Functional Traits, Morphology, and Herbage Production of Vernalised and Non-Vernalised Chicory cv. Choice (Cichorium intybus L.) in Response to Defoliation Frequency and Height

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Establishment of Chicory cv. Choice
2.2. Management of Chicory (Experiment 1)
2.3. Morphological and Physiological Measurements (Experiment 1)
2.4. Herbage Production Measurements (Experiment 1)
2.5. Plant Function of Vernalised Chicory Plants (Experiment 2)
2.6. Data Analysis
3. Results
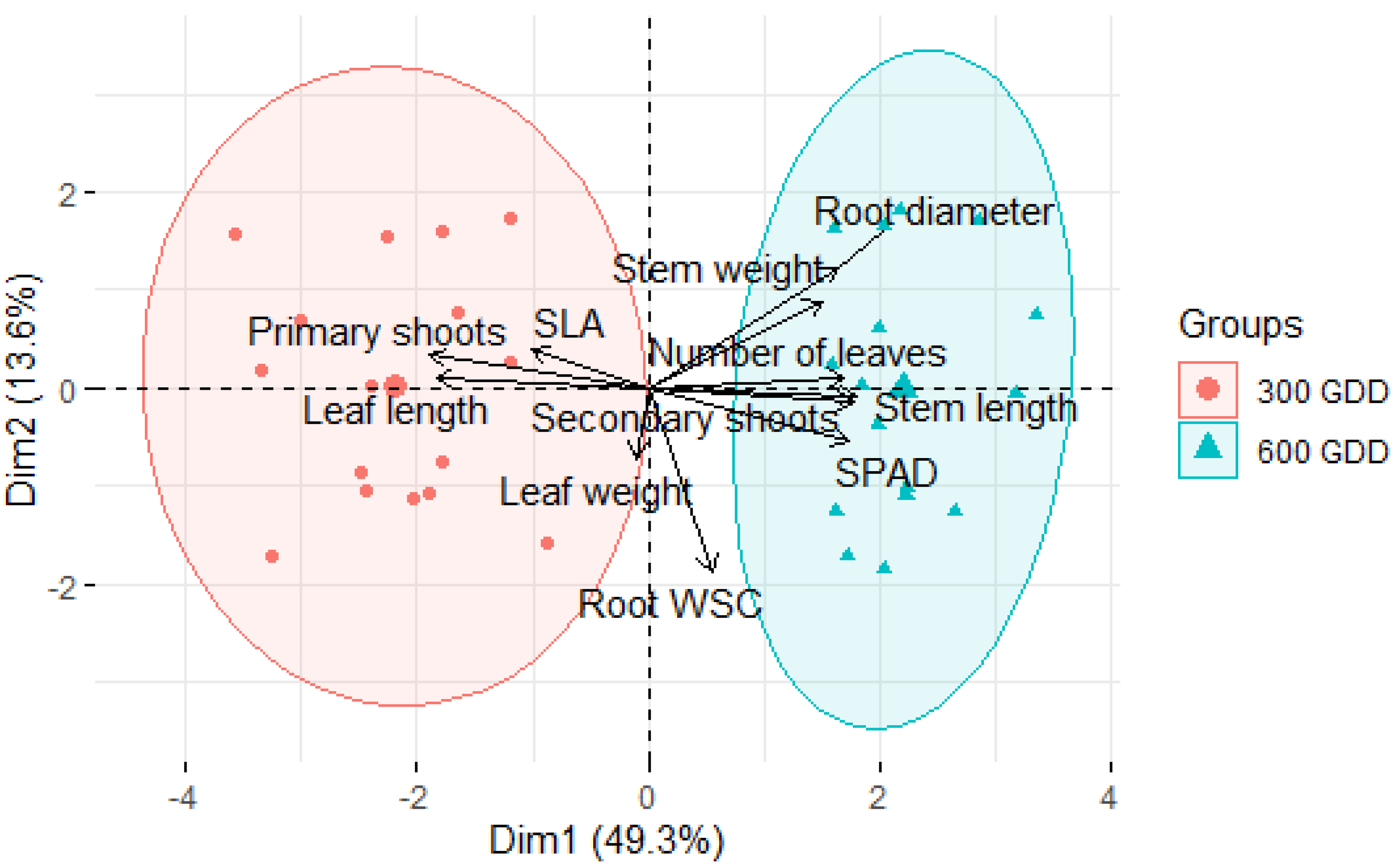
3.1. Morphology of Individual Chicory Plants
3.2. Herbage Production and Plant Density of Chicory Sward
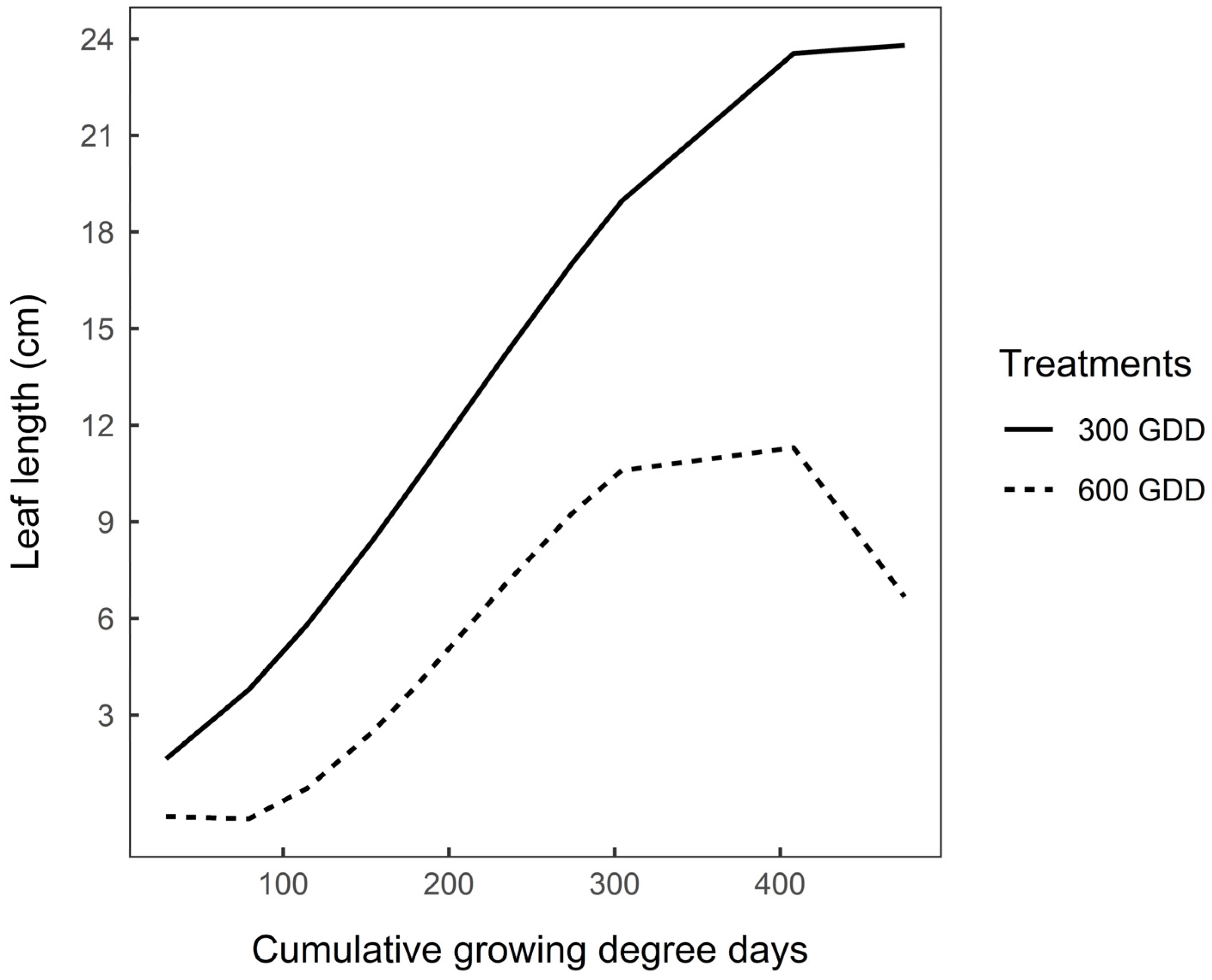
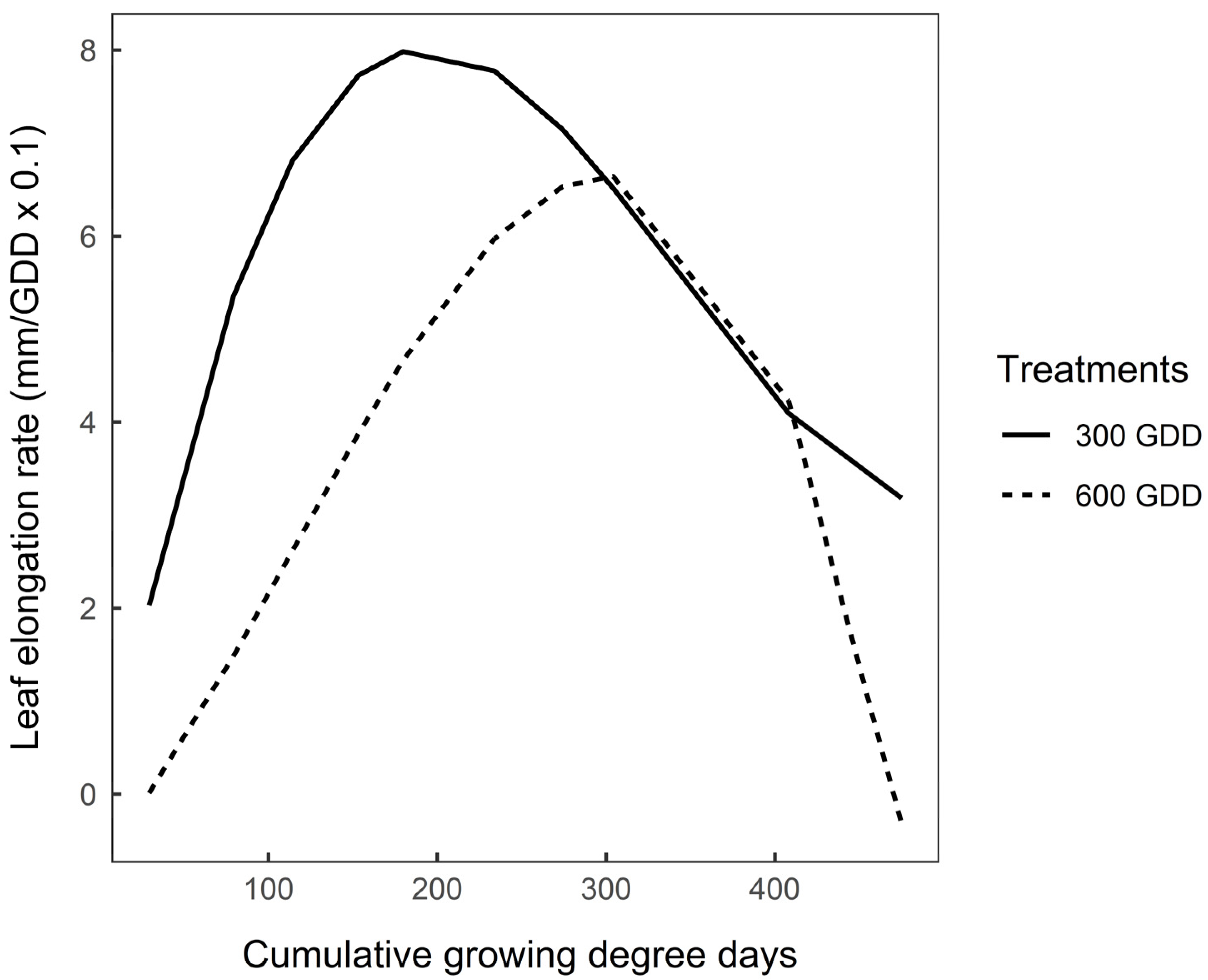
3.3. Functional Traits after Vernalisation
4. Discussion
4.1. Morphology and Functional Response of Chicory Plants
4.1.1. Above Ground Plant Morphology of Non-Vernalised Chicory
4.1.2. Above Ground Plant Morphology of Vernalised Chicory Plants
4.1.3. Root Size and Concentration of Stored Carbohydrates
4.2. Herbage Production and Plant Density of Chicory Swards
4.2.1. Before Vernalisation
4.2.2. After Vernalisation
4.3. Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Malcolm, B.J.; Carey, P.L.; Teixeira, E.I.; Johnstone, P.R.; Maley, S.C.; De Ruiter, J.M. Potential of catch crops to reduce nitrogen leaching in New Zealand winter grazing systems. J. N. Z. Grasslands 2018, 80, 207–214. [Google Scholar]
- Bryant, R.H.; Snow, V.O.; Shorten, P.R.; Welten, B.G. Can alternative forages substantially reduce N leaching? Findings from a review and associated modelling. N. Z. J. Agric. Res. 2019, 63, 3–28. [Google Scholar] [CrossRef]
- Gregorini, P.; Beukes, P.C.; Dalley, D.; Romera, A.J. Screening for diets that reduce urinary nitrogen excretion and methane emissions while maintaining or increasing production by dairy cows. Sci. Total Environ. 2016, 551–552, 32–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welten, B.G.; Ledgard, S.F.; Judge, A.A.; Sprosen, M.S.; McGowan, A.W.; Dexter, M.M. Efficacy of different temperate pasture species to reduce nitrogen leaching from cattle urine applied in different seasons: A soil lysimeter study. Soil Use Manag. 2019, 35, 653–663. [Google Scholar] [CrossRef]
- Brown, H.E.; Moot, D.J.; Pollock, K.M. Long term growth rates and water extraction patterns of dryland chicory, lucerne and red clover. In Legumes for Dryland Pastures; Moot, D.J., Ed.; New Zealand Grassland Association: New Zealand, 2003; pp. 91–100. [Google Scholar]
- Lee, J.M.; Hemmingson, N.R.; Minnee, E.M.K.; Clark, C.E.F. Management strategies for chicory (Cichorium intybus) and plantain (Plantago lanceolata): Impact on dry matter yield, nutritive characteristics and plant density. Crop Pasture Sci. 2015, 66, 168–183. [Google Scholar] [CrossRef]
- Rumball, W.; Keogh, R.G.; Miller, J.E.; Claydon, R.B. Choice’ forage chicory (Cichorium intybus L.). N. Z. J. Agric. Res. 2003, 46, 49–51. [Google Scholar] [CrossRef] [Green Version]
- Mangwe, M.C.; Bryant, R.H.; Beck, M.R.; Beale, N.; Bunt, C.; Gregorini, P. Forage herbs as an alternative to ryegrass-white clover to alter urination patterns in grazing dairy systems. Anim. Feed Sci. Technol. 2019, 252, 11–22. [Google Scholar] [CrossRef]
- Mangwe, M.; Bryant, R.; Gregorini, P. Rumen Fermentation and Fatty Acid Composition of Milk of Mid Lactating Dairy Cows Grazing Chicory and Ryegrass. Animals 2020, 10, 169. [Google Scholar] [CrossRef] [Green Version]
- Minneé, E.M.K.; Waghorn, G.C.; Lee, J.M.; Clark, C.E.F. Including chicory or plantain in a perennial ryegrass/white clover-based diet of dairy cattle in late lactation: Feed intake, milk production and rumen digestion. Anim. Feed Sci. Technol. 2017, 227, 52–61. [Google Scholar] [CrossRef]
- Muir, S.K.; Ward, G.N.; Jacobs, J.L. Herbage intake and milk production of late-lactation dairy cows offered a second-year chicory crop during summer. J. Dairy Sci. 2015, 98, 8825–8835. [Google Scholar] [CrossRef] [Green Version]
- Mangwe, M.; Bryant, R.H.; Beck, M.R.; Fleming, A.E.; Gregorini, P. Grazed chicory, plantain or ryegrass–white clover alters milk yield and fatty acid composition of late-lactating dairy cows. Anim. Prod. Sci. 2020, 60, 107–113. [Google Scholar] [CrossRef]
- Li, G.; Kemp, P.D. Forage Chicory (Cichorium intybus L.): A Review of Its Agronomy and Animal Production. Adv. Agron. 2005, 88, 187–222. [Google Scholar]
- Chen, A.; Bryant, R.H.; Edwards, G.R. Morphology and nutritive value of perennial ryegrass cultivars at different phenological stages. Grass Forage Sci. 2019, 74, 576–581. [Google Scholar] [CrossRef]
- Ta, H.T.; Teixeira, E.I.; Brown, H.E.; Moot, D.J. Yield and quality changes in lucerne of different fall dormancy ratings under three defoliation regimes. Eur. J. Agron. 2020, 115, 126012. [Google Scholar] [CrossRef]
- Gregorini, P.; Minnee, E.M.K.; Griffiths, W.; Lee, J.M. Dairy cows increase ingestive mastication and reduce ruminative chewing when grazing chicory and plantain. J. Dairy Sci. 2013, 96, 7798–7805. [Google Scholar] [CrossRef]
- Clark, D.A.; Anderson, C.B.; Hongwen, G. Liveweight gain and intake of friesian bulls grazing ‘grasslands puna’ chicory (Cichorium intybus L.) or pasture. N. Z. J. Agric. Res. 1990, 33, 219–224. [Google Scholar] [CrossRef]
- Clapham, W.M.; Fedders, J.M.; Belesky, D.P.; Foster, J.G. Developmental Dynamics of Forage Chicory. Agron. J. 2001, 93, 443–450. [Google Scholar] [CrossRef]
- Barry, T.N. The feeding value of chicory (Cichorium intybus) for ruminant livestock. J. Agric. Sci. 1998, 131, 251–257. [Google Scholar] [CrossRef]
- Li, G.D.; Kemp, P.D.; Hodgson, J. Herbage production and persistence of Puna chicory (Cichorium intybus L.) under grazing management over 4 years. N. Z. J. Agric. Res. 1997, 40, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Li, G.D.; Kemp, P.D.; Hodgson, J. Regrowth, morphology and persistence of Grasslands Puna chicory (Cichorium intybus L.) in response to grazing frequency and intensity. Grass Forage Sci. 1997, 52, 33–41. [Google Scholar] [CrossRef]
- Labreveux, M.; Hall, M.H.; Sanderson, M.A. Productivity of chicory and plantain cultivars under grazing. Agron. J. 2004, 96, 710–716. [Google Scholar] [CrossRef] [Green Version]
- Glassey, C.B.; Clark, C.E.F.; Roach, C.G.; Lee, J.M. Herbicide application and direct drilling improves establishment and yield of chicory and plantain. Grass Forage Sci. 2013, 68, 178–185. [Google Scholar] [CrossRef]
- Monti, A.; Brugnoli, E.; Scartazza, A.; Amaducci, M.T. The effect of transient and continuous drought on yield, photosynthesis and carbon isotope discrimination in sugar beet (Beta vulgaris L.). J. Exp. Bot. 2006, 57, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cranston, L.M.; Kenyon, P.R.; Morris, S.T.; Lopez-Villalobos, N.; Kemp, P.D. Morphological and Physiological Responses of Plantain (Plantago lanceolata) and Chicory (Cichorium intybus) to Water Stress and Defoliation Frequency. J. Agron. Crop Sci. 2016, 202, 13–24. [Google Scholar] [CrossRef]
- Lee, J.M.; Minnee, E.M.K.; Clark, C.E.F. Patterns in non-structural carbohydrate and nitrogen reserves in chicory (Cichorium intybus L.) and plantain (Plantago lanceolata L.) during regrowth in summer. Crop Pasture Sci. 2015, 66, 1071–1078. [Google Scholar] [CrossRef]
- Moreno García, C.A.; Schellberg, J.; Ewert, F.; Brüser, K.; Canales-Prati, P.; Linstädter, A.; Oomen, R.J.; Ruppert, J.C.; Perelman, S.B. Response of community-aggregated plant functional traits along grazing gradients: Insights from African semi-arid grasslands. Appl. Veg. Sci. 2014, 17, 470–481. [Google Scholar] [CrossRef]
- Gastal, F.; Lemaire, G. Defoliation, Shoot Plasticity, Sward Structure and Herbage Utilization in Pasture: Review of the Underlying Ecophysiological Processes. Agriculture 2015, 5, 1146–1171. [Google Scholar] [CrossRef] [Green Version]
- Violle, C.; Navas, M.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E.; Oikos, S.; May, N.; Violle, C.; et al. Let the Concept of Trait Be Functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Duru, M.; Ducrocq, H. Growth and senescence of the successive grass leaves on a tiller. Ontogenic development and effect of temperature. Ann. Bot. 2000, 85, 635–643. [Google Scholar] [CrossRef]
- Durand, J.L.; Schäufele, R.; Gastal, F. Grass leaf elongation rate as a function of developmental stage and temperature: Morphological analysis and modelling. Ann. Bot. 1999, 83, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Granier, C.; Massonnet, C.; Turc, O.; Muller, B.; Chenu, K.; Tardieu, F. Individual leaf development in Arabidopsis thaliana: A stable thermal-time-based programme. Ann. Bot. 2002, 89, 595–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, A.; Schurr, U. Dynamics of leaf and root growth: Endogenous control versus environmental impact. Ann. Bot. 2005, 95, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.M.; Kemp, P.D.; Jaya, I.K.D.; Osborne, M.A. Establishment, growth and development of plantain and chicory under grazing. Proc. N. Z. Grassl. Assoc. 2007, 69, 41–45. [Google Scholar]
- McMaster, G.S.; Wilhelm, W.W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Cranston, L.M.; Kenyon, P.R.; Morris, S.T.; Lopez-Villalobos, N.; Kemp, P.D. Effect of post-grazing height on the productivity, population and morphology of a herb and legume mix. N. Z. J. Agric. Res. 2015, 58, 397–411. [Google Scholar] [CrossRef]
- Pollock, C.J.; Jones, T. Seasonal patterns of fructan metabolism in forage grasses. New Phytol. 1979, 83, 9–15. [Google Scholar] [CrossRef]
- Yuan, Z.; Cao, Q.; Zhang, K.; Ata-Ul-Karim, S.T.; Tan, Y.; Zhu, Y.; Cao, W.; Liu, X. Optimal leaf positions for SPAD meter measurement in rice. Front. Plant Sci. 2016, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lüdecke, D. ggeffects: Tidy Data Frames of Marginal Effects from Regression Models. J. Open Source Softw. 2018, 3, 772. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R package Version 1.0.7. Available online: http://www.sthda.com/english/rpkgs/factoextra (accessed on 2 May 2020).
- Rumball, W. Grasslands puna’ chicory (Cichorium intybus L.). N. Z. J. Exp. Agric. 1986, 14, 105–107. [Google Scholar] [CrossRef]
- Cichota, R.; McAuliffe, R.; Lee, J.; Minnee, E.; Martin, K.; Brown, H.E.; Moot, D.J.; Snow, V.O. Forage chicory model: Development and evaluation. F. Crop. Res. 2020, 246, 107633. [Google Scholar] [CrossRef]
- Li, G.D.; Kemp, P.D.; Hodgson, J. Control of reproductive growth in Puna chicory by grazing management. Proc. N. Z. Grassl. Assoc. 1994, 56, 213–217. [Google Scholar]
- McNaughton, S.J. Compensatory Plant Growth as a Response to Herbivory. Oikos 1983, 40, 329. [Google Scholar] [CrossRef]
- Briske, D.D.; Richards, J.H. Plant responses to defoliation: A physiological, morphological and demographic evaluation. In Wildland Plants: Physiological Ecology and Developmental Biology; Bedunah, D., Sosebee, R., Eds.; Society for Range Management: Denver, CO, USA, 1995; pp. 635–710. [Google Scholar]
- Kebrom, T.H. A Growing Stem Inhibits Bud Outgrowth–The Overlooked Theory of Apical Dominance. Front. Plant Sci. 2017, 8, 1874. [Google Scholar] [CrossRef]
- Leach, G.J. Regrowth characteristics of lucerne under different systems of grazing management. Aust. J. Agric. Res. 1979, 30, 445–465. [Google Scholar] [CrossRef]
- Ong, C.K. The physiology of tiller death in grasses. 1. The influence of tiller age, size and position. Grass Forage Sci. 1978, 33, 197–203. [Google Scholar] [CrossRef]
- Smith, A.E.; Leinweber, C.L. Incorporation of 14C by Little Bluestem Tillers at Two Stages of Phenological Development 1. Agron. J. 1973, 65, 908–910. [Google Scholar] [CrossRef]
- Solomon, J.K.Q.; Macoon, B.; Lang, D.J. Harvest management based on leaf stage of a tetraploid vs. a diploid cultivar of annual ryegrass. Grass Forage Sci. 2017, 72, 743–756. [Google Scholar] [CrossRef] [Green Version]
- Doorenbos, J.; Riemens, P. Effect of Vernalization and Daylength on Number and Shape of Leaves in Chicory and Endive. Acta Bolanica Neerl. 1959, 8, 63–67. [Google Scholar] [CrossRef]
- Vallet, C.; Lemaire, G.; Monties, B.; Chabbert, B. Cell Wall Fractionation of Alfalfa Stem in Relation to Internode Development: Biochemistry Aspect. J. Agric. Food Chem. 1998, 46, 3458–3467. [Google Scholar] [CrossRef]
- Shi, P.; Liu, M.; Yu, X.; Gielis, J.; Ratkowsky, D.A. Proportional relationship between leaf area and the product of leaf length and width of four types of special leaf shapes. Forests 2019, 10, 178. [Google Scholar] [CrossRef] [Green Version]
- Holland, J.N.; Cheng, W.; Crossley, D.A. Herbivore-induced changes in plant carbon allocation: Assessment of below-ground C fluxes using carbon-14. Oecologia 1996, 107, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Donaghy, D.J.; Fulkerson, W.J. The importance of water-soluble carbohydrate reserves on regrowth and root growth of Lolium perenne (L.). Grass Forage Sci. 1997, 52, 401–407. [Google Scholar] [CrossRef]
- Donaghy, D.J.; Fulkerson, W.J. Priority for allocation of water-soluble carbohydrate reserves during regrowth of Lolium perenne. Grass Forage Sci. 1998, 53, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Kendall, W.A. The Persistence of Red Clover and Carbohydrate Concentration in the Roots at Various Temperatures. Agron. J. 1958, 50, 657–659. [Google Scholar] [CrossRef]
- Hunt, W.F.; Hay, J.M. A photographic technique for assessing the pasture species performance of grazing animals. Proc. N. Z. Grassl. Assoc. 1990, 51, 191–198. [Google Scholar]
- McCoy, J.E.; Collins, M.; Dougherty, C.T. Amount and quality of chicory herbage ingested by grazing cattle. Crop Sci. 1997, 37, 239–242. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Total Precipitation (mm) | Irrigation (mm) | Avg. Max Temp (°C) | Avg. Min Temp (°C) | |||
|---|---|---|---|---|---|---|---|
| During Exp. | 10-yrs Mean | During Exp. | During Exp. | 10-yrs Mean | During Exp. | 10-yrs Mean | |
| Feb-18 | 123 | 28.3 | 17.8 | 22.5 | 22.0 | 12.1 | 11.8 |
| Mar-18 | 31.6 | 57.1 | 0.4 | 20.7 | 20.1 | 11.4 | 10.0 |
| Apr-18 | 91.0 | 65.6 | 2.3 | 17.6 | 17.7 | 6.7 | 7.8 |
| May-18 | 47.6 | 77.0 | 0 | 14.3 | 14.8 | 5.4 | 5.1 |
| Jun-18 | 58.8 | 72.8 | 0 | 10.6 | 11.9 | 3.8 | 2.5 |
| Jul-18 | 26.2 | 43.9 | 0 | 13.0 | 11.4 | 2.3 | 1.5 |
| Aug-18 | 15.8 | 57.6 | 0 | 13.0 | 12.7 | 3.6 | 3.6 |
| Sep-18 | 39.4 | 34.6 | 0 | 14.3 | 14.6 | 4.7 | 4.8 |
| Oct-18 | 55.6 | 50.8 | 10.5 | 16.9 | 16.6 | 6.1 | 6.5 |
| Nov-18 | 107.4 | 33.0 | 2.1 | 18.1 | 18.8 | 8.6 | 8.3 |
| Dec-18 | 57.0 | 44.2 | 2.1 | 19.4 | 20.9 | 12.3 | 11.0 |
| Jan-19 | 36.2 | 44.5 | 24.1 | 23.9 | 22.1 | 13.1 | 11.8 |
| Feb-19 | 29.2 | 28.3 | 44.8 | 24.3 | 22.0 | 11.6 | 11.8 |
| Mar-19 | 23.8 | 57.1 | 14.8 | 21.8 | 20.1 | 11.9 | 10.0 |
| Total/Ave | 742.6 | 694.8 | 118.9 | 17.9 | 17.6 | 8.1 | 7.6 |
| Year | Harvest Period | Defoliation Intervals | |||
|---|---|---|---|---|---|
| 300 GDD | 600 GDD | ||||
| Date | Accumulated GDD | Date | Accumulated GDD | ||
| 2018 | 1 | 15-Feb | 15-Feb | ||
| 2 | 26-Mar | 480 | |||
| 3 | 28-Apr | 298 1 | 6-Apr | 609 1 | |
| 4 | 5-Oct | 740 | 5-Oct | 907 | |
| 5 | 13-Nov | 320 1 | |||
| 6 | 13-Dec | 298 1 | 13-Dec | 636 1 | |
| 2019 | 7 | 8-Jan | 313 1 | ||
| 8 | 5-Feb | 402 1 | 5-Feb | 715 1 | |
| Interval (I) 1 | Height (H) 1 | Age (A) 1 | I × H | I × A | H × A | I × H × A | |
|---|---|---|---|---|---|---|---|
| At individual Plant Level | |||||||
| Number of leaves | 0.006 | 0.777 | <0.0001 | 0.319 | 0.048 | 0.728 | 0.351 |
| Number of primary shoots | <0.001 | 0.996 | <0.001 | 0.706 | <0.001 | 0.281 | 0.304 |
| Number of secondary shoots | <0.001 | 0.894 | <0.001 | 0.240 | 0.001 | 0.796 | 0.240 |
| Total shoots per plant | <0.001 | 0.748 | <0.0001 | 0.193 | <0.001 | 0.423 | 0.308 |
| Absolute leaf mass (g DM 2) | <0.001 | 0.275 | 0.294 | 0.965 | <0.001 | 0.249 | 0.233 |
| Absolute stem mass (g DM) | <0.001 | 0.091 | <0.001 | 0.571 | <0.001 | 0.091 | 0.571 |
| Total plant weight (g DM) | <0.001 | 0.465 | <0.001 | 0.652 | 0.073 | 0.484 | 0.841 |
| Root diameter (mm) | <0.001 | 0.203 | <0.001 | 0.032 | 0.764 | 0.911 | 0.858 |
| Roots WSC 3 (g/kg DM) | <0.001 | 0.149 | <0.001 | 0.356 | 0.211 | 0.86 | 0.77 |
| At Pasture Level | |||||||
| Pre graze mass (kg/ha DM) | <0.001 | 0.522 | 0.678 | 0.600 | 0.566 | 0.580 | 0.536 |
| Post graze mass (kg/ha DM) | 0.961 | 0.014 | <0.001 | 0.798 | 0.033 | 0.434 | 0.306 |
| Accumulated herbage mass | <0.001 | 0.646 | <0.001 | 0.467 | 0.002 | 0.771 | 0.724 |
| Plant density (plants/m 2) | 0.778 | 0.496 | <0.001 | 0.695 | 0.643 | 0.343 | 0.253 |
| Interval | 300 GDD 1 | 600 GDD 1 | SEM 2 | ||
|---|---|---|---|---|---|
| Height | 4 cm | 8 cm | 4 cm | 8 cm | |
| Before Vernalisation (February–May 2018) | |||||
| Number of leaves (leaves/plant) | 15.2 | 13.8 | 13.9 | 15.4 | 2.11 |
| Number of primary shoots (shoots/plant) | 1.58 | 1.25 | 1.33 | 1.25 | 0.31 |
| Number of secondary shoots (shoots/plant) | 0 | 0 | 0 | 0 | |
| Total shoots per plant (shoots/plant) | 1.58 | 1.25 | 1.33 | 1.25 | 0.7 |
| Absolute leaf mass (g DM) | 2.44 b | 2.24 b | 3.57 a | 3.78 a | 0.26 |
| Absolute stem mass (g DM) | 0 | 0 | 0 | 0 | |
| Total plant weight (g DM) | 2.44 b | 2.24 b | 3.57 a | 3.78 a | 0.51 |
| Root diameter (mm) | 13.4 b | 14.7 b | 16.5 b | 18.8 a | 0.83 |
| Roots water soluble carbohydrates (g/kg DM) | 756 b | 766 b | 803 a | 870 a | 38.1 |
| After Vernalisation (October 2018–February 2019) | |||||
| Number of leaves (leaves/plant) | 31.6 b | 29.9 b | 38.3 a | 35.0 a | 1.49 |
| Number of primary shoots (shoots/plant) | 3.99 a | 4.46 a | 2.19 b | 2.12 b | 0.22 |
| Number of secondary shoots (shoots/plant) | 1.38 b | 0.38 b | 6.97 a | 8.53 a | 0.62 |
| Total shoots per plant (shoots/plant) | 5.37 b | 4.84 b | 9.16 a | 10.7 a | 0.51 |
| Absolute leaf mass (g DM) | 3.53 | 3.19 | 3.01 | 2.98 | 0.18 |
| Absolute stem mass (g DM) | 1.23 b | 1.77 b | 3.22 a | 4.30 a | 0.27 |
| Total plant weight (g DM) | 4.76 b | 4.96 b | 6.23 a | 7.28 a | 0.39 |
| Root diameter (mm) | 18.1 c | 19.0 c | 22.8 b | 26.8 a | 1.30 |
| Roots water soluble carbohydrates (g/kg DM) | 402 b | 417 b | 522 a | 568 a | 38.1 |
| Interval | 300 GDD 1 | 600 GDD 1 | SEM 3 | ||
|---|---|---|---|---|---|
| Height | 4 cm | 8 cm | 4 cm | 8 cm | |
| Before vernalisation (February–May 2018) | |||||
| Pre graze mass (kg/ha DM) | 3510 b | 3293 b | 4648 a | 5032 a | 415 |
| Post graze mass (kg/ha DM) | 627 | 774 | 586 | 687 | 111 |
| Accumulated herbage mass (kg/ha DM) | 4192 | 4242 | 4637 | 4358 | 213 |
| Plant density (plants/m2) | 127 | 124 | 130.5 | 122 | 8.25 |
| After vernalisation (October 2018–February 2019) | |||||
| Pre graze mass (kg/ha DM) | 3510 b | 3659 b | 5248 a | 5407 a | 293 |
| Post graze mass (kg/ha DM) | 944 c | 1077 b | 973 c | 1182 a | 78.8 |
| Accumulated herbage mass (kg/ha DM) | 10,284 b | 10,316 b | 11,659 a | 11,576 a | 320 |
| Plant density (plants/m2) 2 | 73 | 71 | 65 | 68 | 6.36 |
| Item | 600 GDD 1 | 300 GDD 1 | SEM 5 | p-Value |
|---|---|---|---|---|
| Number of live leaves per plant | 42.6 | 28.2 | 4.13 | 0.019 |
| Number of primary shoots per plant | 2.10 | 3.79 | 0.34 | 0.011 |
| Number of secondary shoots per plant 3 | 8.40 ± 0.4 | - | - | - |
| Absolute leaf mass per plant (g DM2) | 2.36 | 3.05 | 0.56 | 0.269 |
| Absolute stem mass (g DM2) 3 | 2.99 ± 1.2 | - | - | - |
| Final stem lenght (mm) 3 | 370 ± 160 | - | - | - |
| Absolute dead material mass (g DM2) | 0.87 | 0.75 | 0.11 | 0.213 |
| Root diameter (mm) | 21.4 | 19.1 | 1.22 | 0.272 |
| SPAD 4 | 37.5 | 29.2 | 1.41 | <0.001 |
| Specific leaf area (cm2/g) | 18.6 | 29.3 | 2.63 | 0.007 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mangwe, M.C.; Bryant, R.H.; Moreno García, C.A.; Maxwell, T.M.R.; Gregorini, P. Functional Traits, Morphology, and Herbage Production of Vernalised and Non-Vernalised Chicory cv. Choice (Cichorium intybus L.) in Response to Defoliation Frequency and Height. Plants 2020, 9, 611. https://doi.org/10.3390/plants9050611
Mangwe MC, Bryant RH, Moreno García CA, Maxwell TMR, Gregorini P. Functional Traits, Morphology, and Herbage Production of Vernalised and Non-Vernalised Chicory cv. Choice (Cichorium intybus L.) in Response to Defoliation Frequency and Height. Plants. 2020; 9(5):611. https://doi.org/10.3390/plants9050611
Chicago/Turabian StyleMangwe, Mancoba C., Racheal H. Bryant, Cristian A. Moreno García, Thomas M.R. Maxwell, and Pablo Gregorini. 2020. "Functional Traits, Morphology, and Herbage Production of Vernalised and Non-Vernalised Chicory cv. Choice (Cichorium intybus L.) in Response to Defoliation Frequency and Height" Plants 9, no. 5: 611. https://doi.org/10.3390/plants9050611