Emerging Advanced Technologies to Mitigate the Impact of Climate Change in Africa


{kind=link}
{kind=link}
Abstract
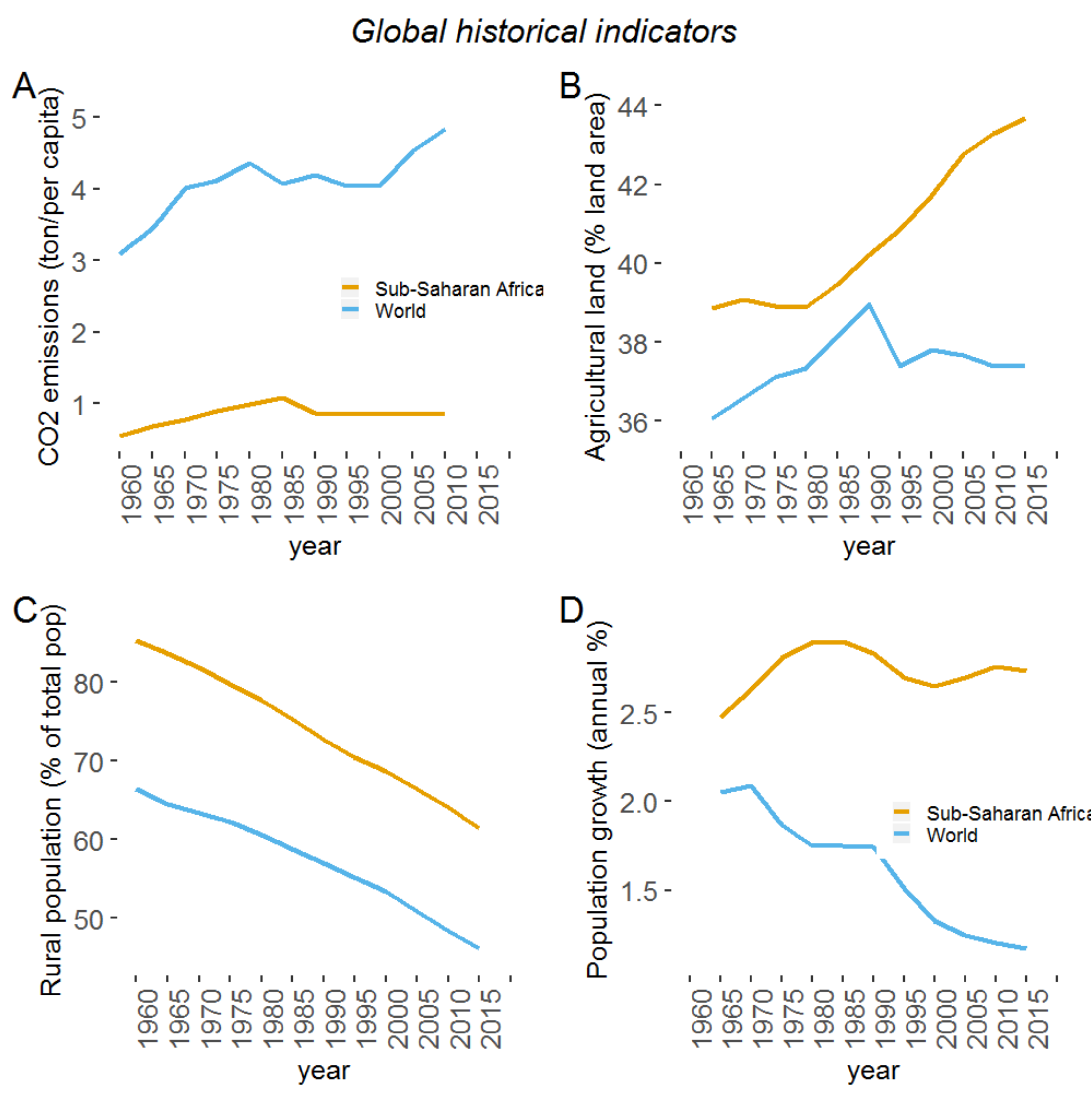
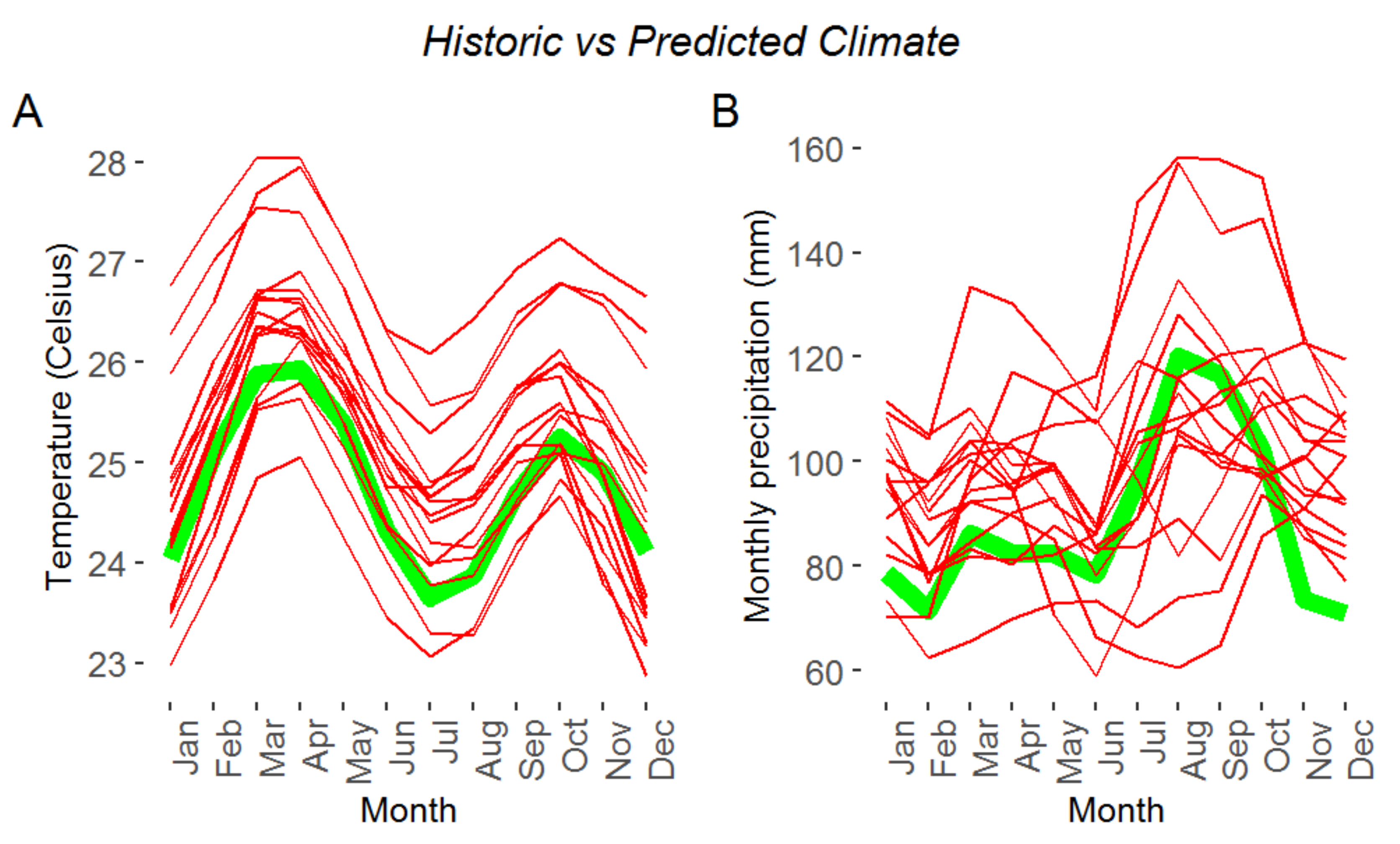
:1. Introduction
2. Resources in Plant Breeding
3. Advances in Genomic Selection
4. Advances in Plant Phenotyping
5. Modelling and Computational Biology Applications
6. Synthetic Biology
7. Computational Resources on Plant Genomics
8. Challenges and the Way Forward
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Girvetz, E.; Ramirez-Villegas, J.; Claessens, L.; Lamanna, C.; Navarro-Racines, C.; Nowak, A.; Thornton, P.; Rosenstock, T.S.; Rosenstock, T.S.; Nowak, A.; et al. Future Climate Projections in Africa: Where Are We Headed? The Climate-Smart Agriculture Papers: Investigating the Business of a Productive, Resilient and Low Emission Future; Springer International Publishing: Cham, Swizerland, 2019. [Google Scholar]
- Niang, I.; Ruppel, O.C.; Abdrabo, M.A.; Essel, A.; Lennard, C.; Padgham, J.; Urquhart, P. Africa. In ClimateChange 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working GroupII to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Barros, V.R., Field, C.B., Dokken, D.J., Mastrandrea, M.D., Mach, K.J., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1199–1265. [Google Scholar]
- Ofoegbu, C.; Chirwa, P.W.; Francis, J.; Babalola, F.D. Assessing local-level forest use and management capacity as a climate-change adaptation strategy in Vhembe district of South Africa. Clim. Dev. 2019, 11, 501–512. [Google Scholar] [CrossRef]
- Mueller, B.; Seneviratne, S.I. Hot days induced by precipitation deficits at the global scale Proceedings of the National Academy of Sciences. Natl. Acad. Sci. 2012, 109, 12398–12403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rippke, U.; Ramirez-Villegas, J.; Jarvis, A.; Vermeulen, S.J.; Parker, L.; Mer, F.; Diekkrüger, B.; Challinor, A.J.; Howden, M. Timescales of transformational climate change adaptation in sub-Saharan African agriculture. Nat. Clim. Chang. 2016, 6, 605–609. [Google Scholar] [CrossRef]
- Ahmed, S.; Griffin, T.S.; Kraner, D.; Schaffner, M.K.; Sharma, D.; Hazel, M.; Leitch, A.R.; Orians, C.M.; Han, W.; Stepp, J.R.; et al. Environmental Factors Variably Impact Tea Secondary Metabolites in the Context of Climate Change. Front. Plant Sci. 2019, 10, 939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, W.; Tao, F. Vulnerability of African maize yield to climate change and variability during 1961–2010. Food Secur. 2014, 6, 471–481. [Google Scholar] [CrossRef]
- Rurinda, J.; Van Wijk, M.T.; Mapfumo, P.; Descheemaeker, K.; Supit, I.; Giller, K.E. Climate change and maize yield in southern Africa: What can farm management do? Glob. Chang. Biol. 2015, 21, 4588–4601. [Google Scholar] [CrossRef]
- Vogel, C.; Steynor, A.; Manyuchi, A. Climate services in Africa: Re-imagining an inclusive, robust and sustainable service. Clim. Serv. 2019, 15, 100107. [Google Scholar] [CrossRef]
- Harfouche, A.L.; Jacobson, D.A.; Kainer, D.; Romero, J.C.; Harfouche, A.H.; Mugnozza, G.S.; Moshelion, M.; Tuskan, G.A.; Keurentjes, J.J.B.; Altman, A. Accelerating climate resilient plant breeding by applying next-generation artificial intelligence. Trends Biotechnol. 2019, 37, 1217–1235. [Google Scholar] [CrossRef]
- Batley, J.; Edwards, D. The application of genomics and bioinformatics to accelerate crop improvement in a changing climate. Curr. Opin. Plant Biol. 2016, 30, 78–81. [Google Scholar] [CrossRef] [Green Version]
- Mortimer, J.C. Plant synthetic biology could drive a revolution in biofuels and medicine. Exp. Biol. Med. 2019, 244, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.G.; Rodgers-Melnick, E.; Buckler, E.S. On the Road to Breeding 4.0: Unraveling the Good, the Bad, and the Boring of Crop Quantitative Genomics. Ann. Rev. Genet. 2018, 52, 421–444. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Adamski, N.M. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lander, E.S.; Botstein, D. Mapping Mendelian factors underlying quantitative traits using RFLP markers. Genetics 1989, 121, 185–199. [Google Scholar] [PubMed]
- Risch, N.; Merikangas, K.M. The future of genetic studies of complex human diseases. Science 1996, 273, 1516–1517. [Google Scholar] [CrossRef] [Green Version]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. Int. Soc. Microb. Ecol. 2013, 8, 790–803. [Google Scholar] [CrossRef] [Green Version]
- Montesinos-López, O.A.; Martín-Vallejo, J.; Crossa, J.; Gianola, D.; Hernández-Suárez, C.M.; Montesinos-López, A.; Juliana, P.; Singh, R. New Deep Learning Genomic-Based Prediction Model for Multiple Traits with Binary, Ordinal, and Continuous Phenotypes. G3 2019, 9, 1545–1556. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Hu, S.; Gardner, C.; Lübberstedt, T. Trends. Plant Sci. 2017, 22, 624–637. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Pan, Q.; He, F.; Akhunova, A.; Chao, S.; Trick, H.; Akhunov, E. Transgenerational CRISPR-Cas9 Activity Facilitates Multiplex Gene Editing in Allopolyploid Wheat. CRISPR J. 2018, 1, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J.; Bartlett, M.E.; Lippman, Z.B. Engineering Quantitative Trait Variation for Crop Improvement by Genome Editing. Cell 2017, 171, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Turuspekov, Y.; Baibulatova, A.; Yermekbayev, K.; Tokhetova, L.; Chudinov, V.; Sereda, G.; Ganal, M.; Griffiths, S.; Abugalieva, S. GWAS for plant growth stages and yield components in spring wheat (Triticum aestivum L.) harvested in three regions of Kazakhstan BMC plant biology. Biomed. Cent. 2017, 17, 190. [Google Scholar] [CrossRef] [Green Version]
- Ray, D.K.; Ramankutty, N.; Mueller, N.D.; West, P.C.; Foley, J.A. Recent patterns of crop yield growth and stagnation. Nat. Commun. 2012, 3, 1293. [Google Scholar] [CrossRef] [Green Version]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassi, F.M.; Bentley, A.R.; Charmet, G.; Ortiz, R.; Crossa, J. Breeding schemes for the implementation of genomic selection in wheat (Triticum spp.). Plant Sci. 2016, 242, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Stuber, C.W.; Goodman, M.M.; Moll, R.H. Improvement of yield and ear number resulting from selection at allozyme loci in a maize population. Crop Sci. 1982, 22, 737–740. [Google Scholar] [CrossRef]
- Tanksley, S.D.; Young, N.D.; Paterson, A.H.; Bonierbale, M.W. RFLP mapping in plant breeding: New tools for an old science. Biotechnology 1989, 7, 257–264. [Google Scholar] [CrossRef]
- Bernardo, R. Molecular markers and selection for complex traits in plants: Learning from the last 20 years. Crop Sci. 2008, 48, 1649–1664. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Crouch, J.H. Marker-assisted selection in plant breeding: From publications to practice. Crop Sci. 2008, 48, 391–407. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.; Toubia-Rahme, H.; Steffenson, B.J.; Smith, K.P. Molecular Mapping and Marker-Assisted Selection of Genes for Septoria Speckled Leaf Blotch Resistance in Barley. Phytopathology 2006, 96, 993–999. [Google Scholar] [CrossRef] [Green Version]
- Moreau, L.; Charcosset, A.; Gallais, A. Experimental evaluation of several cycles of marker-assisted selection in maize. Euphytica 2004, 137, 111–118. [Google Scholar] [CrossRef]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar]
- Heffner, E.L.; Sorrells, M.E.; Jannink, J.L. Genomic selection for crop improvement. Crop Sci. 2009, 49, 1–12. [Google Scholar] [CrossRef]
- Su, G.; Guldbrandtsen, B.; Gregersen, V.R.; Lund, M.S. Preliminary investigation on reliability of genomic estimated breeding values in the Danish Holstein population. J. Dairysci. 2010, 93, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- Heffner, E.L.; Lorenz, A.J.; Jannink, J.L.; Sorrells, M.E. Plant breeding with genomic selection: Gain per unit time and cost. Crop Sci. 2010, 50, 1681–1690. [Google Scholar] [CrossRef]
- Bernardo, R.; Yu, J.M. Prospects for genomewide selection for quantitative traits in maize. Crop Sci. 2007, 47, 1082–1090. [Google Scholar] [CrossRef] [Green Version]
- Crossa, J.; Perez, P.; Hickey, J.; Burgueno, J.; Ornella, L.; Ceron-Rojas, J.; Zhang, X.; Dreisigacker, S.; Babu, R.; Li, Y.; et al. Genomic prediction in CIMMYT maize and wheat breeding programs. Heredity 2014, 112, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, A.J.; Smith, K.P.; Jannink, J.L. Potential and optimization of genomic selection for fusarium head blight resistance in six-row barley. Crop Sci. 2012, 52, 1609–1621. [Google Scholar] [CrossRef]
- Asoro, F.G.; Newell, M.A.; Beavis, W.D.; Scott, M.P.; Tinker, N.A.; Jannink, J.L. Genomic, marker-assisted, and pedigree-BLUP selection methods for beta-glucan concentration in elite oat. Crop Sci. 2013, 53, 1894–1906. [Google Scholar] [CrossRef] [Green Version]
- Lariepe, A.; Moreau, L.; Laborde, J.; Bauland, C.; Mezmouk, S.; Decousset, L.; Mary-Huard, T.; Fievet, J.B.; Gallais, A.; Dubreuil, P.; et al. General and specific combining abilities in a maize (Zea mays L.) test-cross hybrid panel: Relative importance of population structure and genetic divergence between parents. Appl. Genet. 2017, 130, 403–417. [Google Scholar] [CrossRef]
- Riedelsheimer, C.; Czedik-Eysenberg, A.; Grieder, C.; Lisec, J.; Technow, F.; Sulpice, R.; Altmann, T.; Stitt, M.; Willmitzer, L.; Melchinger, A.E. Genomic and metabolic prediction of complex heterotic traits in hybrid maize. Nat. Genet. 2012, 44, 217–220. [Google Scholar] [CrossRef]
- Cooper, M.; Gho, C.; Leafgren, R.; Tang, T.; Messina, C. Breeding drought-tolerant maize hybrids for the US corn-belt: Discovery to product. J. Exp. Bot. 2014, 65, 6191–6204. [Google Scholar] [CrossRef] [Green Version]
- Gaffney, J.; Schussler, J.; Löffler, C.; Cai, W.; Paszkiewicz, S.; Messina, C.D.; Groeteke, J.; Keaschall, J.; Cooper, M. Industry-Scale Evaluation of Maize Hybrids Selected for Increased Yield in Drought-Stress Conditions of the US Corn Belt. Crop. Sci. 2015, 55, 1608–1618. [Google Scholar] [CrossRef]
- Crain, J.; Mondal, S.; Rutkoski, J.; Singh, R.P.; Poland, J. Combining high-throughput phenotyping and genomic information to increase prediction and selection accuracy in wheat breeding. Plant Genome 2018, 11, 170043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, B.; Panozzo, J.; Walker, C.K.; Choy, A.L.; Kant, S.; Wong, D.; Tibbits, J.; Daetwyler, H.D.; Rochfort, S.; Hayden, M.; et al. Accelerating wheat breeding for end-use quality with multi-trait genomic predictions incorporating near infrared and nuclear magnetic resonance-derived phenotypes. Theor. Appl. Genet. 2017, 130, 2505–2519. [Google Scholar] [CrossRef] [PubMed]
- Riaz, A.; Periyannan, S.; Aitken, E.; Hickey, L. A rapid phenotyping method for adult plant resistance to leaf rust in wheat. Plant Methods 2016, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Schilling, C.H.; Edwards, J.S.; Palsson, B.O. Toward Metabolic Phenomics: Analysis of Genomic Data Using Flux Balances. Biotechnol. Prog. 1999, 15, 288–295. [Google Scholar] [CrossRef]
- Houle, D.; Govindaraju, D.R.; Omholt, S. Phenomics: The Next Challenge Nature Reviews Genetics; Nature Publishing Group: New York, NY, USA, 2010. [Google Scholar]
- Furber, S. Correction to ‘Microprocessors: The engines of the digital age’. Proc. Math. Phys. Eng. Sci. 2017, 473, 20160893. [Google Scholar] [CrossRef] [Green Version]
- Tester, M.; Langridge, P. Breeding Technologies to Increase Crop Production in a Changing World. Science 2010, 12, 818–822. [Google Scholar] [CrossRef]
- Furbank, R.T.; Tester, M. Phenomics—Technologies to relieve the phenotyping bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef]
- Camargo, A.; Gustavo, L.A. Latin America: A Development Pole for Phenomics. Front. Plant Sci. 2016. [Google Scholar] [CrossRef] [Green Version]
- Camargo, A.V.; Mackay, I.; Mott, R.; Han, J.; Doonan, J.H.; Askew, K.; Corke, F.; Williams, K.; Bentley, A.R. Functional Mapping of Quantitative Trait Loci (QTLs) Associated With Plant Performance in a Wheat MAGIC Mapping Population. Front. Plant Sci. 2018, 9, 887. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, R.; Song, W.; Han, L.; Liu, X.; Sun, X.; Luo, M.; Chen, K.; Zhang, Y.; Yang, H.; et al. Dynamic plant height QTL revealed in maize through remote sensing phenotyping using a high-throughput unmanned aerial vehicle (UAV). Sci. Rep. 2019, 9, 3458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Ji, Y.; Fu, Y.; Guo, H. Ethylene-induced microtubule reorientation is essential for fast inhibition of root elongation in Arabidopsis. J. Integr. Plant Biol. 2018, 60, 864–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, A.J.; Srinivasan, V.; Hart, J.C.; Marshall-Colón, A. Use of computational modeling combined with advanced visualization to develop strategies for the design of crop ideotypes to address food security. Nutr Rev. 2018, 76, 332–347. [Google Scholar] [CrossRef] [PubMed]
- Osborne, B. Near-Infrared Spectroscopy in Food Analysis Encyclopedia of Analytical Chemistry; BRI Australia Ltd.: North Ryde, Australia, 2006. [Google Scholar]
- Wesley, J.; Larroque, O.; Osborne, B.G.; Azudin, N.; Allen, H.; Skerritt, J.H. Measurement of gliadin and glutenin content of flour by NIR spectroscopy. J. Cereal. Sci. 2001, 34, 125–133. [Google Scholar] [CrossRef]
- Anderssen, R.; Edwards, M. Mathematical modelling in the science and technology of plant breeding. Int. J. Numer. Anal. Model. Ser. B 2012, 3, 242–258. [Google Scholar]
- Wiley, P.R.; Tanner, G.J.; Chandler, P.M.; Anderssen, R.S. Molecular Classification of Barley (Hordeum vulgare L.) Mutants Using Derivative NIR Spectroscopy. J. Agric. Food Chem. 2009, 57, 4042–4050. [Google Scholar] [CrossRef]
- A framework for genomics-informed ecophysiological modeling in plants. J. Exp. Bot. 2019, 70, 2561–2574. [CrossRef] [Green Version]
- Küken, A.; Nikoloski, Z. Computational Approaches to Design and Test Plant Synthetic Metabolic Pathways. Plant Physiol. 2019, 179, 894. [Google Scholar] [CrossRef] [Green Version]
- Small, I.; Puchta, H. Emerging tools for synthetic biology in plants. Plant J. Cell Mol. Biol. 2014, 78, 725–726. [Google Scholar] [CrossRef] [Green Version]
- Andres, J.; Blomeier, T.; Zurbriggen, M.D. Synthetic switches and regulatory circuits in plants. Plant Physiol. 2019, 179, 862–884. [Google Scholar] [CrossRef] [Green Version]
- Toda, S.; Blauch, L.R.; Tang, S.K.Y.; Morsut, L.; Lim, W.A. Programming self-organizing multicellular structures with synthetic cell-cell signalling. Science 2018, 361, 156–162. [Google Scholar] [PubMed] [Green Version]
- Shaw, W.M.; Yamauchi, H.; Mead, J.; Gowers, G.-O.F.; Bell, D.J.; Öling, D.; Larsson, N.; Wigglesworth, M.; Ladds, G.; Ellis, T. Engineering a model cell for rational tuning of GPCR signalling. Cell 2019, 177, 782–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarrion-Perdigones, E.E.; Falconi, S.I.; Zandalinas, P.; Juárez, A.; Fernández-del-Carmen, A.; Granell, D. Orzaez Golden Braid: An iterative cloning system for standardized assembly of reusable genetic modules. PLoS ONE 2011, 6, e21622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engler, C.; Youles, M.; Gruetzner, R.; Ehnert, T.-M.; Werner, S.; Jones, J.D.G.; Patron, N.J.; Marillonnet, S. A golden gate modular cloning toolbox for plants. ACS Synth. Biol. 2014, 3, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Roell, M.-S.; Zurbriggen, M.D. The impact of synthetic biology for future agriculture and nutrition. Curr. Opin. Biotechnol. 2020, 61, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Synbio Africa. Available online: https://synbioafrica.com/ (accessed on 19 March 2020).
- The International Wheat Genome Sequencing Consortium (IWGSC); Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef]
- Goldstein, S.; Beka, L.; Graf, J.; Klassen, J.L. Evaluation of strategies for the assembly of diverse bacterial genomes using MinION long-read sequencing. BMC Genom. 2019, 20, 23. [Google Scholar] [CrossRef] [Green Version]
- Boykin, L.M.; Sseruwagi, P.; Alicai, T.; Ateka, E.; Mohammed, I.U.; Stanton, J.-A.L.; Kayuki, C.; Mark, D.; Fute, T.; Erasto, J.; et al. Tree Lab: Portable genomics for Early Detection of Plant Viruses and Pests in Sub-Saharan Africa. Genes 2019, 10, 632. [Google Scholar] [CrossRef] [Green Version]
- Rajarammohan, S.; Pental, D.; Kaur, J. Near-Complete Genome Assembly of Alternaria brassicae-A Necrotrophic Pathogen of Brassica Crops. Molecular plant-microbe interactions. MPMI 2019, 32, 928–930. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Poorter, H.; Dumschott, K.; Bolger, M.E.; Arend, D.; Osorio, S.; Gundlach, H.; Mayer, K.F.X.; Lange, M.; Scholz, U.; et al. Computational aspects underlying genome to phenome analysis in plants. Plant J. 2019, 97, 182–198. [Google Scholar] [CrossRef] [PubMed]
- Merchant, N.; Lyons, E.; Goff, S.; Vaughn, M.W.; Ware, R.; Micklos, D.; Antin, P. The iPlant Collaborative: Cyberinfrastructure for Enabling Data to Discovery for the Life Sciences. PLoS Biol. 2016, e1002342. [Google Scholar] [CrossRef] [PubMed] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francisco Ribeiro, P.; Camargo Rodriguez, A.V. Emerging Advanced Technologies to Mitigate the Impact of Climate Change in Africa. Plants 2020, 9, 381. https://doi.org/10.3390/plants9030381
Francisco Ribeiro P, Camargo Rodriguez AV. Emerging Advanced Technologies to Mitigate the Impact of Climate Change in Africa. Plants. 2020; 9(3):381. https://doi.org/10.3390/plants9030381
Chicago/Turabian StyleFrancisco Ribeiro, Priscilla, and Anyela Valentina Camargo Rodriguez. 2020. "Emerging Advanced Technologies to Mitigate the Impact of Climate Change in Africa" Plants 9, no. 3: 381. https://doi.org/10.3390/plants9030381