Easy-to-Use InDel Markers for Genetic Mapping between Col-0 and Ler-0 Accessions of Arabidopsis thaliana


Abstract
:1. Introduction
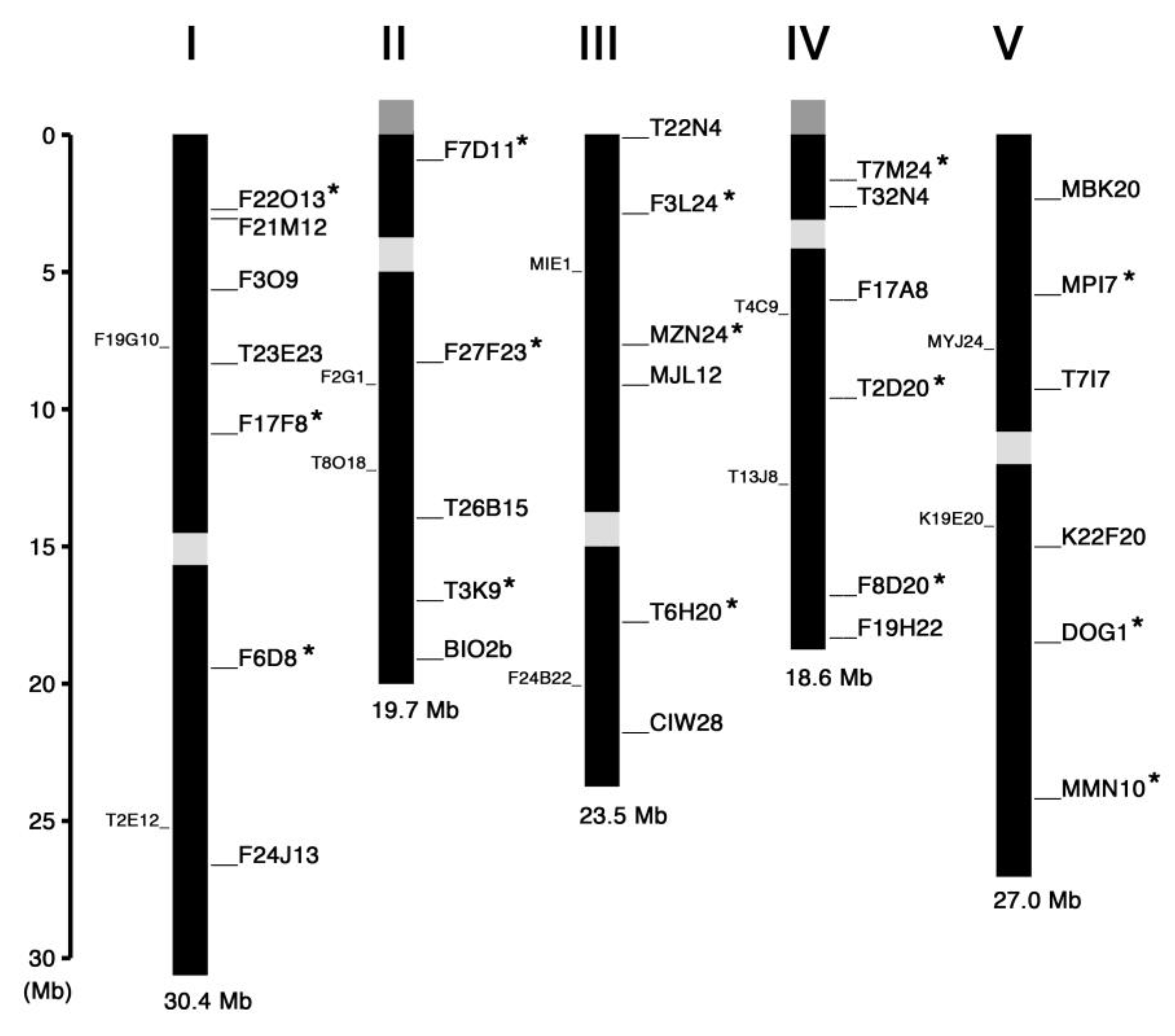
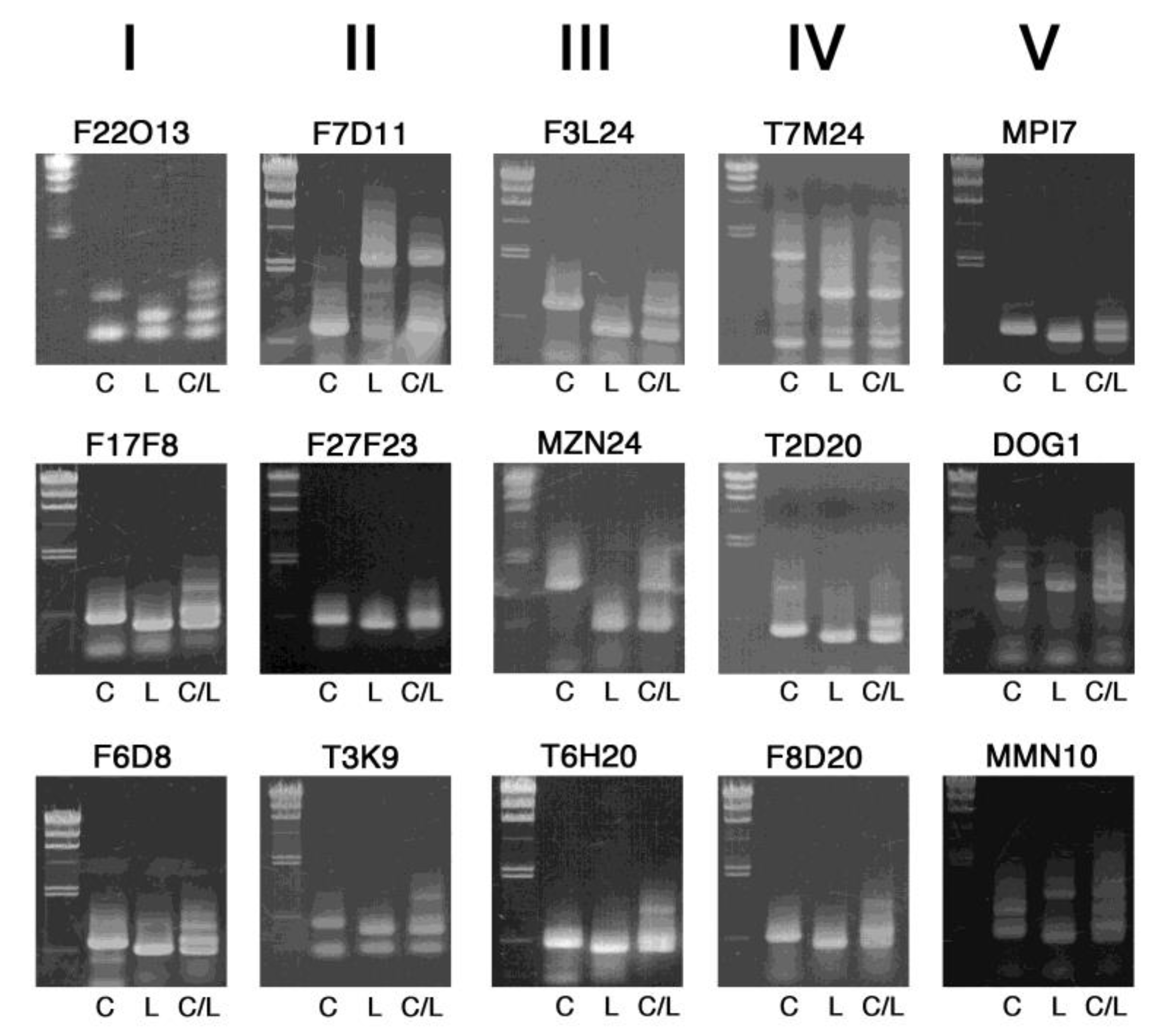
2. Results and Discussion
3. Materials and Methods
3.1. DNA Extraction
3.2. PCR and Agarose Gel Electrophoresis
Author Contributions
Funding
Conflicts of Interest
References
- Jander, G.; Norris, S.R.; Rounsley, S.D.; Bush, D.F.; Levin, I.M.; Last, R.L. Arabidopsis map-based cloning in the post-genome era. Plant Physiol. 2002, 129, 440–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, J.L.; Cnudde, F.; Gerats, T. Forward genetics and map-based cloning approaches. Trends Plant Sci. 2003, 8, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Mao, Y.; Xu, N.; Zhang, B.; Wei, P.; Yang, D.-L.; Wang, Z.; Zhang, Z.; Zheng, R.; Yang, L. Multigeneration analysis reveals the inheritance, specificity, and patterns of CRISPR/Cas-induced gene modifications in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 4632–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauser, F.; Schiml, S.; Puchta, H. Both CRISPR/Cas-based nucleases and nickases can be used efficiently for genome engineering in Arabidopsis thaliana. Plant J. 2014, 79, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Chen, J.; Dai, X.; Zhang, D.; Zhao, Y. An effective strategy for reliably isolating heritable and Cas9-free Arabidopsis mutants generated by CRISPR/Cas9-mediated genome editing. Plant Physiol. 2016, 171, 1794–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konieczny, A.; Ausubel, F.M. A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specific PCR-based markers. Plant J. 1993, 4, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J.; Drenkarci, E.; Preuss, D.; Ausubel, F.M. Use of cleaved amplified polymorphic sequences (CAPS) as genetic markers in Arabidopsis thaliana. Methods Mol. Biol. 1998, 82, 173–182. [Google Scholar] [PubMed]
- Bell, C.J.; Ecker, J.R. Assignment of 30 microsatellite loci to the linkage map of Arabidopsis. Genomics 1994, 19, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Neff, M.M.; Neff, J.D.; Chory, J.; Pepper, A.E. dCAPS, a simple technique for the genetic analysis of single nucleotide polymorphisms: Experimental applications in Arabidopsis thaliana genetics. Plant J. 1998, 14, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Li, L.; Peng, Z.; Wei, B.; Tang, S.; Ding, M.; Liu, J.; Zhang, F.; Zhao, Y.; Gu, H.; et al. A platform of high-density INDEL/CAPS markers for map-based cloning in Arabidopsis. Plant J. 2010, 63, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Pacurar, D.I.; Pacurar, M.L.; Street, N.; Bussell, J.D.; Pop, T.I.; Gutierrez, L.; Bellini, C. A collection of INDEL markers for map-based cloning in seven Arabidopsis accessions. J. Exp. Bot. 2012, 63, 2491–2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Glazebrook, J.; Li, X. Identification of components in disease-resistance signaling in Arabidopsis by map-based cloning. Methods Mol. Biol. 2007, 354, 69–78. [Google Scholar] [PubMed]
- Xing, J.; Weng, Q.; Si, H.; Han, J.; Dong, J. Identification and molecular tagging of two Arabidopsis resistance genes to Botrytis cinerea. Front. Agric. China 2011, 5, 430–436. [Google Scholar] [CrossRef]
- Bryant, F.; Hughes, D.; Hassani-Pak, K.; Eastmond, P. Basic LEUCINE ZIPPER TRANSCRIPTION FACTOR67 transactivates DELAY OF GERMINATION1 to establish primary seed dormancy in Arabidopsis. Plant Cell 2019, 31, 1276–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsuhashi, M. Technical report: Part 2. Basic requirements for designing optimal PCR primers. J. Clin. Lab. Anal. 1996, 10, 285–293. [Google Scholar] [CrossRef]
- Zapata, L.; Ding, J.; Willing, E.M.; Hartwig, B.; Bezdan, D.; Jiao, W.B.; Patel, V.; Velikkakam James, G.; Koornneef, M.; Ossowski, S.; et al. Chromosome-level assembly of Arabidopsis thaliana Ler reveals the extent of translocation and inversion polymorphisms. Proc. Natl. Acad. Sci. USA 2016, 113, 4052–4060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, B.; Sumit, R.; Srivastava, S.; Bhattacharyya, M. Sequence based polymorphic (SBP) marker technology for targeted genomic regions: Its application in generating a molecular map of the Arabidopsis thaliana genome. BMC Genom. 2012, 13, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Marker Name | Chr. | Position (Mbp) | Gene ID * | Length (bp) (Col-0/Ler) | Primer Sequence |
|---|---|---|---|---|---|
| F22O13 | 1 | 2.826 | At1g04667 | 460/210 | F: GTGTTGGGGAGAGCTTATAG |
| R: TCCACTTCCAACCATCAGAG | |||||
| F21M12 | 1 | 3.138 | At1g09700 (HYL1) | 580/500 | F: AAGACTCCATCTTGACACTG |
| R: CCTCAACCTACTGATCATTG | |||||
| F3O9 | 1 | 5.653 | At1g16530 (ASL9) | 610/530 | F: TTTTGGTCGGGTATGGAATG |
| R: CCAGAAGTTGCTCGTTAAAG | |||||
| T23E23 | 1 | 8.494 | At1g24000 | 540/420 | F: AAGGTCTTGTAGCGATCTAG |
| R: AACCCAACTGGCTCATTTTG | |||||
| F17F8 | 1 | 11.016 | At1g30930 | 460/360 | F: GGAAGAGGATTGACTCAAAG |
| R: CTACCGCTAGGACTTTCATG | |||||
| F6D8 | 1 | 19.621 | At1g52690 (LEA7) | 640/540 | F: GAGACACAGAGGAAGTGAAG |
| R: CTGACCAGCAAATTCTCAAG | |||||
| F24J13 | 1 | 26.624 | At1g70610 (ABCB26) | 600/500 | F: GCTACCCTTCAAGAGATGAG |
| R: TCGTAGAGTTGCAGCAAAAG | |||||
| F7D11 | 2 | 1.614 | At2g04622 | 670/580 | F: AGCGAACTTCGTTGATGTTC |
| R: CAATGTATATGCTCTTCTAGAG | |||||
| F27F23 | 2 | 8.410 | At2g19410 | 460/400 | F: TGACCAGTTGTACCAATGTG |
| R: GTCTGCGACAAAAAATACTG | |||||
| T26B15 | 2 | 13.825 | At2g32560 | 470/410 | F: AACACACTCTCTCTCTCTTG |
| R: AGGTCAAGAACCGACATTTG | |||||
| T3K9 | 2 | 17.107 | At2g40990 | 470/360 | F: TCATCGGAAGGAGCATTATG |
| R: AGGATGTTCCAGAGAGAATG | |||||
| BIO2b | 2 | 18.012 | At2g43360 (BIO2) | 390/460 | F: TGTACCTCCCTGAAGTTATG |
| R: TCTTGACCTCCTCTTCCATG | |||||
| T22N4 | 3 | 0.130 | At3g01345 | 2040/400 | F: TGACTGTTTGACTCCAAGTG |
| R: GTTACGAACCTCTGGTATTG | |||||
| F3L24 | 3 | 2.849 | At3g09270 (GSTU8) | 770/470 | F: TGAGCAATGATGGTTAGCAG |
| R: GAACGTAACTGCTTACGTAG | |||||
| MZN24 | 3 | 7.665 | At3g21750 (UGT71B1) | 1250/500 | F: ATCCGAACCGAAATCAACTG |
| R: GACTGAACGAGAGGAACATG | |||||
| MJL12 | 3 | 9.194 | At3g25240 | 650/490 | F: GGAGGCTAGAGACTCATATG |
| R: AGGGGATATTCGACTGAGAG | |||||
| T6H20 | 3 | 17.243 | At3g46820 (TOPP5) | 510/430 | F: AGTCCACCATGCATACAAAG |
| R: TGCATTGGTTTCTCTGCTTG | |||||
| CIW28 | 3 | 21.869 | At3g59140 (ABCC10) | 630/450 | F: GAGCACAAGTCTCTTACAAG |
| R: CCCTAAGTTTCACAAAGAATG | |||||
| T7M24 | 4 | 1.788 | At4g03826 | 1350/710 | F: TTTGGCGCTGTTGCCAATTG |
| R: TAATGCGCGAGGTGGATATG | |||||
| T32N4 | 4 | 2.549 | At4g04985 | 1180/900 | F: CTCAAGGTCGACATGATAATG |
| R: GTATAACGCGGGTCAATCTC | |||||
| F17A8 | 4 | 6.109 | At4g09670 | 1280/560 | F: TGCTCGAGAGACTTTTCGAG |
| R: CATAGACAGCCACACCAATG | |||||
| T2D20 | 4 | 9.652 | At4g17200 | 420/360 | F: TGGTCTTCTTATGCTCCAAG |
| R: AGAGGAAGCACACAGTATTG | |||||
| F8D20 | 4 | 16.924 | At4g35700 (DAZ3) | 540/460 | F: GGCGAGGATTGACTTAAATG |
| R: ACTGTTGCGATAATGCAGTG | |||||
| F19H22 | 4 | 18.133 | At4g38870 | 380/320 | F: GCGTTGTTGAGTGTAGCAAG |
| R: GAGATCGATCGTCATCTTTC | |||||
| MBK20 | 5 | 2.476 | At5g07770 (FH16) | 410/350 | F: AGAGACCCTTTTCTCTGTTG |
| R: GGAGCTTACCATCATATCAG | |||||
| MPI7 | 5 | 5.902 | At5g17860 (CAX7) | 570/490 | F: TCCAATTAGACCGCATATTAG |
| R: TTCGTTGCTTGAGACACTAG | |||||
| T7I7 | 5 | 9.271 | At5g26594 (RR24) | 650/500 | F: TGGCACCAAGAAGCAACTAG |
| R: TCCTAACTATCAACCAACTTG | |||||
| K22F20 | 5 | 15.027 | At5g00540 | 490/420 | F: ACCGCTACCATTTGTTCTTG |
| R: CCAACGTTCTTCCCTGTTAG | |||||
| DOG1 | 5 | 18.591 | At5g45830 (DOG1) | 1010/1300 | F: GCGTGTTTGTGTTTTGTGTG |
| R: ATCCGCTGTCTCAGGACATC | |||||
| MMN10 | 5 | 24.108 | At5g59840 (RABE1B) | 530/460 | F: TGAAGGATGACTCGTCTGTG |
| R: GATGGCTCTTTCACCACTAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, T.; Nishii, Y.; Matsuo, H.; Takahashi, T. Easy-to-Use InDel Markers for Genetic Mapping between Col-0 and Ler-0 Accessions of Arabidopsis thaliana. Plants 2020, 9, 779. https://doi.org/10.3390/plants9060779
Tanaka T, Nishii Y, Matsuo H, Takahashi T. Easy-to-Use InDel Markers for Genetic Mapping between Col-0 and Ler-0 Accessions of Arabidopsis thaliana. Plants. 2020; 9(6):779. https://doi.org/10.3390/plants9060779
Chicago/Turabian StyleTanaka, Takahiro, Yuichi Nishii, Hirotoshi Matsuo, and Taku Takahashi. 2020. "Easy-to-Use InDel Markers for Genetic Mapping between Col-0 and Ler-0 Accessions of Arabidopsis thaliana" Plants 9, no. 6: 779. https://doi.org/10.3390/plants9060779