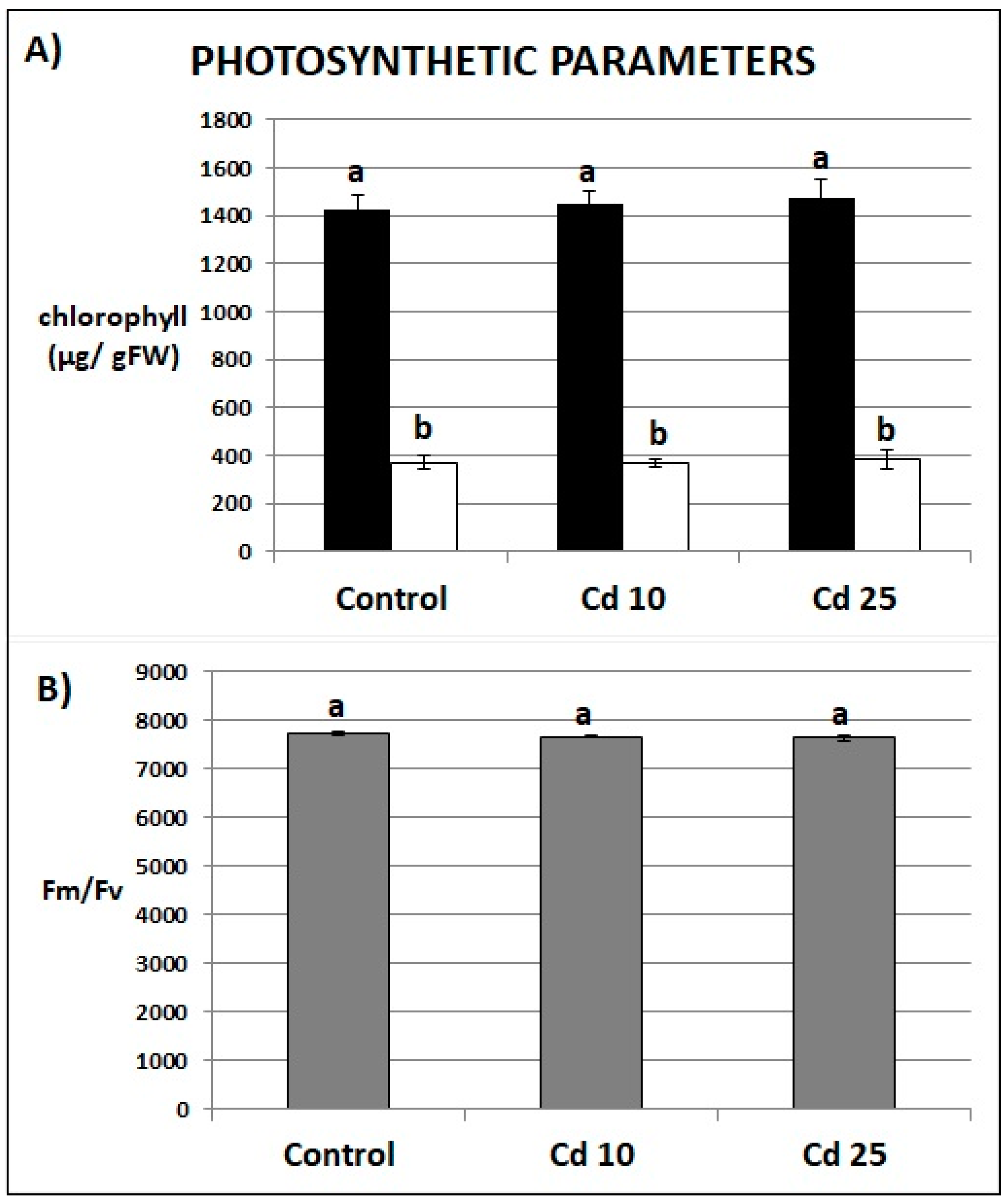
2.4. Photosynthesis Parameters of Recovered Plants
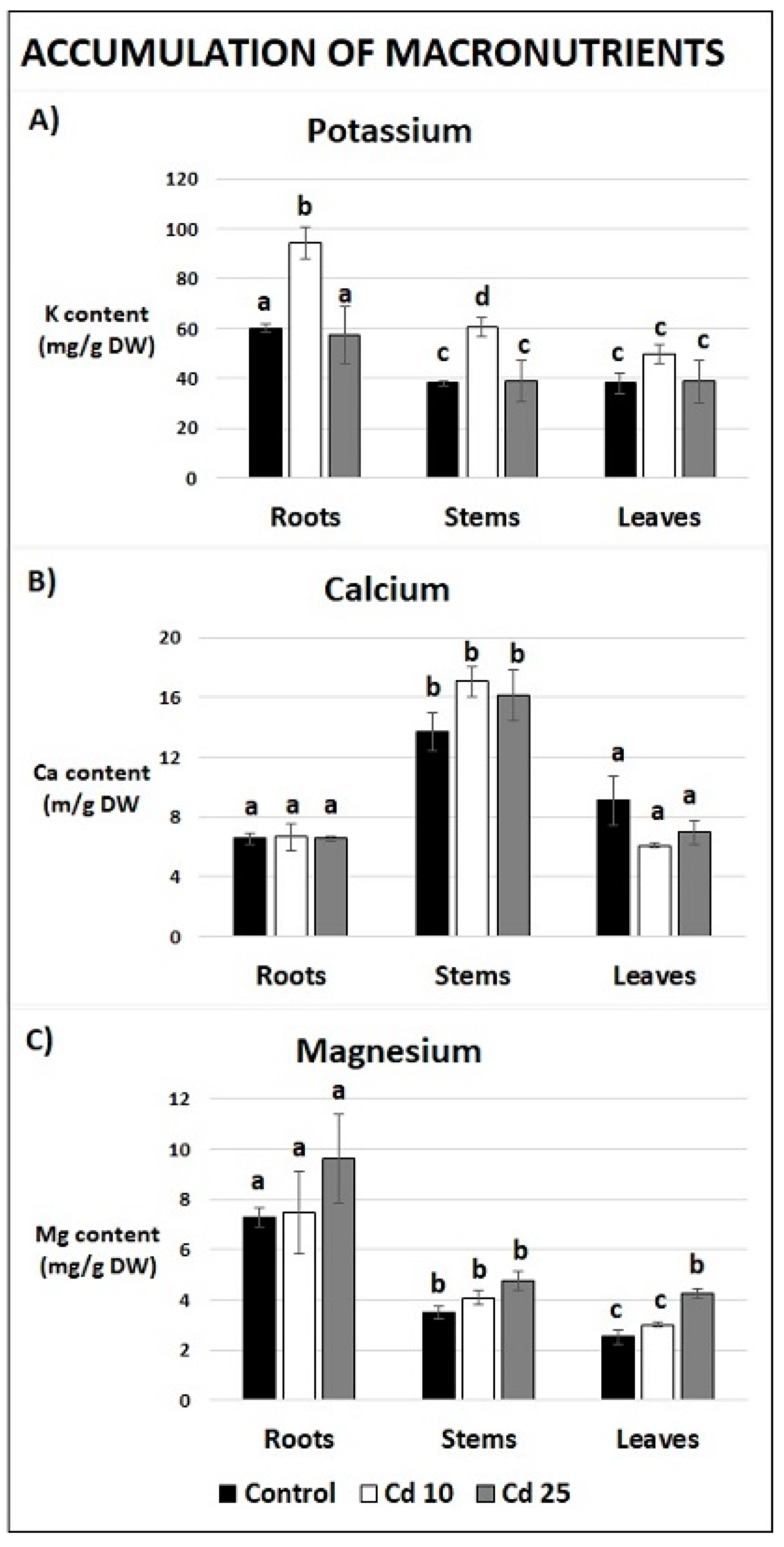
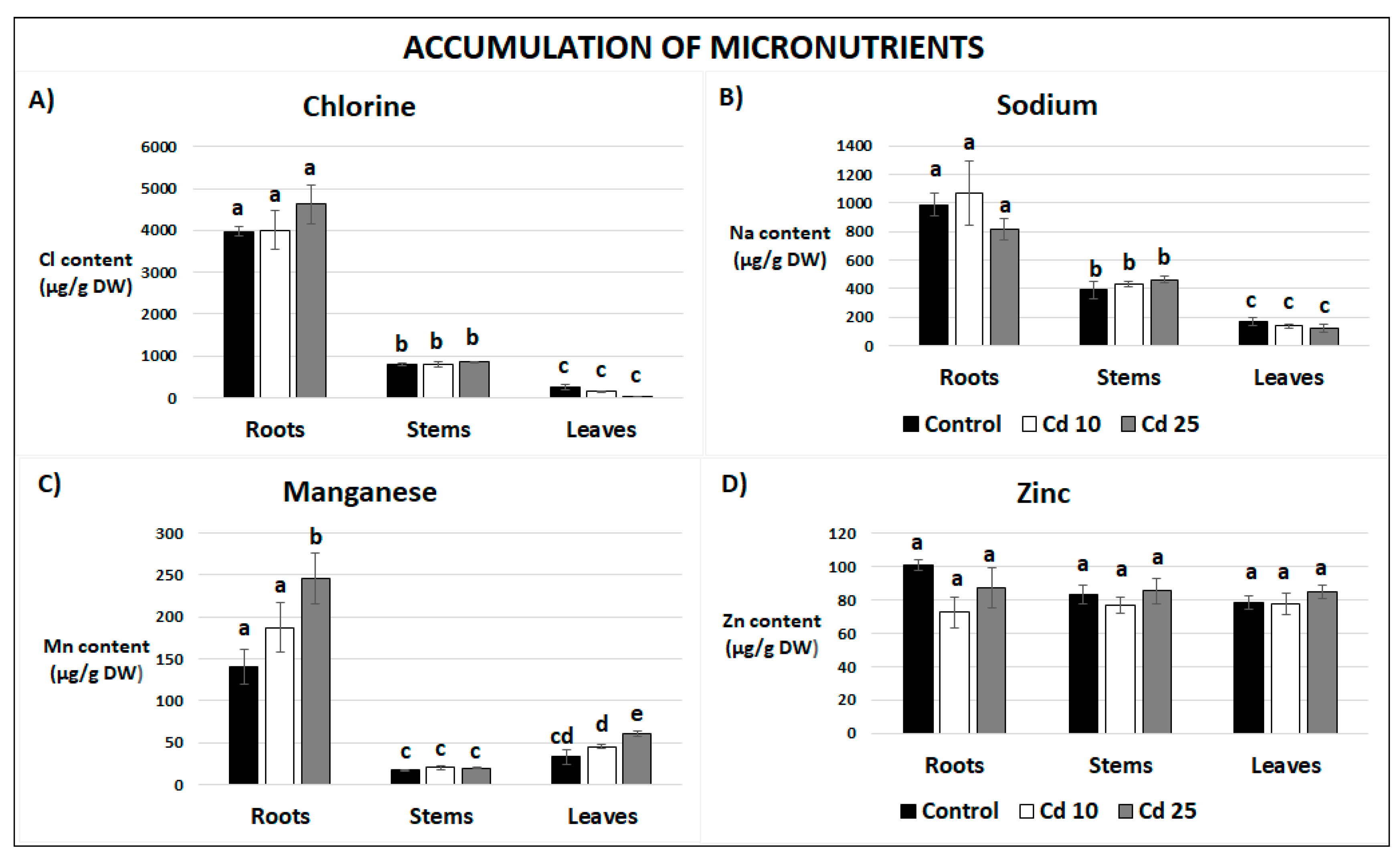
2.5. Content of Essential and Non-Essential Elements in Recovered Plants
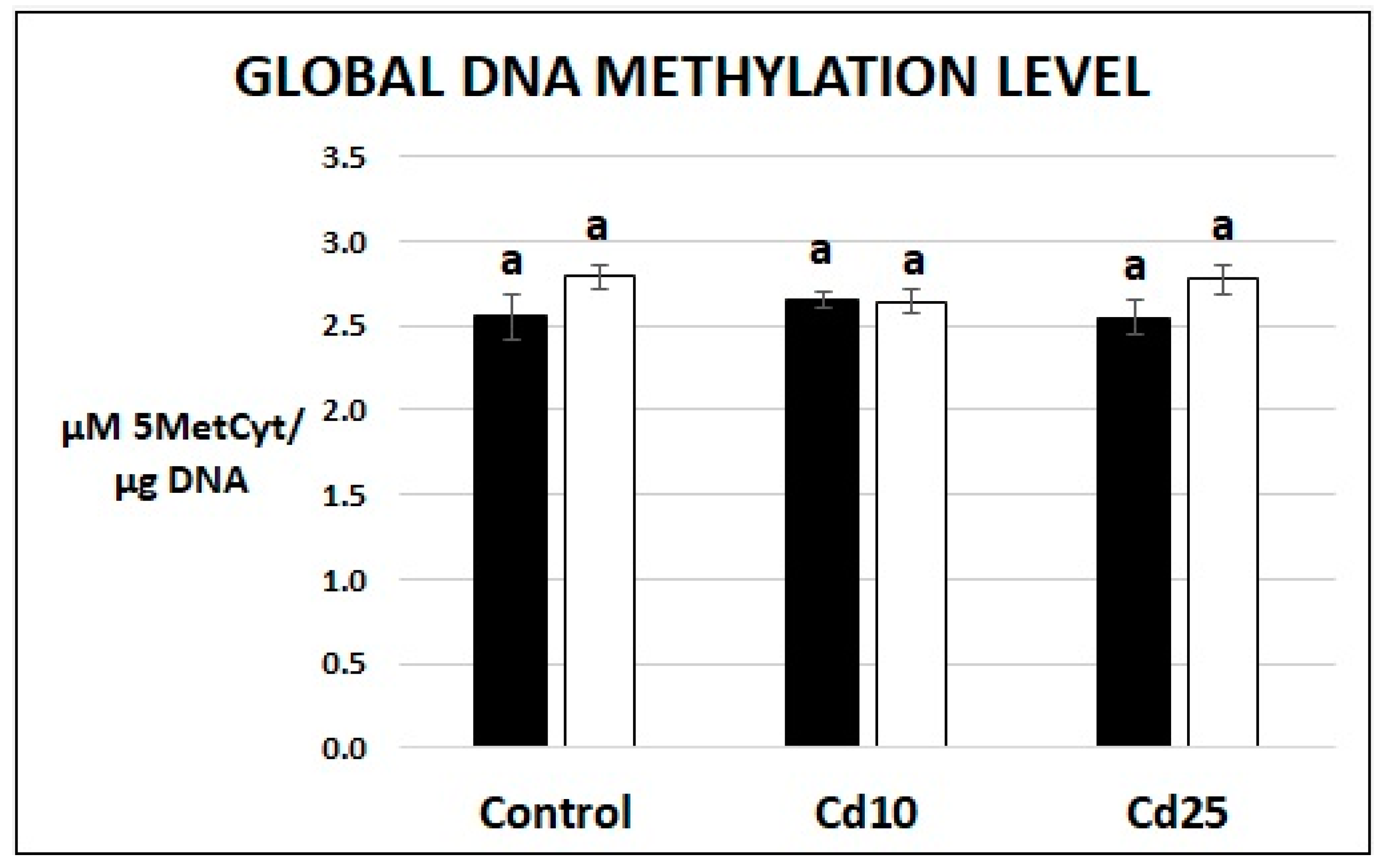
2.6. Examination of Global DNA Methylation Level
3. Discussion
4. Materials and Methods
4.1. Plant Cultivation and Treatment
4.2. Measurement of Cd Content
4.3. Measurements of Growth Parameters
4.4. Estimation of Cell Viability
4.5. Evaluation of Membrane Integrity
4.6. Measurements of Chlorophyll Content and Photosynthesis Parameters
4.7. Assessment of the Elements Content
4.8. Analysis of the Level of DNA Methylation
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arasimowicz-Jelonek, M.; Floryszak-Wieczorek, J.; Deckert, J.; Rucińska-Sobkowiak, R.; Gzyl, J.; Pawlak-Sprada, S.; Abramowski, D.; Jelonek, T.; Gwóźdź, E.A. Nitric oxide implication in cadmium-induced programmed cell death in roots and signaling response of yellow lupine plants. J. Plant Physiol. Biochem. 2012, 58, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-C.; Chao, Y.-Y.; Hong, C.-Y.; Kao, C.H. The role of hydrogen peroxide in cadmium-inhibited root growth of rice seedlings. Plant Growth Regul. 2012, 66, 27–35. [Google Scholar] [CrossRef]
- Garnier, L.; Simon-Plas, F.; Thuleau, P.; Agnel, J.-P.; Blein, J.-P.; Ranjeva, R.; Montillet, J.-L. Cadmium affects tobacco cells by a series of three waves of reactive oxygen species that contribute to cytotoxicity. Plant Cell Environ. 2006, 29, 1956–1969. [Google Scholar] [CrossRef] [PubMed]
- Gzyl, J.; Rymer, K.; Gwóźdź, E.A. Differential response of antioxidant enzymes to cadmium stress in tolerant and sensitive cell line of cucumber (Cucumis sativus L.). Acta Biochim. Pol. 2009, 56, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Alharby, H.F.; Razafindrabe, B.H.N.; Masayuki, F. Hydrogen peroxide pretreatment mitigates cadmium-induced oxidative stress in Brassica napus L.: An instrict study on antioxidant defense and glyoxalase systems. Front. Plant Sci. 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Gajewska, E.; Skłodowska, M. Differential effect of equal copper, cadmium and nickel concentrations on biochemical reactions in wheat seedlings. Ecotoxicol. Environ. Saf. 2010, 73, 996–1003. [Google Scholar] [CrossRef]
- Guo, Q.; Meng, L.; Zhang, Y.-N.; Mao, P.-C.; Tian, X.-X.; Li, S.-S.; Zhang, L. Antioxidative systems, metal ion homeostasis and cadmium distribution in Iris lactea exposed to cadmium stress. Ecotoxicol. Environ. Saf. 2017, 139, 50–55. [Google Scholar] [CrossRef]
- Souguir, D.; Ferjani, E.E.; Ledoigt, G.; Goupil, P. Transcript accumulation of stress-related genes in Vicia faba roots under a short term exposure to cadmium. Plant Biosyst. 2015, 149, 280–288. [Google Scholar] [CrossRef]
- Pena, L.B.; Pasquini, L.A.; Tomaro, M.L.; Gallego, S.M. 20S Proteosome and accumulation of oxidized and ubiquitinated proteins in maize leaves subjected to cadmium stress. Phytochemistry 2007, 68, 1139–1146. [Google Scholar] [CrossRef]
- Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Méndez, A.A.E.; Gallego, S.M. Oxidative post translation modifications of proteins related to cell cycle are involved in cadmium toxicity in wheat seedlings. Plant Sci. 2012, 196, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chmielowska-Bąk, J.; Izbiańska, K.; Deckert, J. The toxic Doppelganger: On the ionic and molecular mimicry of cadmium. Acta Biochim. Pol. 2013, 60, 369–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andresen, E.; Kappel, S.; Stärk, H.-J.; Riegger, U.; Borovec, J.; Mattusch, J.; Heinz, A.; Schmelzer, C.E.H.; Matoušková, Š.; Dickinson, B.; et al. Cadmium toxicity investigated at the physiological and biophysical levels under environmentally relevant conditions using the aquatic model plant Ceratophyllum demersum. New Phytol. 2016, 210, 1244–1258. [Google Scholar] [CrossRef] [PubMed]
- Rivetta, A.; Negrini NCocucci, M. Involvement of Ca2+-calmodulin in Cd2+ toxicity during the early phases of radish (Raphanus sativus L.) seed germination. Plant Cell Environ. 1997, 20, 600–608. [Google Scholar] [CrossRef]
- Malgieri, G.; Zaccaro, L.; Leone, M.; Bucci, E.; Esposito, S.; Baglivo, I.; Del Gatto, A.; Russo, L.; Scandurra, R.; Pedone, P.V.; et al. Zinc to cadmium replacement in the A. thaliana SUPERAN Cys2His2 Zinc finger induces structural rearrangements of typical DNA base determinant positions. Biopolymers 2011, 95, 801–810. [Google Scholar] [PubMed]
- Li, S.; Yu, J.; Zhu, M.; Zhao, F.; Luan, S. Cadmium impairs ion homeostasis by altering K+ and Ca2+ channel activities in rice root hair cells. Plant Cell Environ. 2012, 35, 1998–2013. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, M.; Liu, Z.; Shi, Y.; Han, T.; Ye, Y.; Gong, N.; Sun, J.; Zhu, C. Different responses of low grain-Cd-accumulating and high grain-Cd-accumulating rice cultivars to Cd stress. Plant Physiol. Biochem. 2015, 96, 261–269. [Google Scholar] [CrossRef]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zao, X.; Zao, X.; Hu, C. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizers. Metallomics 2019, 11, 225. [Google Scholar] [CrossRef]
- Okereafor, U.; Makhatha, M.; Mekuto, L.; Uche-Okereafor, N.; Sebola, T.; Mavumengwana, V. Toxic metal implications on agricultural soils, plants, animals, aquatic life and human health. Int. J. Environ. Res. Public Health 2012, 12, 2204. [Google Scholar] [CrossRef] [Green Version]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Chmielowska-Bąk, J.; Gzyl, J.; Rucińska-Sobkowiak, R.; Arasimowicz-Jelonek, M.; Deckert, J. The new insights into cadmium sensing. Front. Plant Sci. 2014, 5, 24. [Google Scholar]
- Drost, W.; Matzke, M.; Backhaus, T. Heavy metal toxicity in Lemna minor: Studies on the time dependence of growth inhibition and recovery after exposure. Chemosphere 2007, 67, 36–43. [Google Scholar] [CrossRef]
- Fojtová, M.; Fulnečková, J.; Fajkus, J.; Kovařík, A. Recovery of tobacco cells from cadmium stress is accompanied by DNA repair and increased telomerase activity. J. Exp. Bot. 2002, 53, 2151–2158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, M.; Paszkowski, J. Epigenetic memory in plants. EMBO J. 2014, 33, 1987–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, T.; Seki, M. Epigenetic memory for stress response and adaptation in plants. Plant Cell Physiol. 2014, 55, 1859–1863. [Google Scholar] [CrossRef] [PubMed]
- Rehman, Z.U.; Khan, S.; Brusseau, M.L.; Shah, M.T. Lead and cadmium contamination and exposure risk assessment via consumption of vegetables grown in agricultural soils of five-selected regions of Pakistan. Chemosphere 2017, 168, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Rzętała, M.A. Cadmium contamination of sediments in the water reservoirs in Silesian Upland (southern Poland). J. Soils Sediments 2016, 16, 2458–2470. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Yang, L. A review of heavy metal contaminations in urban soils, urban road dusts and agricultural soils from China. Microchem. J. 2010, 94, 99–107. [Google Scholar] [CrossRef]
- Bazihizina, N.; Taiti, C.; Serre, N.; Nocci, C.; Spinelli, F.; Nissim, W.G.; Azzarello, E.; Marti, L.; Redwan, M.; Gonelli, C.; et al. Awaiting better times: A quiescence response and adventitious root primordia formation prolong survival under cadmium stress in Tetradenia riparia (Hochs) Codd. Envrion. Exp. Bot. 2016, 130, 1–10. [Google Scholar] [CrossRef]
- Mohanty, S.; Das, A.B.; Das, P.; Mohanty, P. Effect of a low dose of aluminum on mitotic and meiotic activity, 4C DNA content, and pollen sterility in rice, Oryza sativa L. cv. Lalat. Ecotoxicol. Environ. Saf. 2004, 59, 70–75. [Google Scholar] [CrossRef]
- Abbad, A.M.; Hammad SSoliman, W.S. Influence of copper and lead on germination of three Mimosoideae plant species. Asian J. Agric. Biol. 2017, 5, 320–327. [Google Scholar]
- Tripathi, B.N.; Mehta, S.K.; Gaur, J.P. Recovery of uptake and assimilation of nitrate in Scenedesmus sp. previously exposed to elevated levels of Cu2+ and Zn2+. J. Plant Physiol. 2004, 161, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska ELabudda, M. Dual role of metallic trace elements in stress biology—From negaitve to beneficial impact on plants. Int. J. Mol. Sci. 2019, 20, 3117. [Google Scholar] [CrossRef] [Green Version]
- Chmielowska, J.; Veloso, J.; Gutiérrez, J.; Silvar, C.; Díaz, J. Cross-protection of pepper plants stressed by copper against a vascular pathogen is accompanied by the induction of a defence response. Plant Sci. 2010, 178, 176–182. [Google Scholar] [CrossRef]
- Arasimowicz-Jelonek, M.; Floryszak-Weiczorek, J.; Drzewiecka, K.; Chemilowska-Bąk, J.; Abramowski, D.; Iżbiańska, K. Aluminum induced cross-resistance of potato to Phytophtora infestans. Planta 2013, 239, 679–694. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Li, M.; Zuo, J.; Jiang, W.; Liu, D. Cadmium effects on mineral accumulation, antioxidant defence system and gas exchange in cucumber. Zemdirbyste-Agricul 2015, 102, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.Y.; Shi, G.X.; Xu, Q.S.; Wang, H.X. Cadmium effects on mineral nutrition and stress-related indices in Potamogeton crispus. Russ. J. Plant Physiol. 2011, 58, 253–260. [Google Scholar] [CrossRef]
- Gomes, M.P.; Marques, T.; Soares, A.M. Cadmium effects on mineral nutrition of the Cd-hyperaccumulator Pfaffia glomerata. Biologia 2013, 68, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, M.E.A.; Piotto, F.A.; Franco, M.R.; Borges, K.L.R.; Gaziola, S.A.; Castro, P.R.C.; Azevedo, R.A. Cadmium toxicity degree on tomato development is associated with disbalances in B and Mn status at early stages of plant exposure. Ecotoxicology 2018, 27, 1293–1302. [Google Scholar] [CrossRef]
- Sun, H.; Chen, Z.-H.; Chen, F.; Xie, L.; Zhang, G.; Vincze, E.; We, F. DNA microarray revealed and RNAi plant confirmed key genes conferring low Cd accumulation in barley plants. BMC Plant Biol. 2015, 15, 259. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Liu, X.; Cui, B.; Bi, J.; Wang, X. Concentration-dependnet alterations in gene expression induced by cadmium in Solanum lycopersicum. Environ. Sci. Pollut. Res. 2017, 24, 10528–10536. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Liu, X.; Wan, J.; Zhao, S.; Cui, B. Microarray-based analysis of gene expression in Lycopersicon esculentum seedling roots in response to cadmium, chromium, mercury and lead. Envrion. Sci. Technol. 2015, 49, 1834–1841. [Google Scholar]
- Rekik, I.; Chaâbene, Z.; Kriaa, W.; Rorat, A.; Franck, V.; Hafedh, M.; Elleuch, A. Transcriptome assembly and abiotic related gene expression analysis of date palm reveal candidate genes involved in response to cadmium stress. Comp. Bioch. Physiol. 2019, 225, 108569. [Google Scholar] [CrossRef]
- Xie, M.; Chen, W.; Lai, X.; Dai, H.; Sun, H.; Zhou, X.; Chen, T. Metabolic responses and their correlations with phytochelatins in Amaranthus hypochondriacus under cadmium stress. Environ. Poll. 2019, 252, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.J.; Liu, X.S.; Tao, H.; Tan, S.K.; Chu, S.S.; Oono, Y.; Zhang, X.D.; Chen, J.; Yang, Z.M. Variation of DNA methylation patterns associated with gene expression in rice (Oryza sativa) exposed to cadmium. Plant Cell Environ. 2016, 39, 2629–2649. [Google Scholar] [CrossRef]
- Shafiq, S.; Zeb, Q.; Ali, A.; Sajjad, Y.; Nazir, R.; Widemann, E.; Liu, L. Lead, cadmium and zinc phytotoxicity alter DNA methylation levels to confer heavy metal tolerance in wheat. Int. J. Mol. Sci. 2019, 20, 4676. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.S.; Feng, S.J.; Zhang, B.Q.; Wang, M.Q.; Cao, H.W.; Rono, J.K.; Chen, X.; Yang, Z.M. OsZIP1 functions as metal efflux transporter limiting excess zinc, copper and cadmium accumulation in rice. BMC Plant Biol. 2019, 19, 283. [Google Scholar] [CrossRef]
- Imran, M.; Shafiq, S.; Naeem, M.K.; Widemann, E.; Munir, M.Z.; Jensen, K.B.; Wang, R.R.-C. Histone deacetylase (HADC) gene family in allotetraploid cotton and its diploid progenitors: In silico identification, molecular characterization, and gene expression analysis under multiple abiotic stresses, DNA damage and phytohormone treatments. Int. J. Mol. Sci. 2020, 21, 321. [Google Scholar] [CrossRef] [Green Version]
- Sobkowiak, R.; Deckert, J. Proteins induces by cadmium in soybean cells. J. Plant Physiol. 2006, 163, 1203–1206. [Google Scholar] [CrossRef]
- Striker, G.G. Time is on our side: The importance of considering a recovery periods when assessing flooding tolerance in plants. Ecol. Res. 2011, 27, 983–987. [Google Scholar] [CrossRef]
- Chmielowska-Bąk, J.; Lefèvre, I.; Lutts, S.; Deckert, J. Short term signaling responses in roots of young soybean seedlings exposed to cadmium stress. J. Plant Physiol. 2013, 170, 1585–1594. [Google Scholar] [CrossRef] [PubMed]
- Chmielowska-Bąk, J.; Lefèvre, I.; Lutts, S.; Kulik, A.; Deckert, J. Effect of an ethylene synthesis inhibitor, cobalt chloride, on soybean seedlings subjected to cadmium stress. Acta Societ. Bot. Pol. 2014, 83, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, S.; Firych, A.; Rymer, K.; Deckert, J. Cu,Zn-superoxide dismutase is differently regulated by cadmium and lead in roots of soybean seedlings. Acta Physiogiae Plantarum 2009, 31, 741–747. [Google Scholar] [CrossRef]
- Lehotai, N.; Petö, A.; Bajkán, S.; Erdei, L.; Tari, I.; Kolbert, Z. In vivo and in situ visualization of early physiological events induces by heavy metals in pea root meristem. Acta Physiologiae Plantarum 2011, 33, 2199–2207. [Google Scholar] [CrossRef]
- Cuypers, A.; Smeets, K.; Ruytinx, J.; Opdenakker, K.; Keunen, E.; Remens, T.; Horemans, N.; Vanhoudt, N.; Van Sanden, S.; Van Belleghem, F.; et al. The cellular redox state as a modulator in cadmium and copper responses in Arabidopsis thaliana seedlings. J. Plant Physiol. 2011, 168, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Pavlov, S.S.; Dmitriev, A.Y.; Frontasyeva, M.V. Automation system for neutron activation analysis at the reactor IBR-2, Frank Laboratory of Neutron Physics, Joint Institute for Nuclear Research, Dubna, Russia. J. Radioanal Nucl Chem. 2016, 309, 27–38. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holubek, R.; Deckert, J.; Zinicovscaia, I.; Yushin, N.; Vergel, K.; Frontasyeva, M.; Sirotkin, A.V.; Bajia, D.S.; Chmielowska-Bąk, J. The Recovery of Soybean Plants after Short-Term Cadmium Stress. Plants 2020, 9, 782. https://doi.org/10.3390/plants9060782
Holubek R, Deckert J, Zinicovscaia I, Yushin N, Vergel K, Frontasyeva M, Sirotkin AV, Bajia DS, Chmielowska-Bąk J. The Recovery of Soybean Plants after Short-Term Cadmium Stress. Plants. 2020; 9(6):782. https://doi.org/10.3390/plants9060782
Chicago/Turabian StyleHolubek, Renata, Joanna Deckert, Inga Zinicovscaia, Nikita Yushin, Konstantin Vergel, Marina Frontasyeva, Alexander V. Sirotkin, Donald Samdumu Bajia, and Jagna Chmielowska-Bąk. 2020. "The Recovery of Soybean Plants after Short-Term Cadmium Stress" Plants 9, no. 6: 782. https://doi.org/10.3390/plants9060782