Rootstock-Scion Interaction Affects the Composition and Pathogen Inhibitory Activity of Tobacco (Nicotiana tabacum L.) Root Exudates

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Test Plant Materials
2.2. Preparation of Inoculum of Phytophthora nicotianae
2.3. Disease Resistance Evaluation
2.4. Root Exudates Collection
2.5. Effects of Root Exudates on Phytophthora Nicotianae
2.6. Gas Chromatography-Mass Spectrometer (GC-MS) Analysis of Root Exudates
2.7. Identification of Root Exudates Composition
2.8. Effects of Compounds on Phytophthora nicotianae Mycelia Growth
2.9. Data Statistics
3. Results
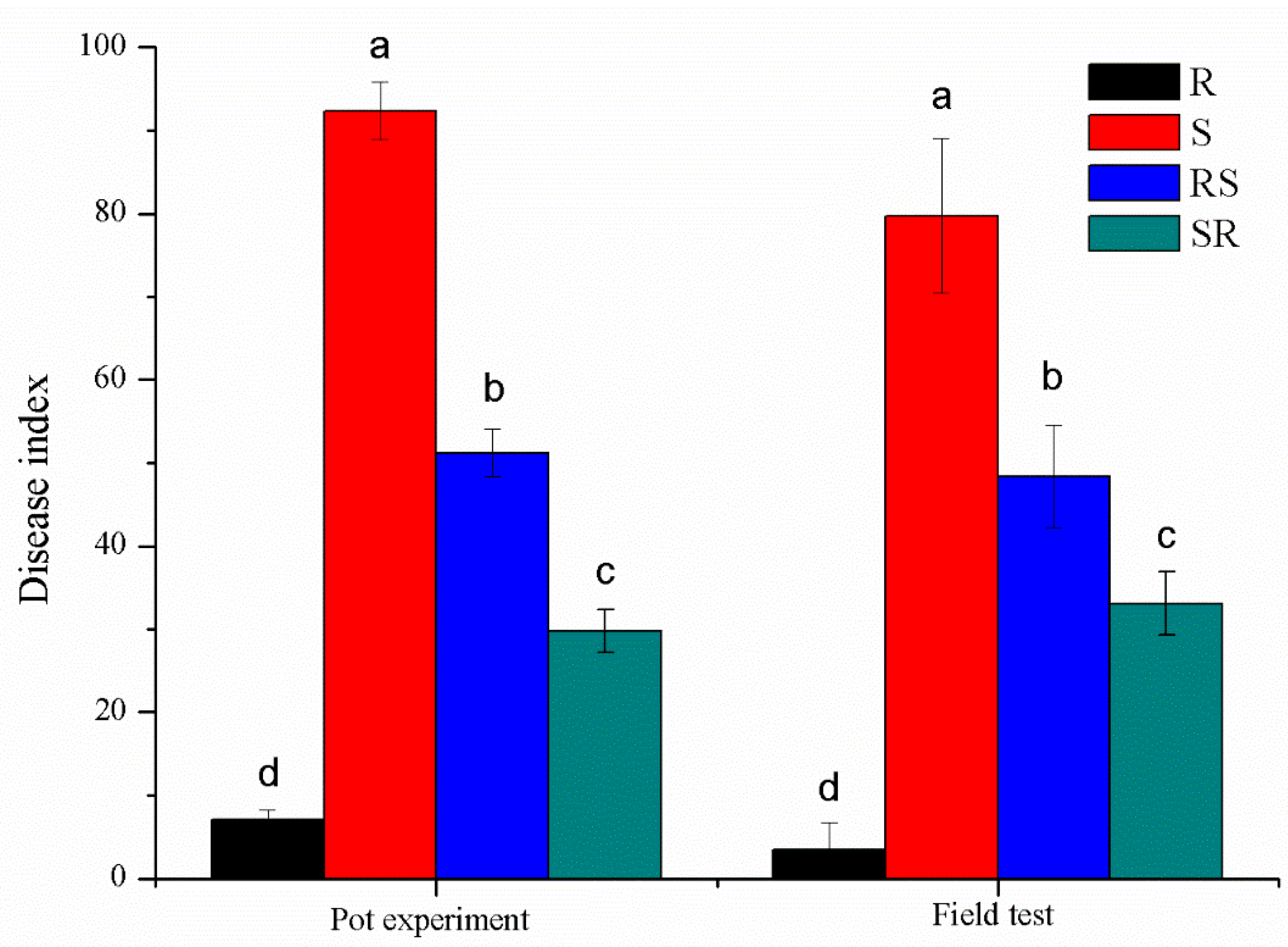
3.1. Variety Resistance Evaluation for Tobacco Black Shank
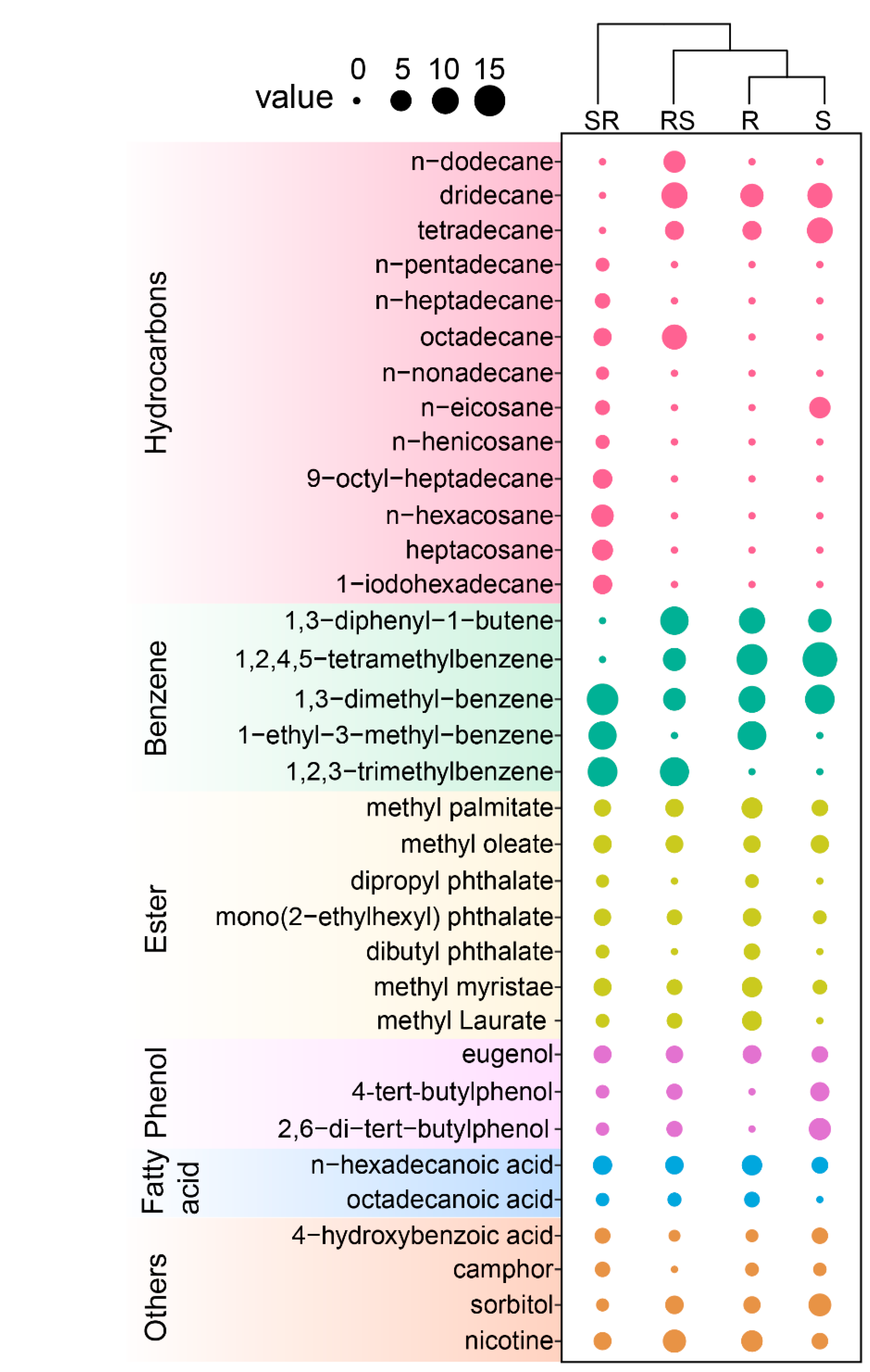
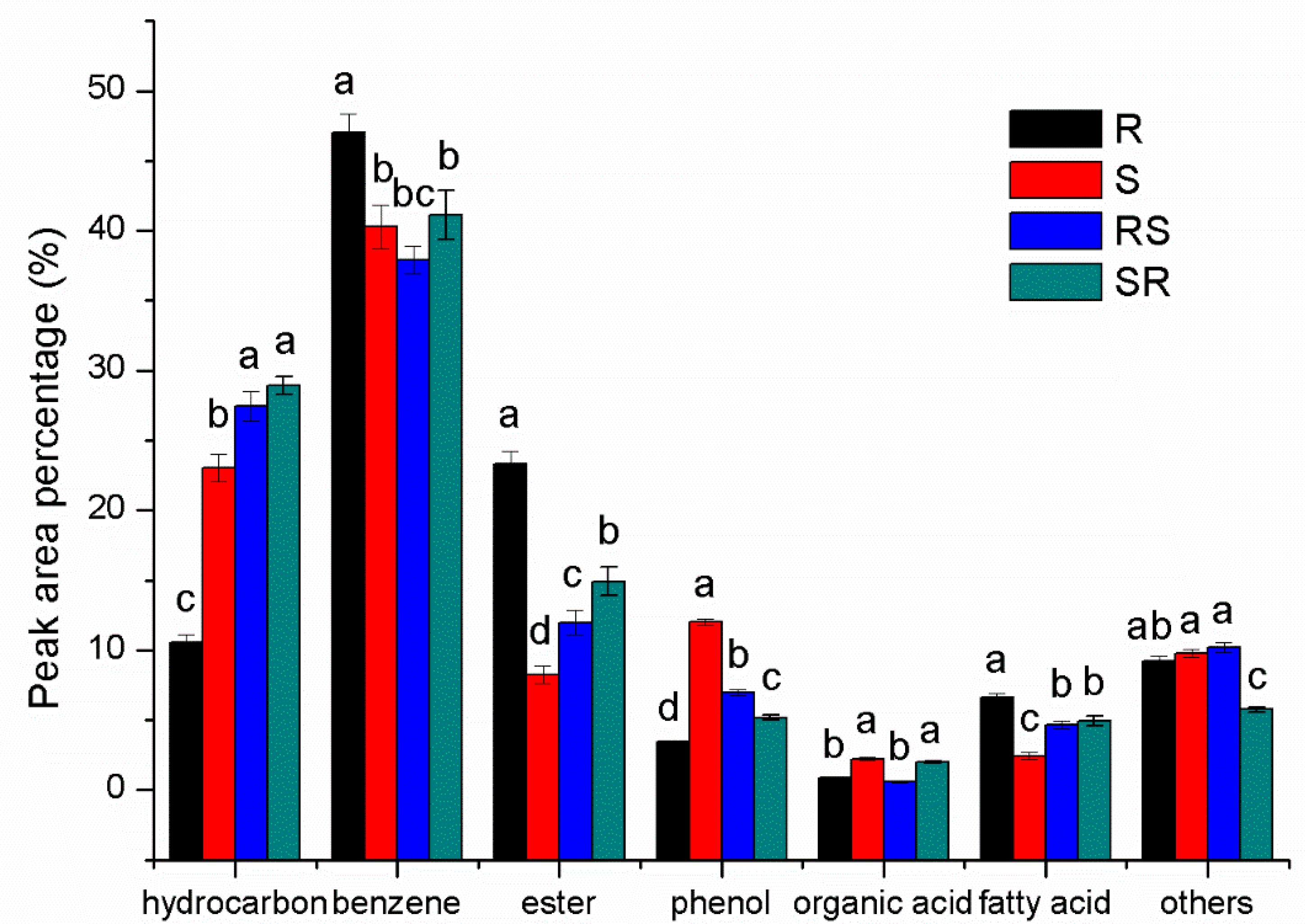
3.2. GC-MS Analysis of Root Exudates
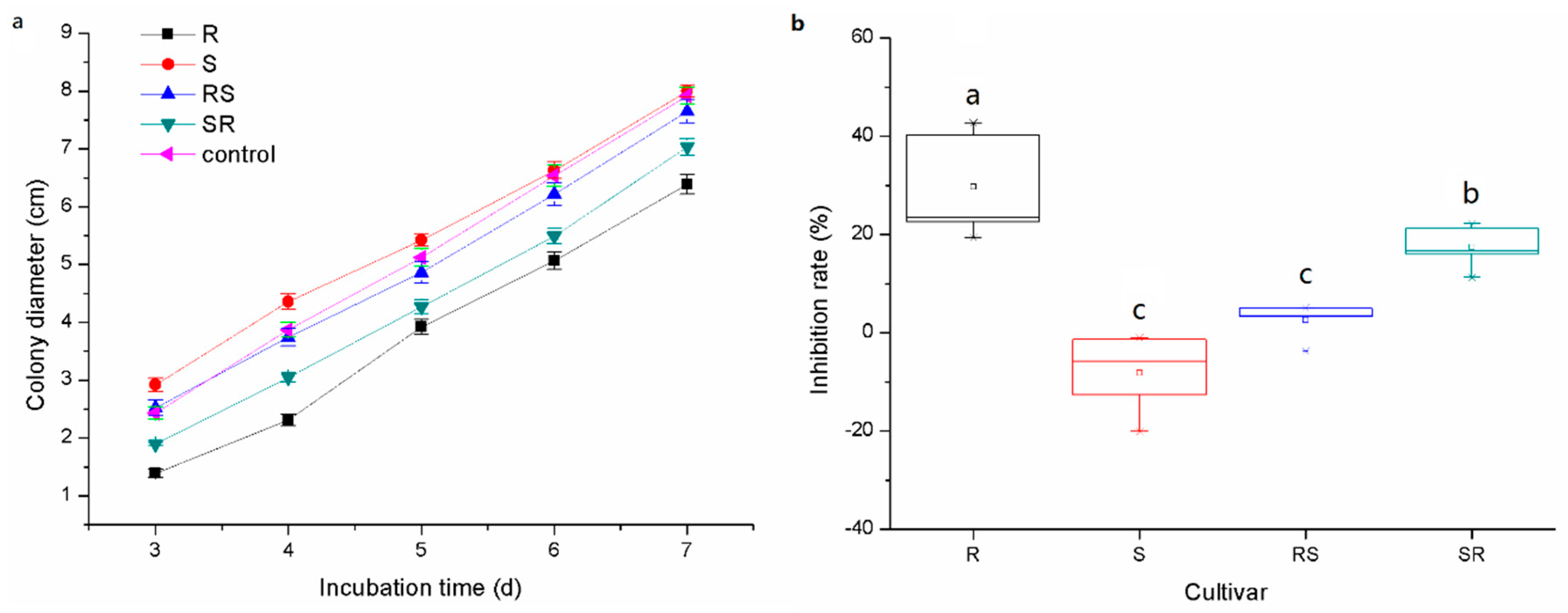
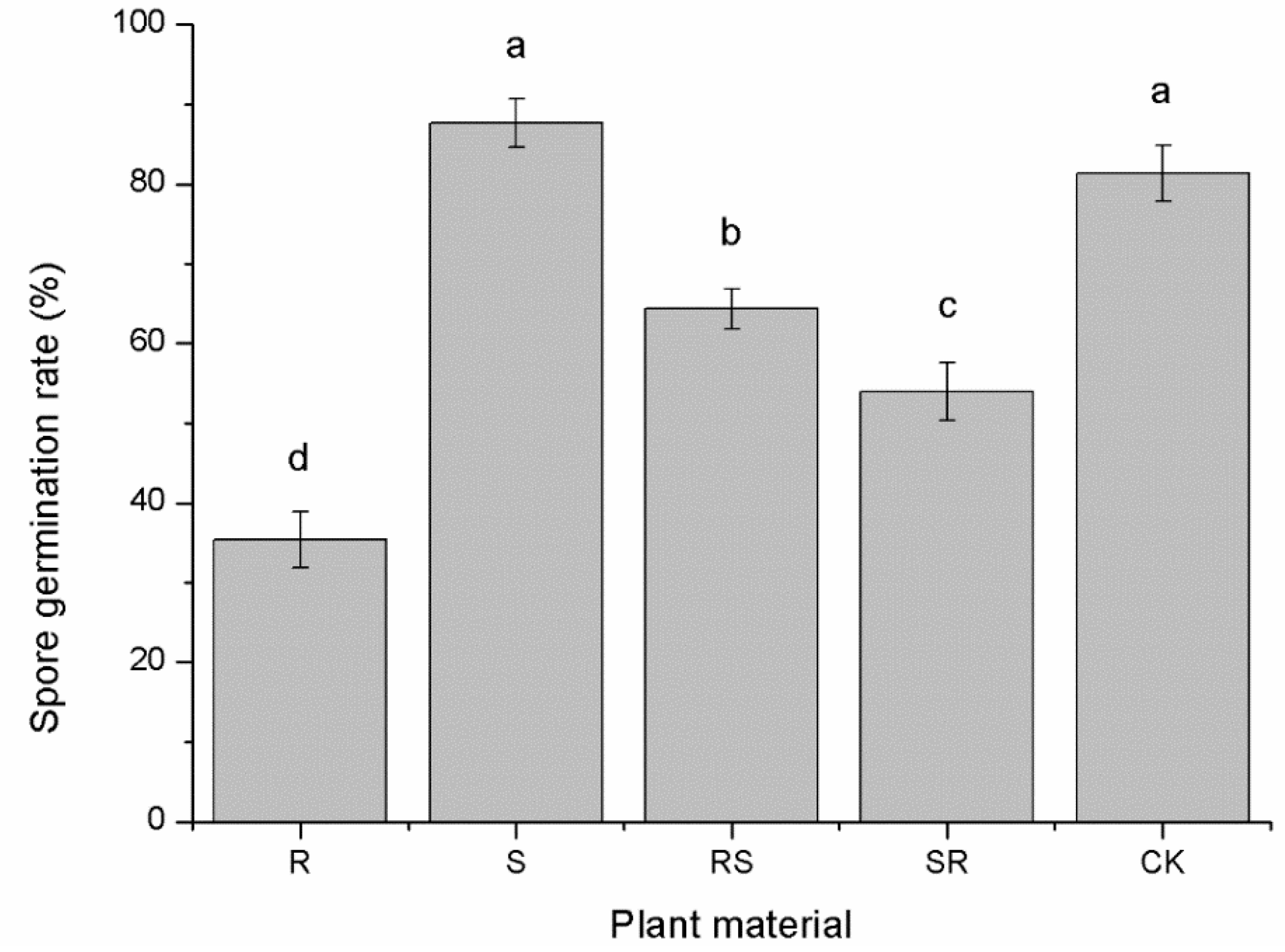
3.3. Allelopathy of Root Exudates on Phytophthora nicotianae
3.4. Allelopathy of Compounds on Phytophthora nicotianae Mycelium Growth
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Panabieres, F.; Ali, G.S.; Allagui, M.B.; Dalio, R.J.D.; Gudmestad, N.C.; Kuhn, M.; Roy, S.G.; Schena, L.; Zampounis, A. Phytophthora nicotianae diseases worldwide: New knowledge of a long-recognised pathogen. Phytopatho. Mediterr. 2016, 55, 20–40. [Google Scholar]
- Wang, Y.; Wei, K.; Han, X.; Zhao, D.; Zheng, Y.; Chao, J.; Gou, J.; Kong, F.; Zhang, C.S. The Antifungal Effect of Garlic Essential Oil on Phytophthora nicotianae and the Inhibitory Component Involved. Biomolecules 2019, 9, 632. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.C.; Chen, X.J.; Cai, L.T.; Cao, Y.; Lu, N.; Xia, H.Q.; Wang, M.S.; Shang, S.H. Race distribution and distribution of sensitivities to mefenoxam among isolates of Phytophthora parasitica var. nicotianae in Guizhou province of China. Crop Prot. 2013, 52, 136–140. [Google Scholar]
- Gilardi, G.; Gullino, M.L.; Garibaldi, A. Critical aspects of grafting as a possible strategy to manage soil-borne pathogens. Sci. Hortic. 2013, 149, 19–21. [Google Scholar] [CrossRef] [Green Version]
- Abebe, A.M.; Wai, K.P.P.; Siddique, M.I.; Mo, H.; Yoo, J.Y.; Jegal, Y.; Byeon, S.; Jang, K.; Jeon, S.; Hwang, J.; et al. Evaluation of Phytophthora root rot- and bacterial wilt-resistant inbred lines and their crosses for use as rootstocks in pepper (Capsicum annuum L.). Hortic. Environ. Biotechnol. 2016, 57, 598–605. [Google Scholar] [CrossRef]
- Haegi, A.; De Felice, S.; Scotton, M.; Luongo, L.; Belisario, A. Fusarium oxysporum f.sp. melonis-melon interaction: Effect of grafting combination on pathogen gene expression. Eur. J. Plant. Pathol. 2017, 149, 787–796. [Google Scholar] [CrossRef]
- Mackey, M.; Kurosky, A.; Nazar, R.N. A Graft Mimic Strategy for Verticillium Resistance in Tomato. Mol. Biotechnol. 2018, 60, 665–669. [Google Scholar] [CrossRef]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J.H. Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, water stress and organic pollutants. Sci. Hortic. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- Wang, H.; Shi, J.; Luo, Z.; Chen, Y.; Zhang, Y.; Li, Z. Resistant identification of tobacco grafted seedlings to black shank. Plant Prot. 2018, 44, 168–171, 224. [Google Scholar]
- Liu, J.; Zhu, B.; Hu, S.; Wang, C.; Yang, T. Effects of grafting on growth, development and black shank resistance of flue-cured tobacco. Tob. Sci. Technol. 2018, 51, 8–16. [Google Scholar]
- Wu, F.; An, M. Effects of Watermelon Cultivars with Different Resistances to Fusarium oxysporum f.sp. niveum and Grafting on Rhizosphere Soil Microorganism Population and Community Structure. Chin. Agric. Sci. 2011, 44, 4636–4644. [Google Scholar]
- Guan, W.; Zhao, X.; Hassell, R.; Thies, J.A. Defense mechanisms involved in disease resistance of grafted vegetables. HortScience 2012, 47, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Bi, H.G.T.; Li, G.X.; Wu, Q.; Li, M.; Ai, X.Z. Root characteristics of grafted peppers and their resistance to Fusarium solani. Biol. Plant. 2017, 61, 579–586. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Van Der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Baetz, U.; Martinoia, E. Root exudates: The hidden part of plant defense. Trends Plant Sci. 2014, 19, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere interactions: Root exudates, microbes, and microbial communities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- De-la-Pena, C.; Badri, D.V.; Lei, Z.; Watson, B.S.; Brandao, M.M.; Silva-Filho, M.C.; Sumner, L.W.; Vivanco, J.M. Root secretion of defense-related proteins is development-dependent and correlated with flowering time. J. Biol. Chem. 2010, 285, 30654–30665. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Chen, Z.; Du, L.; Xie, Y.; Ye, X. Allelopathy of the root exudates from different resistant eggplants to verticillium wilt (Verticillium dahliae Kleb). Acta Ecol. Sin. 2011, 31, 3964–3972. [Google Scholar]
- Ling, N.; Zhang, W.; Wang, D.; Mao, J.; Huang, Q.; Guo, S.; Shen, D. Root Exudates from Grafted-Root Watermelon Showed a Certain Contribution in Inhibiting Fusarium oxysporum f. sp. niveum. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Zhang, C.; Gao, J.; Han, T.; Tian, X.; Wang, F. Integrated control of tobacco black shank by combined use of riboflavin and Bacillus subtilis strain Tpb55. BioControl 2017, 62, 835–845. [Google Scholar] [CrossRef]
- Lee, J.M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Zhang, C.; Feng, C.; Zheng, Y.; Wang, J.; Wang, F. Root Exudates Metabolic Profiling Suggests Distinct Defense Mechanisms Between Resistant and Susceptible Tobacco Cultivars Against Black Shank Disease. Front. Plant Sci. 2020, 11, 559775. [Google Scholar] [CrossRef] [PubMed]
- Badri, D.V.; De-La-Peña, C.; Lei, Z.; Manter, D.K.; Chaparro, J.M.; Guimarães, R.L.; Sumner, L.W.; Vivanco, J.M. Root Secreted Metabolites and Proteins Are Involved in the Early Events of Plant-Plant Recognition Prior to Competition. PLoS ONE 2012, 7, e46640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, R.A. Natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef]
- Bertin, C.; Yang, X.; Weston, L. The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 2003, 256, 67–83. [Google Scholar] [CrossRef]
- Hao, W.Y.; Ren, L.X.; Ran, W.; Shen, Q.R. Allelopathic effects of root exudates from watermelon and rice plants on Fusarium oxysporum f.sp. niveum. Plant Soil 2010, 336, 485–497. [Google Scholar] [CrossRef]
- Whalley, W.M.; Taylor, G.S. Influence of pea-root exudates on germination of conidia and chlamydospores of physiologic races of Fusarium oxysporum f. pisi. Ann. Appl. Biol. 1973, 73, 269–276. [Google Scholar] [CrossRef]
- Steinkellner, S.; Mammerler, R.; Vierheilig, H. Germination of Fusarium oxysporum in root exudates from tomato plants challenged with different Fusarium oxysporum strains. Eur. J. Plant Pathol. 2008, 122, 395. [Google Scholar] [CrossRef]
- Naqvi, S.M.A.; Chauhan, S.K. Effect of root exudates on the spore germiantion of rhizosphere and rhizoplane mycoflora of chilli (Capsicum annuum L.) cultivars. Plant Soil 1980, 55, 397–402. [Google Scholar] [CrossRef]
- Stevenson, P.C.; Padgham, D.E.; Haware, M.P. Root exudates associated with the resistance of four chickpea cultivars (Cicer arietinum) to two races of Fusarium oxysporum f.sp. cicero. Plant Pathol. 1995, 44, 686–694. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Liu, D.; Ling, N.; Bao, W.; Ying, R.; Shen, Q. Influence of Root Exudates of Watermelon on Fusarium oxysporum f. sp. niveum. Soil Sci. Soc. Am. J. 2008, 73, 1150–1156. [Google Scholar] [CrossRef]
- Schalchli, H.; Pardo, F.; Hormazabal, E.; Palma, R.; Guerrero, J.; Bensch, E. Antifungal activity of wheat root exudate extracts on Gaeumannomyces graminis var. tritici growth. J. Soil Sci. Plant Nutr. 2012, 12, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Dong, K.; Zheng, Y.; Yang, Z.; Tang, L.; Xiao, J. Allelopathic effects and components analysis of root exudates of faba bean cultivars with different degrees of resistance to Fusarium oxysporum. Chin. J. Eco-Agric. 2014, 22, 292–299. [Google Scholar] [CrossRef]
- Wang, J.; Lyu, Y.; Yu, D.; Zhang, W.; Piao, F.; Shen, S. Effects of Root Exudates from Different Resistant Pepper Varieties on Phytophthora capsici. China Veget. 2014, 1, 13–16. [Google Scholar] [CrossRef]
- Ren, Z.; Gai, Q. Root Exudates of Resistant and Susceptible Cotton Cultivars Sand lts Effects on Fusarium oxysporum f. sp. vasinfectum. Acta Agric. Boreali-Occident. Sin. 2016, 25, 702–706. [Google Scholar]
- Li, X.; Liu, B.; Heia, S.; Liu, D.; Han, Z.; Zhou, K.; Cui, J.; Luo, J.; Zheng, Y. The effect of root exudates from two transgenic insect-resistant cotton lines on the growth of Fusarium oxysporum. Transgenic Res. 2009, 18, 757–767. [Google Scholar] [CrossRef]
- Zhao, W.; Zheng, X.; Zhang, Y.; Zhong, C.; Yang, Y.; Yu, W. Effects of Root Exudates from Tomato Grafted with Different Rootstocks on Ralstonia solanacearum and Seedling Growth. China Veget. 2019, 5, 58–63. [Google Scholar]
- Weston, L.A.; Ryan, P.R.; Watt, M. Mechanisms for cellular transport and release of allelochemicals from plant roots into the rhizosphere. J. Exp. Bot. 2012, 63, 3445–3454. [Google Scholar] [CrossRef]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Yu, H.; Shen, G.; Gao, X. Determination of tobacco root exudates by GC-MS. Acta Tab. Sin. 2013, 19, 64–72. [Google Scholar]
- Deng, J.; Zhang, S.; Zhang, F.; Zhang, Y.; Hu, F.; Li, H. Autotoxins exuded from roots and the effects of PAEs on antioxidant capacity in roots of tobacco seedlings. Acta Ecol. Sin. 2017, 37, 495–504. [Google Scholar]
- Yang, R.X.; Gao, Z.G.; Liu, X.; Yao, Y.; Cheng, Y. Root exudates from muskmelon (Cucumis melon. L) induce autotoxicity and promote growth of Fusarium oxysporum f. sp. Melonis. Allelopath. J. 2014, 33, 175–188. [Google Scholar]
- Tian, G.; Bi, Y.; Sun, Z.; Zhang, L. Phenolic acids in the plow layer soil of strawberry fields and their effects on the occurrence of strawberry anthracnose. Eur. J. Plant Pathol. 2015, 581–594. [Google Scholar] [CrossRef]
- Tereshina, V.M.; Memorskaya, A.S.; Morozova, E.V.; Feofilova, E.P. Alterations in the carbohydrate composition of the cytosol of fungal spores caused by temperature variations and the storage process. Microbiology 2000, 69, 422–427. [Google Scholar] [CrossRef]
- Feofilova, E.P.; Ivashechkin, A.A.; Alekhin, A.I.; Sergeeva, Y.E. Fungal spores: Dormancy, germination, chemical composition, and role in biotechnology (review). Appl. Biochem. Microbiol. 2012, 48, 1–11. [Google Scholar] [CrossRef]
- Pfyffer, G.E.; Boraschi Gaia, C.; Weber, B.; Hoesch, L.; Orpin, C.G.; Rast, D.M. A further report on the occurrence of acyclic sugar alcohols in fungi. Mycol. Res. 1990, 94, 219–222. [Google Scholar] [CrossRef]
- Campbell, A.M.; Moon, R.P.; Duncan, J.M.; Gurr, S.J.; Kinghorn, J.R. Protoplast formation and regeneration from sporangia and encysted zoospores of Phytophthora infestans. Physiol. Mol. Plant Pathol. 1989, 34, 299–307. [Google Scholar] [CrossRef]
- Morris, E.P.F.; Ward, W.B. Chemoattraction of zoospores of the soybean pathogen, Phytophthora sojae, by isoflavones. Physiol. Mol. Plant Pathol. 1992, 40, 17–22. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, Y.; Song, G.; Gao, X.; Zhao, Y.; Jia, M.; Chen, Y.; Suo, B.; Chen, Q.; Wu, D.; et al. Phytophthora sojae zoospores differ in chemotaxis to the root and root exudates of host soybean and nonhost common bean. J. Gen. Plant Pathol. 2019, 85, 201–210. [Google Scholar] [CrossRef]
- Hsu, S.T.; Huang, H.C. Chemotaxes of Erwinia carotovora Subsp. carotovora and Erwinia chrysanthemi. Curr. Plant Sci. Biotechnol. Agric. 1987, 256. [Google Scholar] [CrossRef]
- Hu, X.; Xie, L.; Yu, C.; Li, Y.; Liu, S.; Zhang, C.; Liao, X. Chemotaxis of Bacillus megaterium strain A6 towards organic acid and saccharide from roots exudates of rapeseed. Chin. J. Oil Crop Sci. 2011, 33, 416–419. [Google Scholar]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; Smith, S.E.; Harvey, P.R.; Ryan, M.H.; Veneklaas, J.E.; Lambers, H.; et al. Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 2011, 4, 59–61. [Google Scholar] [CrossRef]
- Yuan, J.; Wu, Y.; Zhao, M.; Wen, T.; Huang, Q.; Shen, Q. Effect of phenolic acids from banana root exudates on root colonization and pathogen suppressive properties of Bacillus amyloliquefaciens NJN-6. Biol. Control 2018, 125, 131–137. [Google Scholar] [CrossRef]
- Li, X.; Ding, C.; Hua, K.; Zhang, T.; Zhang, Y.; Zhao, L.; Yang, Y.; Liu, J.; Wang, X. Soil sickness of peanuts is attributable to modifications in soil microbes induced by peanut root exudates rather than to direct allelopathy. Soil Biol. Biochem. 2014, 78, 149–159. [Google Scholar] [CrossRef]
- Zhang, K.; Xu, T.; Shen, F.; Shi, B.; Gu, M.; Shou, A.; Li, X. Phenolic Acids in Nicotiana tobacco L. Root Exudate and Their Autotoxicity Effects. Southwest China J. Agric. Sci. 2013, 26, 2552–2557. [Google Scholar]
- Yu, H.; Song, X.; Wang, S.; Cao, L.; Guo, L.; Wang, X.; Peng, G. Effects of Low Molecular Weight Organic Acids on Soil Enzymes Activities and Bacterial Community Structure. Chin. Agric. Sci. 2015, 48, 4936–4947. [Google Scholar]
- Huang, W. Study on Synergetic Effect of 2,4-DTBP and Fusarium on the Occurrence of Fusarium Wilt in Lanzhou Lily. J. Gansu Agric. Univ. 2018, 12, 35. [Google Scholar]
- Zhao, C.; Zhou, H.; Chai, Q.; Huang, G.; Liu, H.; Zhu, J. Effects of eugenol and intercropped faba-bean on wheat root growth under different water supply conditions. Acta Pratacult. Sin. 2014, 23, 133–139. [Google Scholar]
- Liu, P.; Zhao, H.; Tang, Z.; Zhang, Y.; Lin, H.; Shen, Y.; Wsng, J.; Wan, S. Effects of continuous cropping on root exudates of different resistance peanut (Arachis hypogaea L.) varieties and allelochemicals content in soil. Chin. J. Oil Crop Sci. 2015, 37, 467–474. [Google Scholar]
- Sun, M.; Ren, X.; Yao, H.; Cai, W.; Li, M. Allelopathic effect of 2, 6-Di-tert-butylphenol on pepper and its alleviating substances. Jiangsu Agric. Sci. 2017, 45, 124–126. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Diameter (cm) | Inhibition Rate (%) | ||||
|---|---|---|---|---|---|---|
| 5 μg/mL | 50 μg/mL | 500 μg/mL | 5 μg/mL | 50 μg/mL | 500 μg/mL | |
| p-tert-butylphenol | 3.68 ± 0.20 e | 0.22 ± 0.03 g | 0 i | 13.00 a | 94.82 a | 100.00 a |
| 4-hydroxybenzoic acid | 4.25 ± 0.09 b | 3.45 ± 0.05 d | 0 i | −0.47 d | 18.82 e | 100.00 a |
| mono (2-ethylhexyl) phthalate | 3.65 ± 0.05 e | 1.78 ± 0.13 f | 0 i | 13.71a | 58.12 b | 100.00 a |
| lauric acid | 4.28 ± 0.23 b | 2.53 ± 0.08 e | 0.67 ± 0.06 h | −1.18 d | 40.47 d | 83.78 b |
| 2,6-bis (1,1-dimethylethyl) phenol | 3.92 ± 0.19 d | 3.92 ± 0.20 c | 1.15 ± 0.05 g | 7.33 b | 7.76 h | 72.15 c |
| 2-benzenedicarboxylic acid dipropyl ester | 4.32 ± 0.10 ab | 3.75 ± 0.17 cd | 1.42 ± 0.08 f | −2.13 d | 11.76 f | 65.62 d |
| tetradecanoic acid | 4.38 ± 0.03 a | 4.05 ± 0.05 bc | 1.88 ± 0.13 e | −3.55 d | 4.71 i | 54.48 e |
| dibutyl phthalate | 4.32 ± 0.24 a | 4.15 ± 0.25 b | 3.88 ± 0.03 c | −2.13 d | 2.35 j | 6.05 h |
| methyl palmitate | 4.45 ± 0.13 a | 4.42 ± 0.20 a | 4.3 ± 0.40 ab | −5.20 e | −4.00 k | −4.12 j |
| nicotine | 4.18 ± 0.12 bc | 4.98 ± 0.03 a | 4.42 ± 0.10 a | 1.18 d | −17.18 l | −7.02 k |
| eugenol | 4.05 ± 0.05 c | 2.07 ± 0.03 f | 0 i | 4.26 c | 51.29 c | 100.00 a |
| sorbitol | 4.26 ± 0.11 b | 4.37 ± 0.10 a | 4.05 ± 0.08 b | −0.71 d | −2.82 k | 1.94 i |
| octadecanoic acid | 4.31 ± 0.0.06 a | 3.96 ± 0.07 c | 3.34 ± 0.05 d | −1.89 | 6.82 h | 19.13 g |
| n- hexadecanoic acid | 4.28 ± 0.10 b | 3.85 ± 0.09 c | 2.92 ± 0.10 d | −1.18 | 9.41 g | 29.30 f |
| DMSO (control) | 4.23 ± 0.06 bc | 4.25 ± 0.09 b | 4.13 ± 0.15 b | - | - | - |
| Compound | Mycelium Diameter (cm) | ||||
|---|---|---|---|---|---|
| 6.25 mM | 12.5 mM | 25 mM | 50 mM | 100 mM | |
| glucose | 5.28 ± 0.16 b | 5.20 ± 0.20 b | 5.68 ± 0.16 a | 5.52 ± 0.23 b | 5.37 ± 0.23 a |
| sorbitol | 5.90 ± 0.23 a | 5.80 ± 0.18 a | 5.62 ± 0.08 a | 5.90 ± 0.05 a | 5.63 ± 0.21 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.-S.; Zheng, Y.; Peng, L.; Cao, J. Rootstock-Scion Interaction Affects the Composition and Pathogen Inhibitory Activity of Tobacco (Nicotiana tabacum L.) Root Exudates. Plants 2020, 9, 1652. https://doi.org/10.3390/plants9121652
Zhang C-S, Zheng Y, Peng L, Cao J. Rootstock-Scion Interaction Affects the Composition and Pathogen Inhibitory Activity of Tobacco (Nicotiana tabacum L.) Root Exudates. Plants. 2020; 9(12):1652. https://doi.org/10.3390/plants9121652
Chicago/Turabian StyleZhang, Cheng-Sheng, Yanfen Zheng, Lijuan Peng, and Jianmin Cao. 2020. "Rootstock-Scion Interaction Affects the Composition and Pathogen Inhibitory Activity of Tobacco (Nicotiana tabacum L.) Root Exudates" Plants 9, no. 12: 1652. https://doi.org/10.3390/plants9121652