Identification, Characterization, and Stress Responsiveness of Glucose-6-phosphate Dehydrogenase Genes in Highland Barley


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Identification of HvG6PDH Genes and Bioinformatics
2.3. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR)
2.4. Subcellular Localization
2.5. SDS-PAGE of HvG6PDH Proteins from Escherichia coli
2.6. Extraction and Determination of G6PDH Activity
2.7. Determination of Hydrogen Peroxide and Superoxide Anion Contents
2.8. Statistical Analysis
3. Results
3.1. Identification and Preliminary Characterization of Putative HvG6PDH Genes in Highland Barley
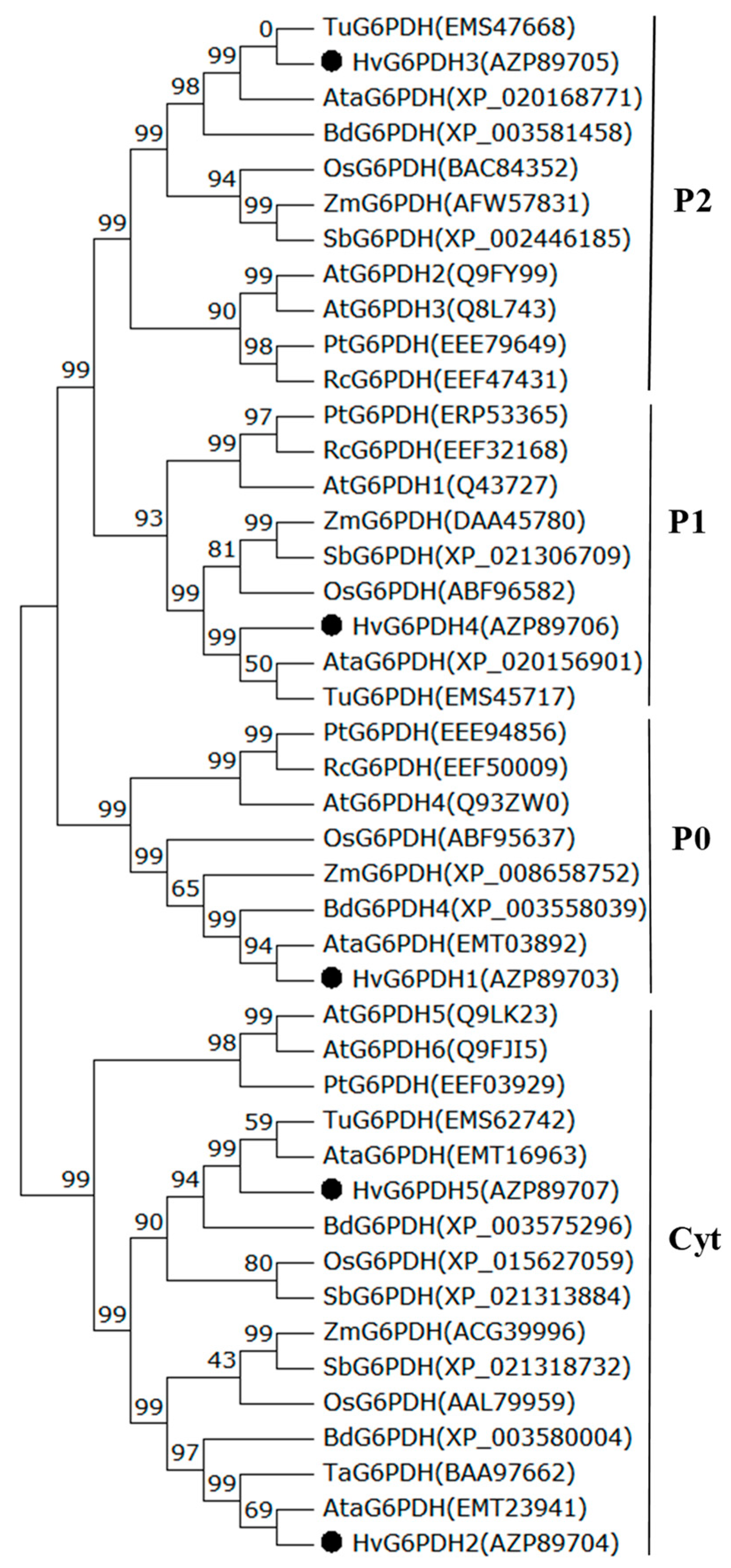
3.2. Phylogenetic Profiling and Subcellular Localization of HvG6PDH Proteins
3.3. NaCl and PEG Treatments Induce the G6PDH Activity in Highland Barley
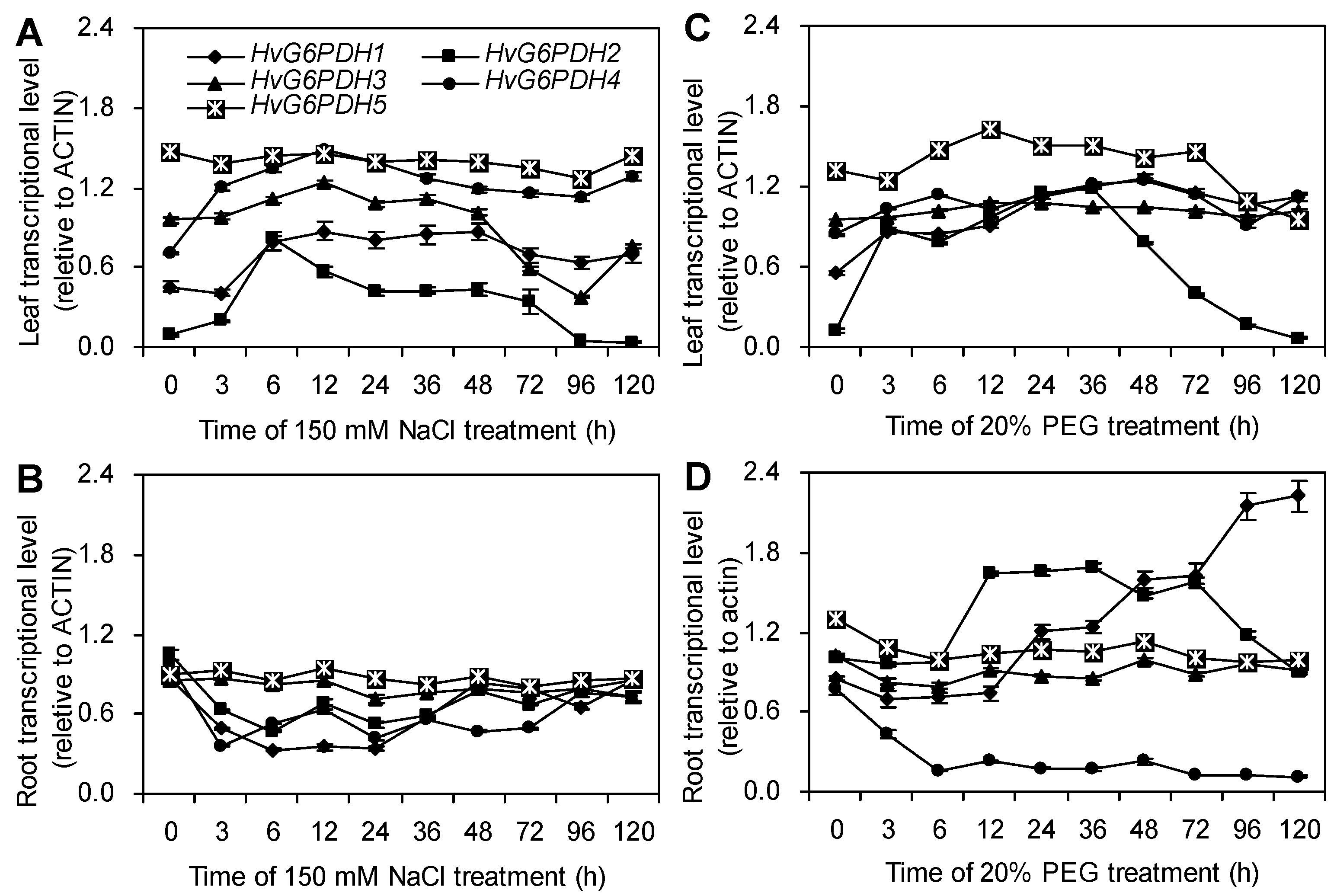
3.4. NaCl and PEG Treatments Activate the Expression of HvG6PDH Genes in Highland Barley
3.5. Abscisic Acid and Jasmonic Acid Positively Regulate the Activity and Expression of G6PDH
3.6. G6PDH Is Involved in ROS Scavenging under Salt and Drought Stresses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Boscaiu, M.; Vicente, O. Mechanisms of drought and salt stress tolerance in plants. J. Biotechnol. 2017, 256, S7. [Google Scholar] [CrossRef]
- Dal Santo, S.; Stampfl, H.; Krasensky, J.; Kempa, S.; Gibon, Y.; Petutschnig, E.; Rozhon, W.; Heuck, A.; Clausen, T.; Jonak, C. Stress-induced GSK3 regulates the redox stress response by phosphorylating glucose-6-phosphate dehydrogenase in Arabidopsis. Plant Cell 2012, 24, 3380–3392. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Tahi, H.; Wahbi, S.; Wakrim, R.; Aganchich, B.; Serraj, R.; Centritto, M. Water relations, photosynthesis, growth and water-use efficiency in tomato plants subjected to partial rootzone drying and regulated deficit irrigation. Plant Biosyst. 2007, 141, 265–274. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, L.; Li, Y.; Hou, J.; Huang, J.; Liang, W. Involvement of ABA- and H2O2-dependent cytosolic glucose-6-phosphate dehydrogenase in maintaining redox homeostasis in soybean roots under drought stress. Plant Physiol. Biochem. 2016, 107, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Moller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Von Schaewen, A.; Langenkamper, G.; Graeve, K.; Wenderoth, I.; Scheibe, R. Molecular characterization of the plastidic glucose-6-phosphate-dehydrogenase from potato in comparison to its cytosolic counterpart. Plant Physiol. 1995, 109, 1327–1335. [Google Scholar] [CrossRef] [Green Version]
- Thom, E.; Mohlmann, T.; Quick, W.P.; Camara, B.; Neuhaus, H.E. Sweet pepper plastids: Enzymic equipment, characterisation of the plastidic oxidative pentose-phosphate pathway, and transport of phosphorylated intermediates across the envelope membrane. Planta 1998, 204, 226–233. [Google Scholar] [CrossRef]
- Meyer, T.; Holscher, C.; Schwoppe, C.; von Schaewen, A. Alternative targeting of Arabidopsis plastidic glucose-6-phosphate dehydrogenase G6PD1 involves cysteine-dependent interaction with G6PD4 in the cytosol. Plant J. 2011, 66, 745–758. [Google Scholar] [CrossRef]
- Rao, X.J.; Duan, X.T.; Mao, W.M.; Li, X.X.; Li, Z.H.; Li, Q.; Zheng, Z.G.; Xu, H.M.; Chen, M.; Wang, P.G.; et al. O-GlcNAcylation of G6PD promotes the pentose phosphate pathway and tumor growth. Nat. Commun. 2015, 6, 8468. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Y.; Zheng, X.X.; Yang, J.; Messing, J.; Wu, Y.R. Maize endosperm-specific transcription factors O2 and PBF network the regulation of protein and starch synthesis. Proc. Natl. Acad. Sci. USA 2016, 113, 10842–10847. [Google Scholar] [CrossRef] [Green Version]
- Yuzbasheva, E.Y.; Mostova, E.B.; Andreeva, N.I.; Yuzbashev, T.V.; Laptev, I.A.; Sobolevskaya, T.I.; Sineoky, S.P. Co-expression of glucose-6-phosphate dehydrogenase and acyl-CoA binding protein enhances lipid accumulation in the yeast Yarrowia lipolytica. New Biotechnol. 2017, 39, 18–21. [Google Scholar] [CrossRef]
- Acero-Navarro, K.E.; Jimenez-Ramirez, M.; Villalobos, M.A.; Vargas-Martinez, R.; Perales-Vela, H.V.; Velasco-Garcia, R. Cloning, overexpression, and purification of glucose-6-phosphate dehydrogenase of Pseudomonas aeruginosa. Protein Expr. Purif. 2018, 142, 53–61. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Kruger, N.J.; von Schaewen, A. The oxidative pentose phosphate pathway: Structure and organisation. Curr. Opin. Plant Biol. 2003, 6, 236–246. [Google Scholar] [CrossRef]
- Wakao, S.; Benning, C. Genome-wide analysis of glucose-6-phosphate dehydrogenases in Arabidopsis. Plant J. 2005, 41, 243–256. [Google Scholar] [CrossRef]
- Sindelar, L.; Sindelarova, M. Correlation of viral RNA biosynthesis with glucose-6-phosphate dehydrogenase activity and host resistance. Planta 2002, 215, 862–869. [Google Scholar]
- Liu, J.; Wang, X.M.; Hu, Y.F.; Hu, W.; Bi, Y.R. Glucose-6-phosphate dehydrogenase plays a pivotal role in tolerance to drought stress in soybean roots. Plant Cell Rep. 2013, 32, 415–429. [Google Scholar] [CrossRef]
- Wang, X.M.; Ruan, M.J.; Wan, Q.; He, W.L.; Yang, L.; Liu, X.Y.; He, L.; Yan, L.L.; Bi, Y.R. Nitric oxide and hydrogen peroxide increase glucose-6-phosphate dehydrogenase activities and expression upon drought stress in soybean roots. Plant Cell Rep. 2019, 39, 69–73. [Google Scholar] [CrossRef]
- Valderrama, R.; Corpas, F.J.; Carreras, A.; Gomez-Rodriguez, M.V.; Chaki, M.; Pedrajas, J.R.; Fernandez-Ocana, A.; Del Rio, L.A.; Barroso, J.B. The dehydrogenase-mediated recycling of NADPH is a key antioxidant system against salt-induced oxidative stress in olive plants. Plant Cell Environ. 2006, 29, 1449–1459. [Google Scholar] [CrossRef]
- Wang, X.; Ma, Y.; Huang, C.; Wan, Q.; Li, N.; Bi, Y. Glucose-6-phosphate dehydrogenase plays a central role in modulating reduced glutathione levels in reed callus under salt stress. Planta 2008, 227, 611–623. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, J.; Wang, X.M.; Bi, Y.R. Glucose-6-phosphate dehydrogenase acts as a regulator of cell redox balance in rice suspension cells under salt stress. Plant Growth Regul. 2013, 69, 139–148. [Google Scholar] [CrossRef]
- Cardi, M.; Castiglia, D.; Ferrara, M.; Guerriero, G.; Chiurazzi, M.; Esposito, S. The effects of salt stress cause a diversion of basal metabolism in barley roots: Possible different roles for glucose-6-phosphate dehydrogenase isoforms. Plant Physiol. Biochem. 2015, 86, 44–54. [Google Scholar] [CrossRef]
- Yang, L.; Wang, X.M.; Chang, N.; Nan, W.B.; Wang, S.W.; Ruan, M.J.; Sun, L.L.; Li, S.F.; Bi, Y.R. Cytosolic glucose-6-phosphate dehydro-genase is involved in seed germination and root growth under salinity in Arabidopsis. Front. Plant Sci. 2019, 10, 182. [Google Scholar] [CrossRef] [Green Version]
- Slaski, J.J.; Zhang, G.C.; Basu, U.; Stephens, J.L.; Taylor, G.J. Aluminum resistance in wheat (Triticum aestivum) is associated with rapid, Al-induced changes in activities of glucose-6-phosphate dehydrogenase and 6-pbosphogluconate dehydrogenase in root apices. Physiologia. Plantarum. 1996, 98, 477–484. [Google Scholar] [CrossRef]
- Wang, H.H.; Hou, J.J.; Li, Y.; Zhang, Y.Y.; Huang, J.J.; Liang, W.H. Nitric oxide-mediated cytosolic glucose-6-phosphate dehydrogenase is involved in aluminum toxicity of soybean under high aluminum concentration. Plant Soil. 2017, 416, 39–52. [Google Scholar] [CrossRef]
- Gong, H.L.; Chen, G.S.; Li, F.J.; Wang, X.M.; Hu, Y.F.; Bi, Y.R. Involvement of G6PDH in regulation of heat stress tolerance in the calli from Przewalskia tangutica and Nicotiana tabacum. Biol. Plantarum 2012, 56, 422–430. [Google Scholar] [CrossRef]
- Lin, Y.Z.; Lin, S.Z.; Guo, H.; Zhang, Z.Y.; Chen, X.Y. Functional analysis of PsG6PDH, a cytosolic glucose-6-phosphate dehydrogenase gene from Populus suaveolens, and its contribution to cold tolerance improvement in tobacco plants. Biotechnol. Lett. 2013, 35, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.T.; Fu, Z.W.; Su, Y.C.; Zhang, X.; Li, G.Y.; Guo, J.L.; Que, Y.X.; Xu, L.P. A cytosolic glucose-6-phosphate dehydrogenase gene, ScG6PDH, plays a positive role in response to various abiotic stresses in sugarcane. Sci. Rep. 2014, 4, 7090. [Google Scholar] [CrossRef]
- He, L.; Wang, X.; Feng, R.; He, Q.; Wang, S.; Liang, C.; Yan, L.; Bi, Y. Alternative Pathway is Involved in Nitric Oxide-Enhanced Tolerance to Cadmium Stress in Barley Roots. Plants 2019, 8, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, S.; Guerriero, G.; Vona, V.; Di Martino Rigano, V.; Carfagna, S.; Rigano, C. Glutamate synthase activities and protein changes in relation to nitrogen nutrition in barley: The dependence on diferent plastidic glucose-6P dehydrogenase isoforms. J. Exp. Bot. 2005, 56, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Nyima, T.; Tang, Y.; Zeng, X. Food preparation from hulless barley in Tibet. Adv. Barley Sci. 2013, 151–158. [Google Scholar]
- Zeng, X.Q.; Long, H.; Wang, Z.; Zhao, S.C.; Tang, Y.W.; Huang, Z.Y.; Wang, Y.L.; Xu, Q.J.; Mao, L.K.; Deng, G.B.; et al. The draft genome of Tibetan hulless barley reveals adaptive patterns to the high stressful Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2015, 112, 1095–1100. [Google Scholar] [CrossRef] [Green Version]
- Dai, F.; Nevo, E.; Wu, D.Z.; Comadran, J.; Zhou, M.X.; Qiu, L.; Chen, Z.H.; Beiles, A.; Chen, G.X.; Zhang, G.P. Tibet is one of the centers of domestication of cultivated barley. Proc. Natl. Acad. Sci. USA 2012, 109, 16969–16973. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.Z.; Wang, X.M.; Wang, X.Y.; Wu, K.L.; Li, P.; Chang, N.; Wang, J.F.; Wang, F.; Li, J.L.; Bi, Y.R. Glucose-6-phosphate dehydrogenase and alternative oxidase are involved in the cross tolerance of highland barley to salt stress and UV-B radiation. J. Plant. Physiol. 2015, 181, 83–95. [Google Scholar] [CrossRef]
- Zeng, X.Q.; Bai, L.J.; Wei, Z.X.; Yuan, H.J.; Wang, Y.L.; Xu, Q.J.; Tang, Y.W.; Nyima, T. Transcriptome analysis revealed the drought-responsive genes in Tibetan hulless barley. BMC Genom. 2016, 17, 386. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.J.; Zeng, X.Q.; Ling, Z.H.; Wei, Z.X.; Wang, Y.L.; Zhuang, Z.H.; Xu, Q.J.; Tang, Y.W.; Tashi, N. Transcriptome profiles reveal cold acclimation and freezing tolerance of susceptible and tolerant hulless barley genotypes. Acta. Physiol. Plant. 2017, 39, 275. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, R.; Wan, Q.; Xie, G.; Bi, Y. Glucose-6-phosphate dehydrogenase plays a pivotal role in nitric oxide-involved defense against oxidative stress under salt stress in red kidney bean roots. Plant Cell Physiol. 2007, 48, 511–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.S.; Chen, G.C.; Wang, X.M.; Zhang, Y.L.; Jia, H.L.; Bi, Y.R. Glucose-6-phosphate dehydrogenase-dependent hydrogen peroxide production is involved in the regulation of plasma membrane H+-ATPase and Na+/H+ antiporter protein in salt-stressed callus from Carex moorcroftii. Physiol. Plant. 2011, 141, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCT method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wendt, U.K.; Wenderoth, I.; Tegeler, A.; von Schaewen, A. Molecular characterization of a novel glucose-6-phosphate dehydrogenase from potato (Solanum tuberosum L.). Plant J. 2000, 23, 723–733. [Google Scholar] [CrossRef]
- Hauschild, R.; von Schaewen, A. Differential regulation of glucose-6-phosphate dehydrogenase isoenzyme activities in potato. Plant Physiol. 2003, 133, 47–62. [Google Scholar] [CrossRef] [Green Version]
- Koide, H.; Oda, T. Pathological occurrence of glucose-6-phosphatase in serum in liver diseases. Clin. Chim. Acta 1959, 4, 554–561. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Veljovic-Jovanovic, S.; Noctor, G.; Foyer, C.H. Are leaf hydrogen peroxide concentrations commonly overestimated? The potential influence of artefactual interference by tissue phenolics and ascorbate. Plant Physiol. Biochem. 2002, 40, 501–507. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Ling, H.Q.; Zhao, S.C.; Liu, D.C.; Wang, J.Y.; Sun, H.; Zhang, C.; Fan, H.; Li, D.; Dong, L.L.; Tao, Y.; et al. Draft genome of the wheat A-genome progenitor Triticum urartu. Nature 2013, 496, 87–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenti, V.; Stanghellini, M.A.; Pupillo, P. Glucose-6-phosphate-dehydrogenase isozymes of maize leaves—some comparative properties. Plant Physiol. 1984, 5, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.C.; Gu, Y.Q.; Puiu, D.; Wang, H.; Twardziok, S.O.; Deal, K.R.; Huo, N.X.; Zhu, T.T.; Wang, L.; Wang, Y.; et al. Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature 2017, 551, 498–502. [Google Scholar] [CrossRef]
- Cardi, M.; Chibani, K.; Castiglia, D.; Cafasso, D.; Pizzo, E.; Rouhier, N.; Jacquot, J.P.; Esposito, S. Overexpression, purification and enzymatic characterization of a recombinant plastidial glucose-6-phosphate dehydrogenase from barley (Hordeum vulgare cv. Nure) roots. Plant Physiol. Biochem. 2013, 73, 266–273. [Google Scholar] [CrossRef] [Green Version]
- Long, X.Y.; He, B.; Fang, Y.J.; Tang, C.R. Identification and characterization of the glucose-6-phosphate dehydrogenase gene family in the para rubber tree, Hevea brasiliensis. Front Plant Sci. 2016, 7, 215. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Scharte, J.; Schon, H.; Tjaden, Z.; Weis, E.; von Schaewen, A. Isoenzyme replacement of glucose-6-phosphate dehydrogenase in the cytosol improves stress tolerance in plants. Proc. Natl. Acad. Sci. USA 2009, 106, 8061–8066. [Google Scholar] [CrossRef] [Green Version]
- Pandolfi, P.P.; Sonati, F.; Rivi, R.; Mason, P.; Grosveld, F.; Luzzatto, L. Targeted disruption of the housekeeping gene encoding glucose-6-phosphate-dehydrogenase (G6PD): G6PD is dispensable for pentose synthesis but essential for defense against oxidative stress. EMBO. J. 1995, 14, 5209–5215. [Google Scholar] [CrossRef]
- Juhnke, H.; Krems, B.; Kotter, P.; Entian, K.D. Mutants that show increased sensitivity to hydrogen peroxide reveal an important role for the pentose phosphate pathway in protection of yeast against oxidative stress. Mol. Gen. Genet. 1996, 252, 456–464. [Google Scholar] [CrossRef]
- Leopold, J.A.; Zhang, Y.Y.; Scribner, A.W.; Stanton, R.C.; Loscalzo, J. Glucose-6-phosphate dehydrogenase overexpression decreases endothelial cell oxidant stress and increases bioavailable nitric oxide. Arterioscl. Throm. Vas. 2003, 23, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Mendez, D.; Linares, M.; Diez, A.; Puyet, A.; Bautista, J.M. Stress response and cytoskeletal proteins involved in erythrocyte membrane remodeling upon Plasmodium falciparum invasion are differentially carbonylated in G6PD A(-) deficiency. Free Radical. Biol. Med. 2011, 50, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Honjoh, K.; Machida, T.; Hagisako, T.; Suga, K.; Yonekura, M.; Shimizu, H.; Ohashi, N.; Miyamoto, T.; Hatano, S.; Iio, M. Molecular cloning and characterization of a cDNA for low-temperature inducible cytosolic glucose 6-phosphate dehydrogenase gene from Chlorella vulgaris and expression of the gene in Saccharomyces cerevisiae. Plant Sci. 2007, 172, 649–658. [Google Scholar] [CrossRef]
- Savchenko, T.; Kolla, V.A.; Wang, C.Q.; Nasafi, Z.; Hicks, D.R.; Phadungchob, B.; Chehab, W.E.; Brandizzi, F.; Froehlich, J.; Dehesh, K. Functional convergence of oxylipin and abscisic acid pathways controls stomatal closure in response to drought. Plant Physiol. 2014, 164, 1151–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC. Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Wu, H.; Ma, S.Q.; Xiang, D.H.; Liu, R.Y.; Xiong, L.Z. OsJAZ1 attenuates drought resistance by regulating JA and ABA signaling in rice. Front. Plant Sci. 2017, 8, 2108. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Ye, H.Y.; Yao, R.F.; Zhang, T.; Xiong, L.Z. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling and modulates salt stress tolerance in rice. Plant Sci. 2015, 232, 1–12. [Google Scholar] [CrossRef]
- Cardi, M.; Chibani, K.; Cafasso, D.; Rouhier, N.; Jacquot, J.P.; Esposito, S. Abscisic acid effects on activity and expression of barley (Hordeum vulgare) plastidial glucose-6-phosphate dehydrogenase. J. Exp. Bot. 2011, 62, 4013–4023. [Google Scholar] [CrossRef] [Green Version]
- Dietz, K.J. Redox control, redox signaling, and redox homeostasis in plant cells. Int. Rev. Cytol. 2003, 228, 141–193. [Google Scholar]
- Bowsher, C.G.; Boulton, E.L.; Rose, J.K.C.; Nayagam, S.; Emes, M.J. Reductant for glutamate synthase is generated by the oxidative pentose-phosphate pathway in non-photosynthetic root plastids. Plant J. 1992, 2, 893–898. [Google Scholar] [CrossRef]
- Filosa, S.; Fico, A.; Paglialunga, F.; Balestrieri, M.; Crooke, A.; Verde, P.; Abrescia, P.; Bautista, J.M.; Martini, G. Failure to increase glucose consumption through the pentose-phosphate pathway results in the death of glucose-6-phosphate dehydrogenase gene-deleted mouse embryonic stem cells subjected to oxidative stress. Biochem. J. 2003, 370, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Izawa, S.; Maeda, K.; Miki, T.; Mano, J.; Inoue, Y.; Kimura, A. Importance of glucose-6-phosphate dehydrogenase in the adaptive response to hydrogen peroxide in Saccharomyces cerevisiae. Biochem. J. 1998, 330, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Chromosome Location | Predicted Localization | aa | TP | pI | MW (kD) | GenBank |
|---|---|---|---|---|---|---|---|
| HvG6PDH1 | 4H:433572159-433576826 | Chloroplast | 596 | 27 | 5.4 | 67.0 | MK034128 |
| HvG6PDH2 | 2H:608625462-608631046 | Cytosol | 509 | − | 6.3 | 57.9 | MK034129 |
| HvG6PDH3 | 2H:20076557-20081131 | Chloroplast | 589 | 39 | 8.3 | 65.6 | MK034130 |
| HvG6PDH4 | 6H:114688120-114690749 | Chloroplast | 574 | 37 | 8.2 | 64.8 | MK034131 |
| HvG6PDH5 | 6H:342276577-342281388 | Cytosol | 516 | − | 5.9 | 58.3 | MK034132 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, R.; Wang, X.; He, L.; Wang, S.; Li, J.; Jin, J.; Bi, Y. Identification, Characterization, and Stress Responsiveness of Glucose-6-phosphate Dehydrogenase Genes in Highland Barley. Plants 2020, 9, 1800. https://doi.org/10.3390/plants9121800
Feng R, Wang X, He L, Wang S, Li J, Jin J, Bi Y. Identification, Characterization, and Stress Responsiveness of Glucose-6-phosphate Dehydrogenase Genes in Highland Barley. Plants. 2020; 9(12):1800. https://doi.org/10.3390/plants9121800
Chicago/Turabian StyleFeng, Ruijun, Xiaomin Wang, Li He, Shengwang Wang, Junjie Li, Jie Jin, and Yurong Bi. 2020. "Identification, Characterization, and Stress Responsiveness of Glucose-6-phosphate Dehydrogenase Genes in Highland Barley" Plants 9, no. 12: 1800. https://doi.org/10.3390/plants9121800