Effect of Salinity Stress on Growth and Metabolomic Profiling of Cucumis sativus and Solanum lycopersicum

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
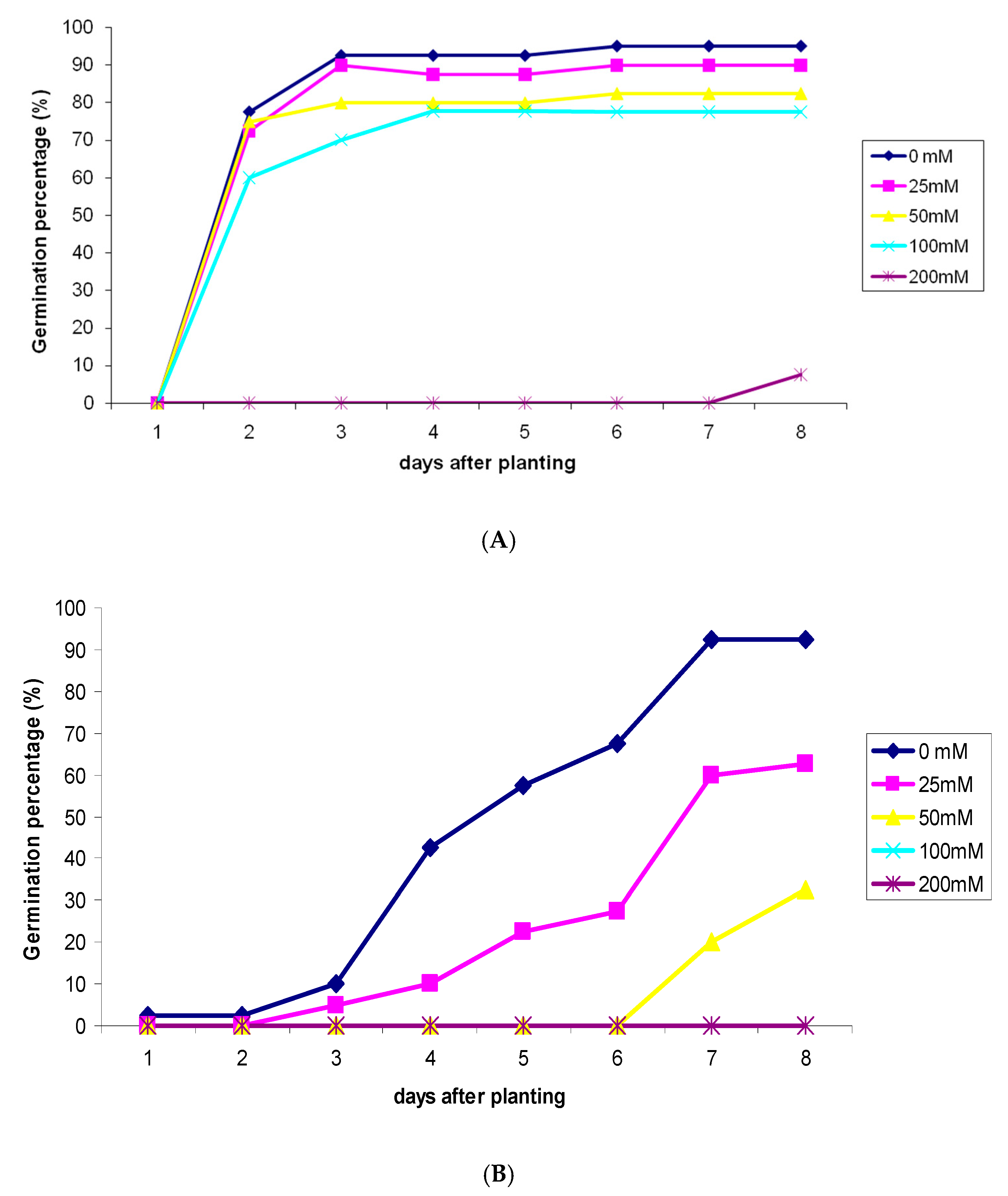
2.1. Effect of Salinity Stress on Seeds Germination Rate of C. sativus and S. lycopersicum
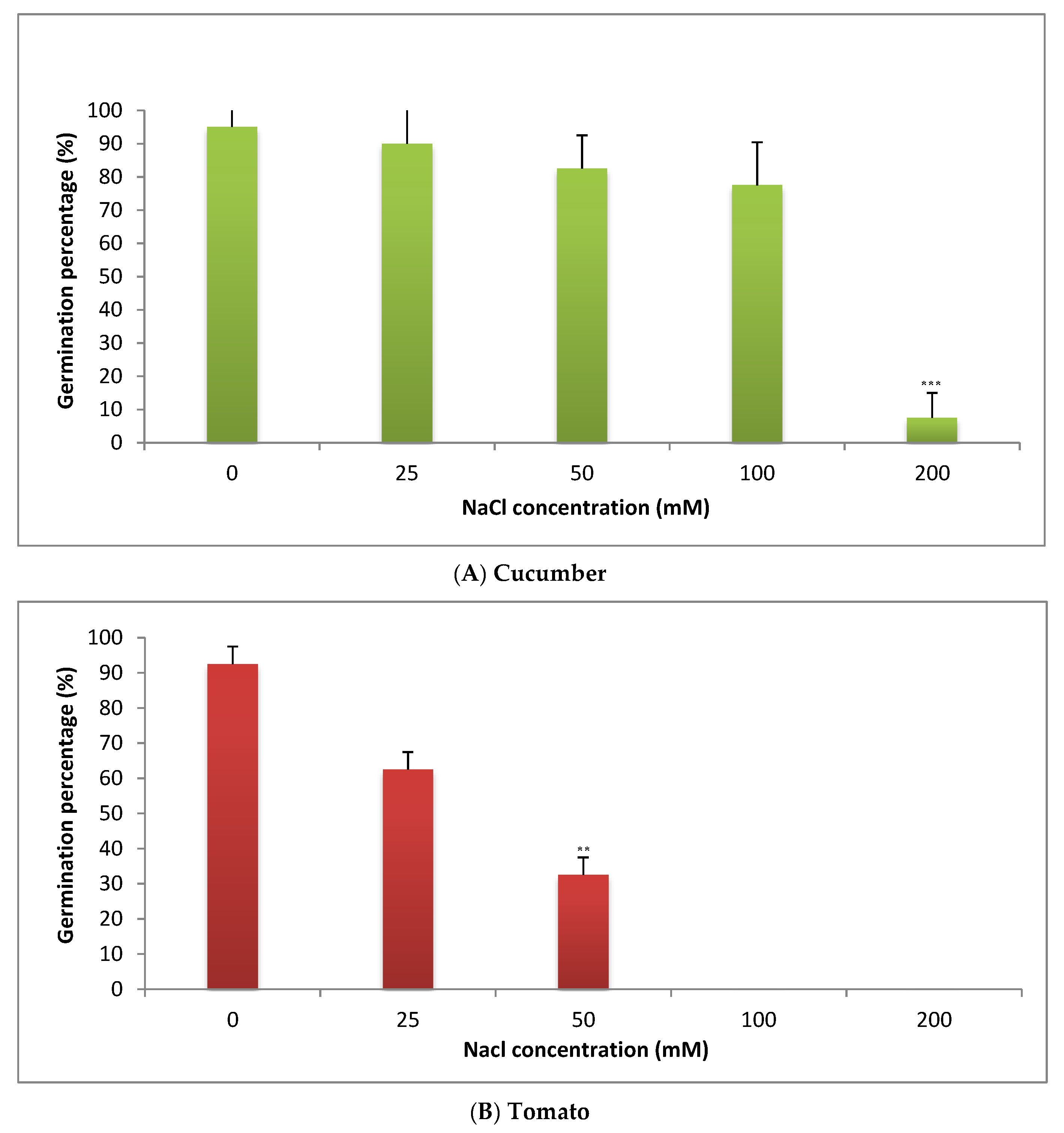
2.2. Effect of Salinity Stress on Percentage of Seeds Germination of C. sativus and S. lycopersicum
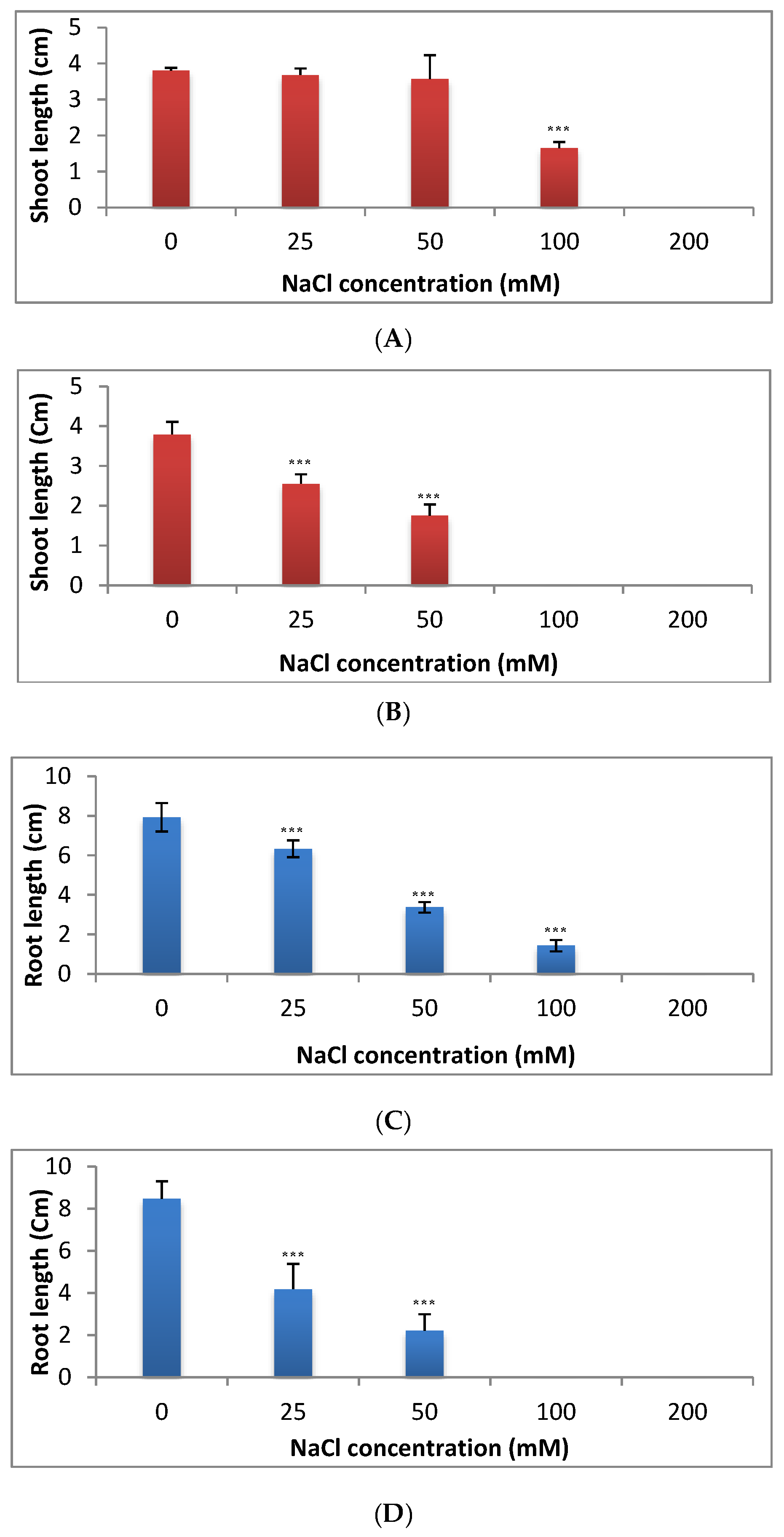
2.3. Effect of Salinity Stress on Seedlings Growth (Shoot and Root Lengths) in C. sativus and S. lycopersicum
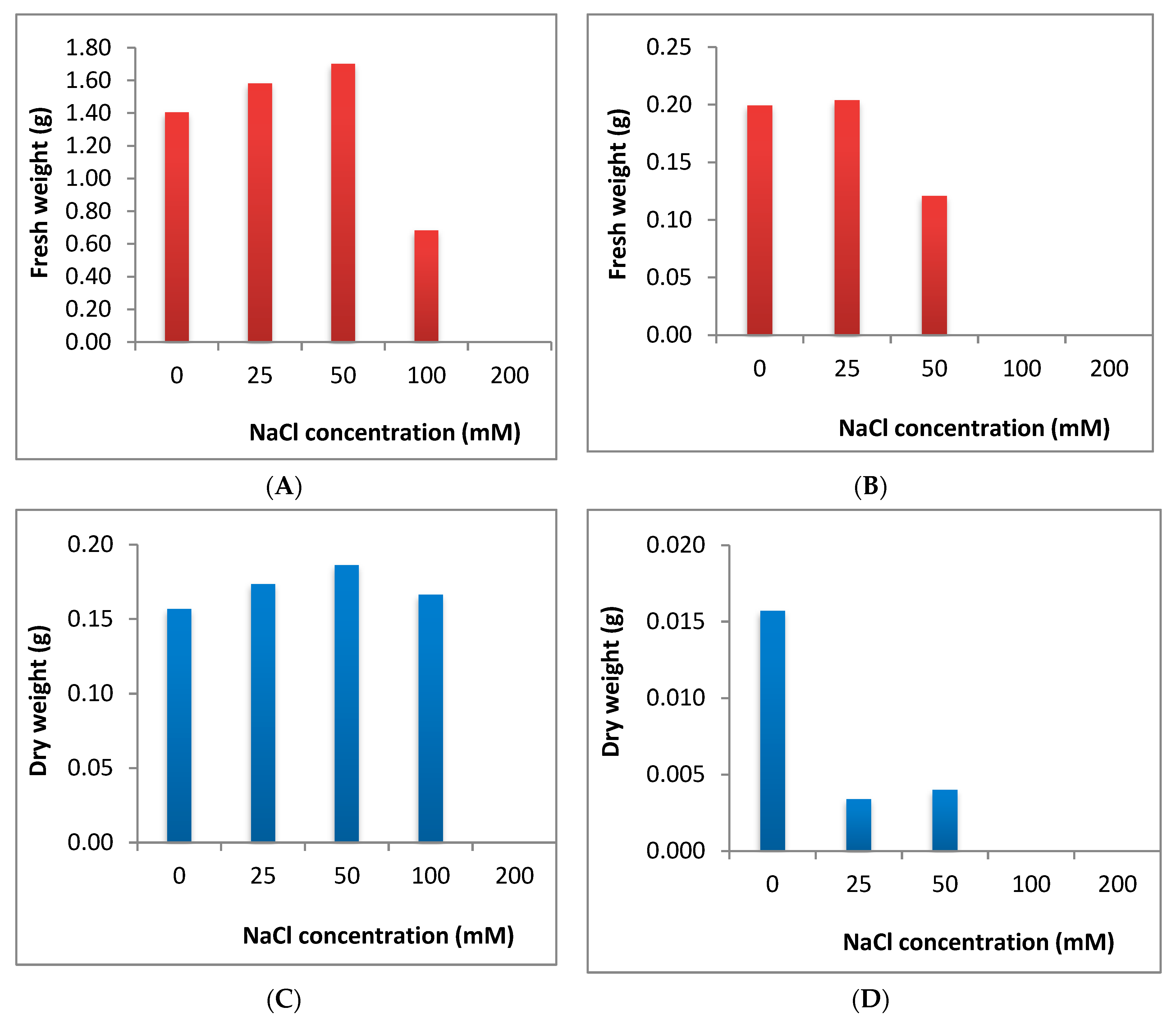
2.4. Effect of Salinity Stress on Fresh and Dry Weights in C. sativus and S. lycopersicum
2.5. Effect of Salinity Stress on Physiological and MetabolomicResponse in C. sativus and S. lycopersicum
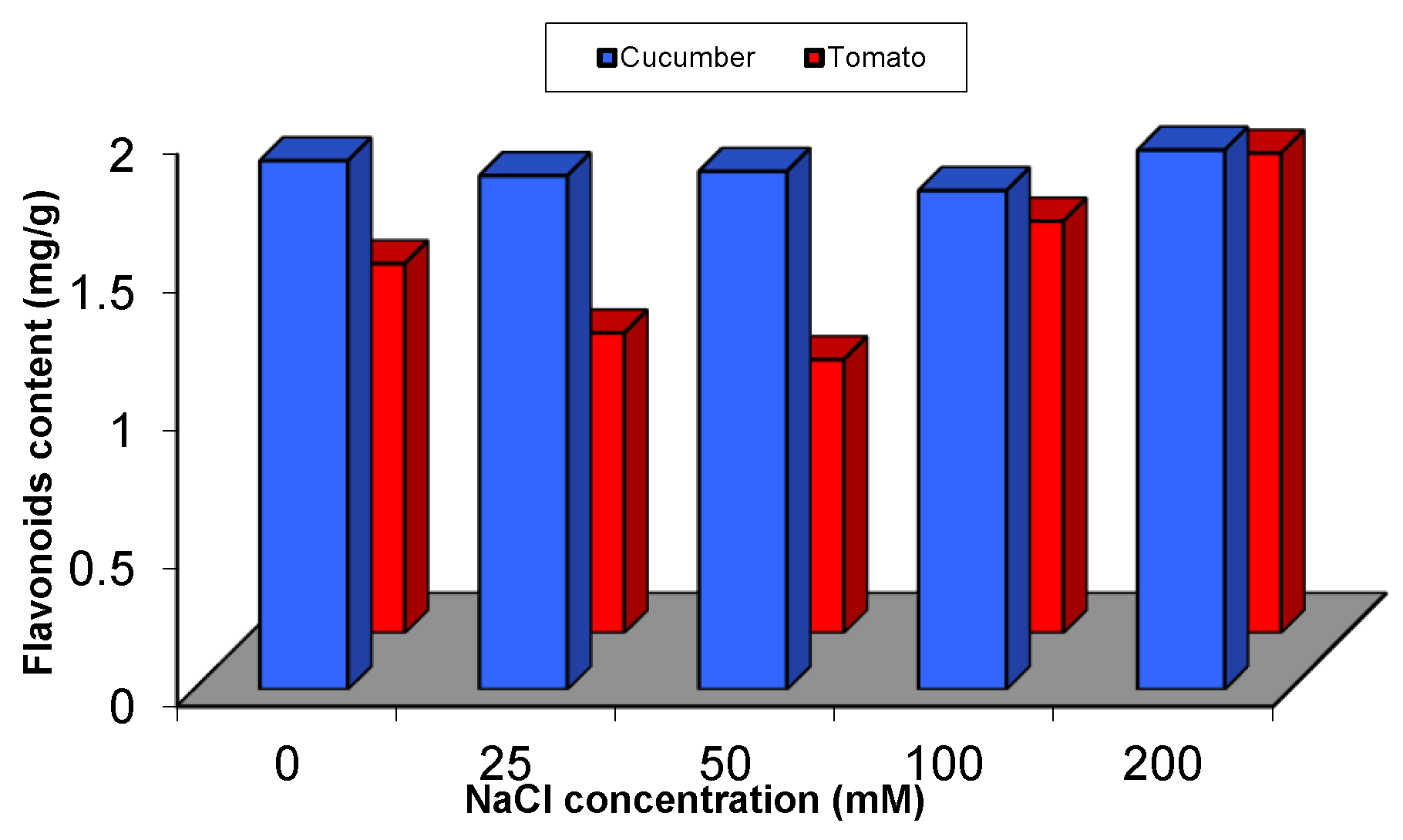
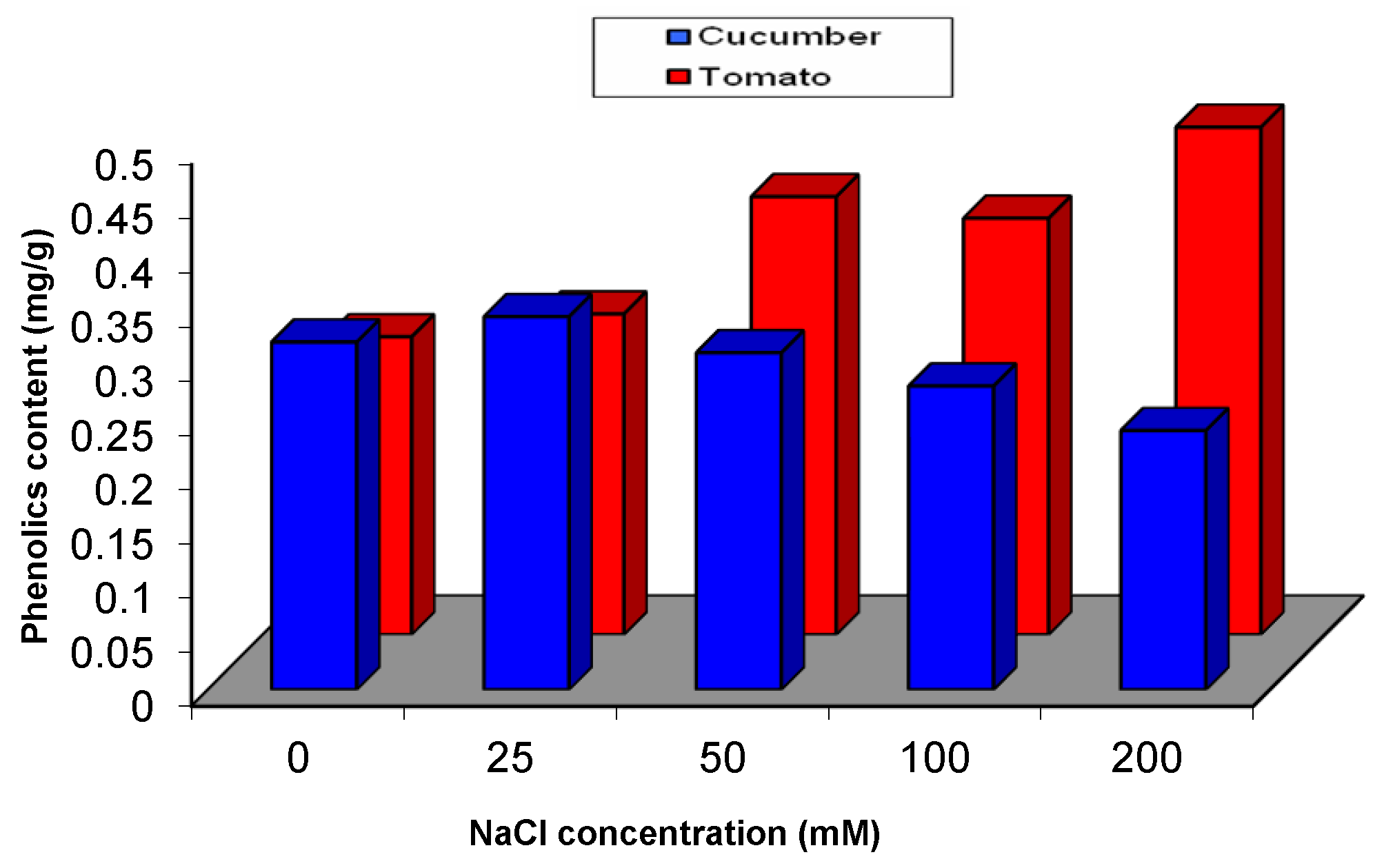
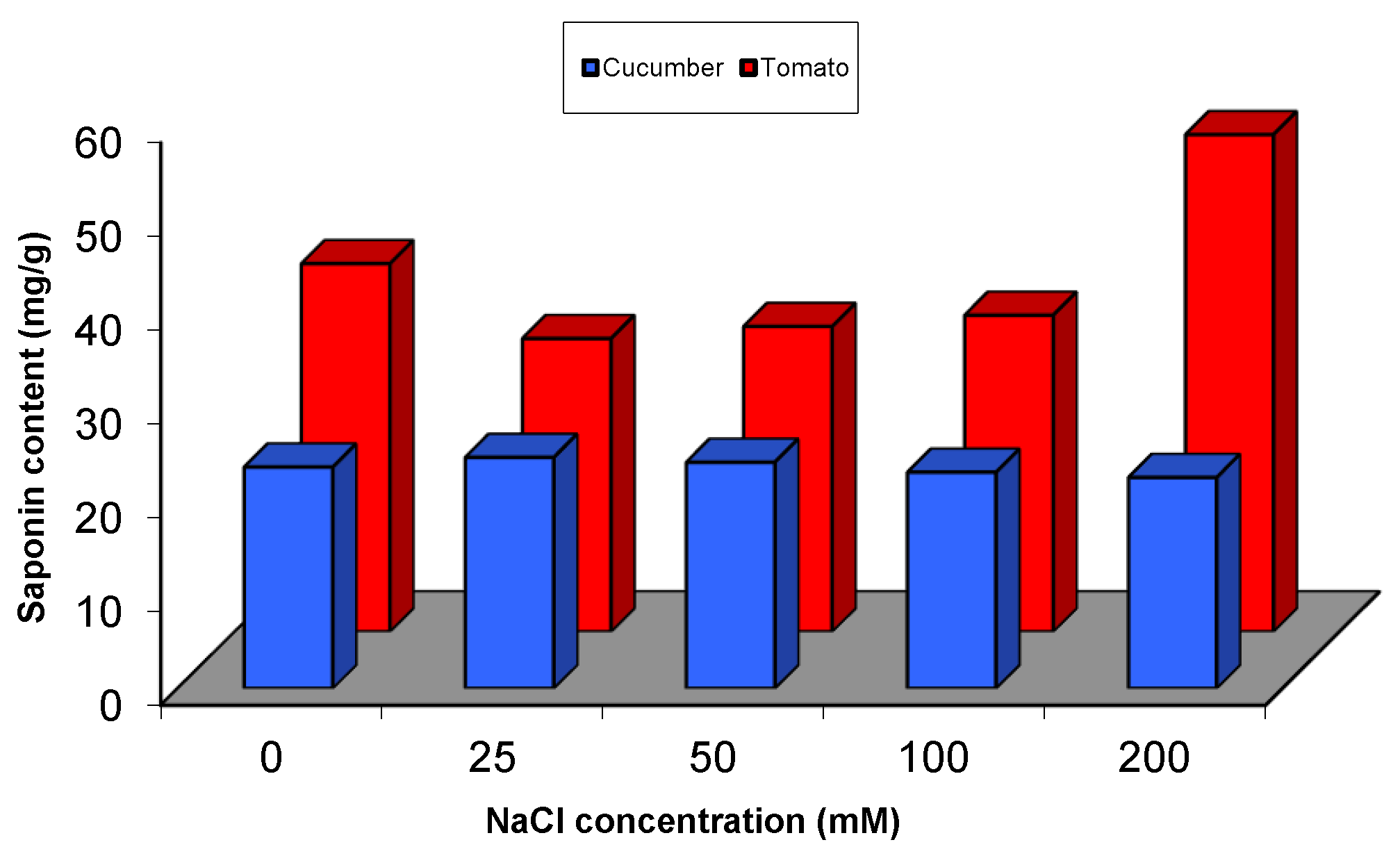
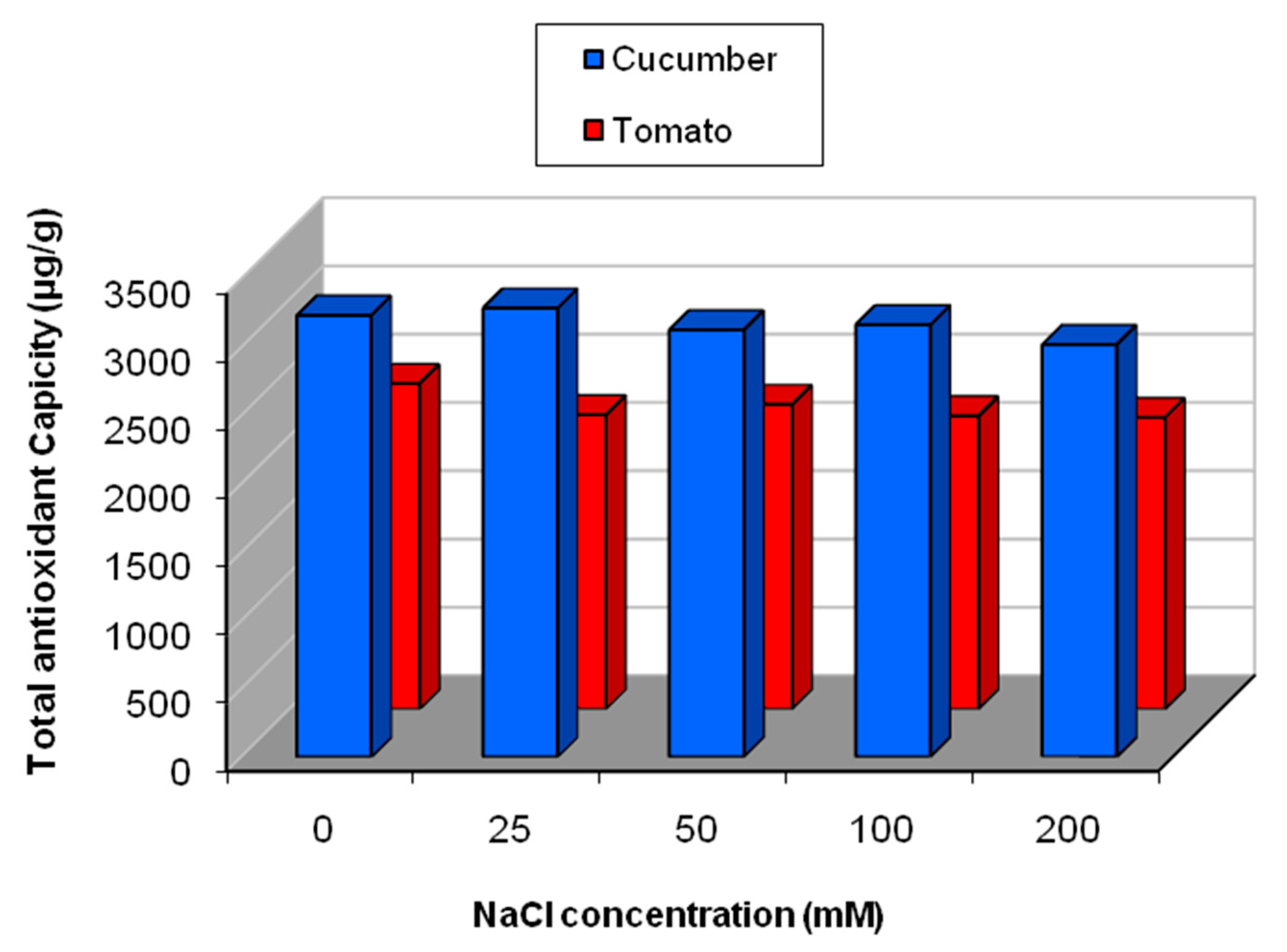
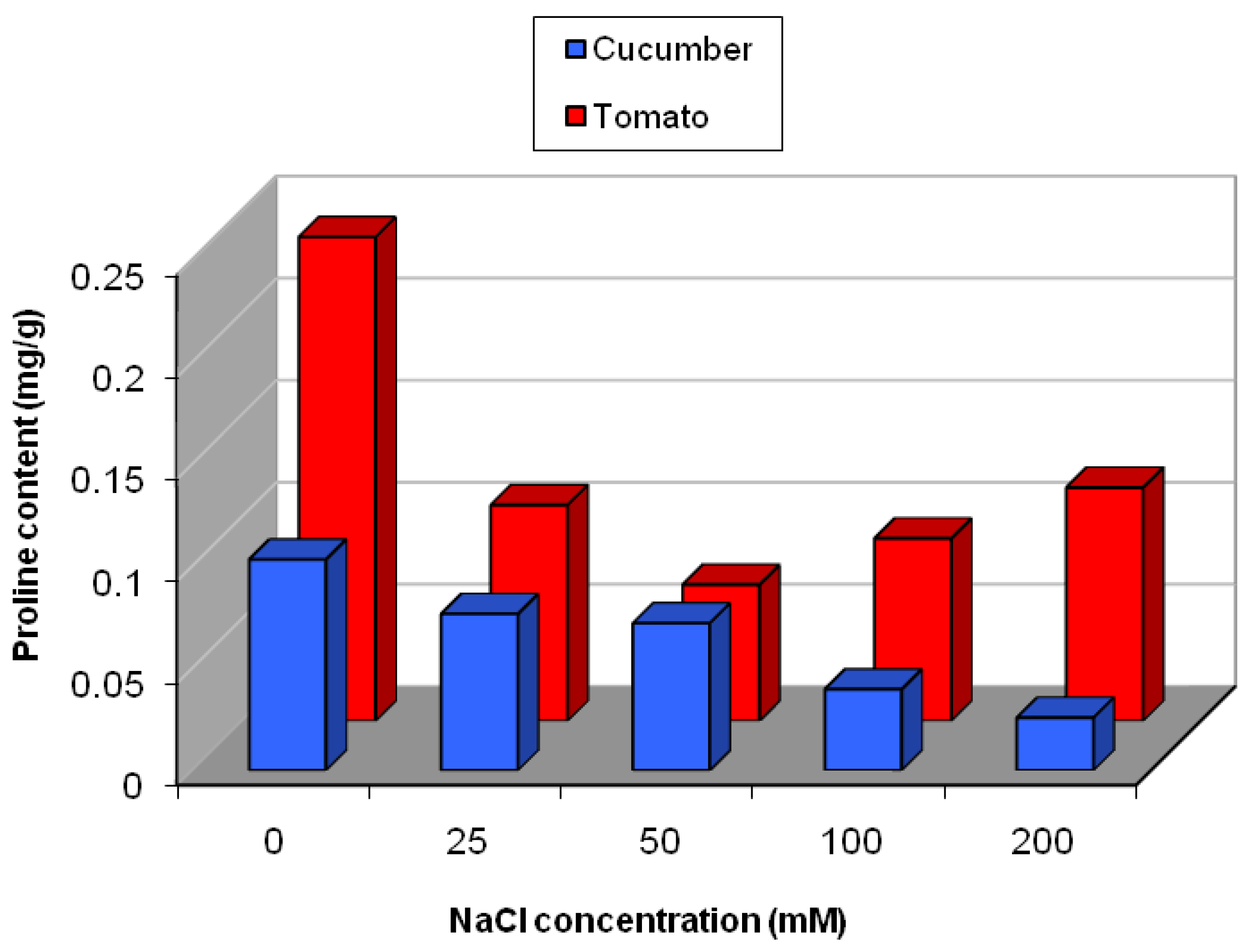
2.5.1. Effect of Salinity Stress on the Content of Secondary and Primary Metabolites
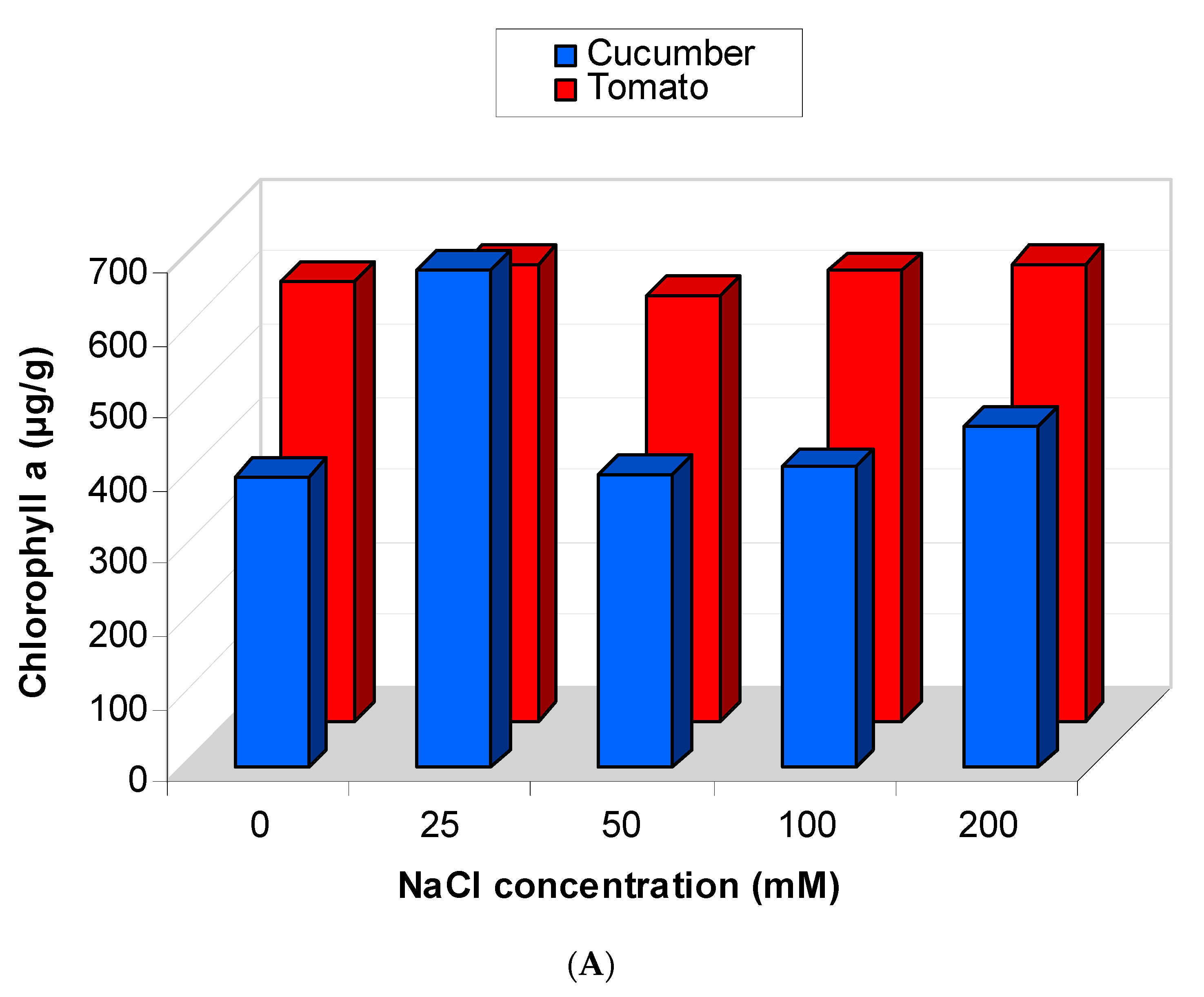
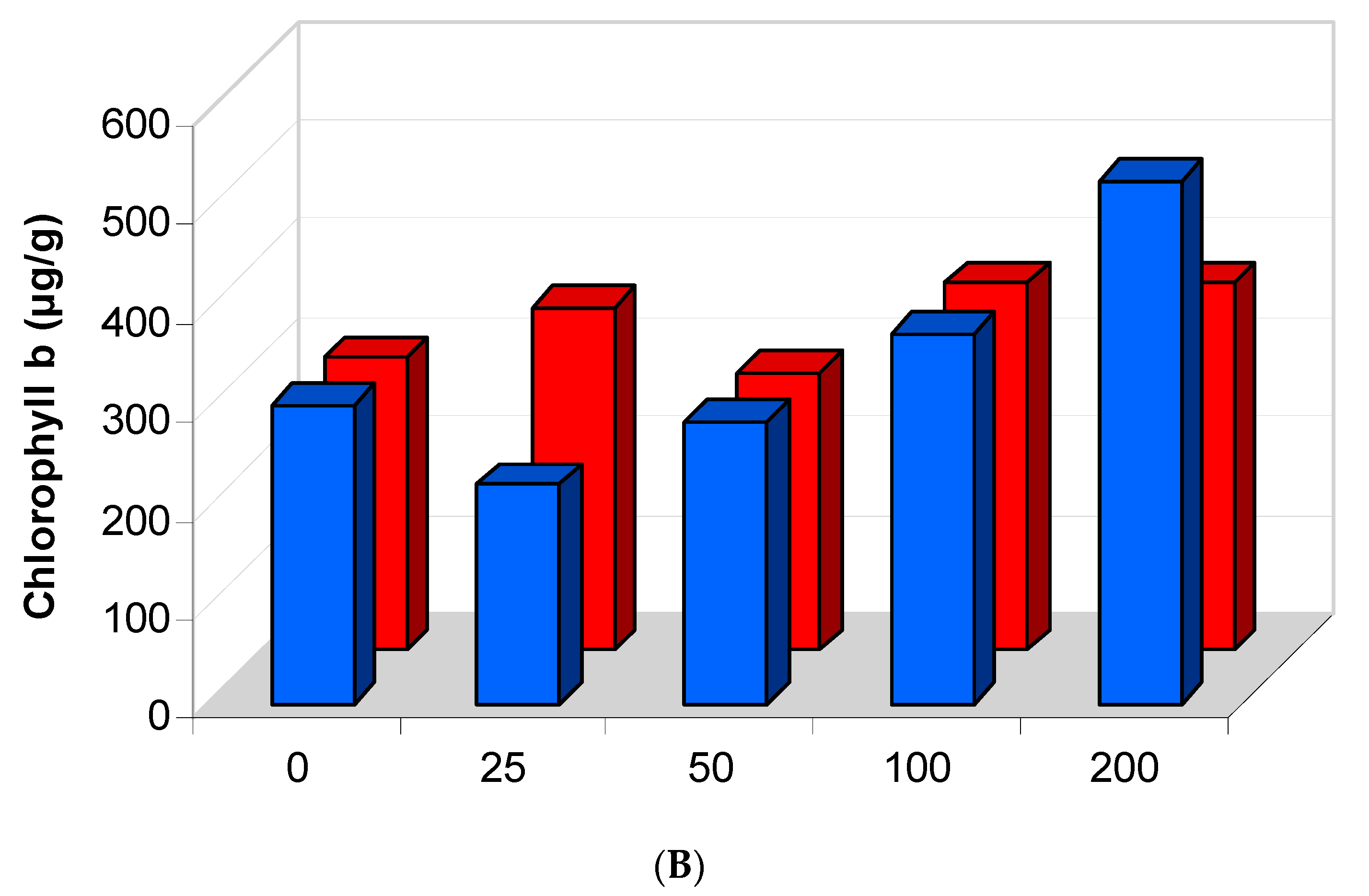
2.5.2. Effect of Salinity Stress on the Content of Photosynthetic Pigments
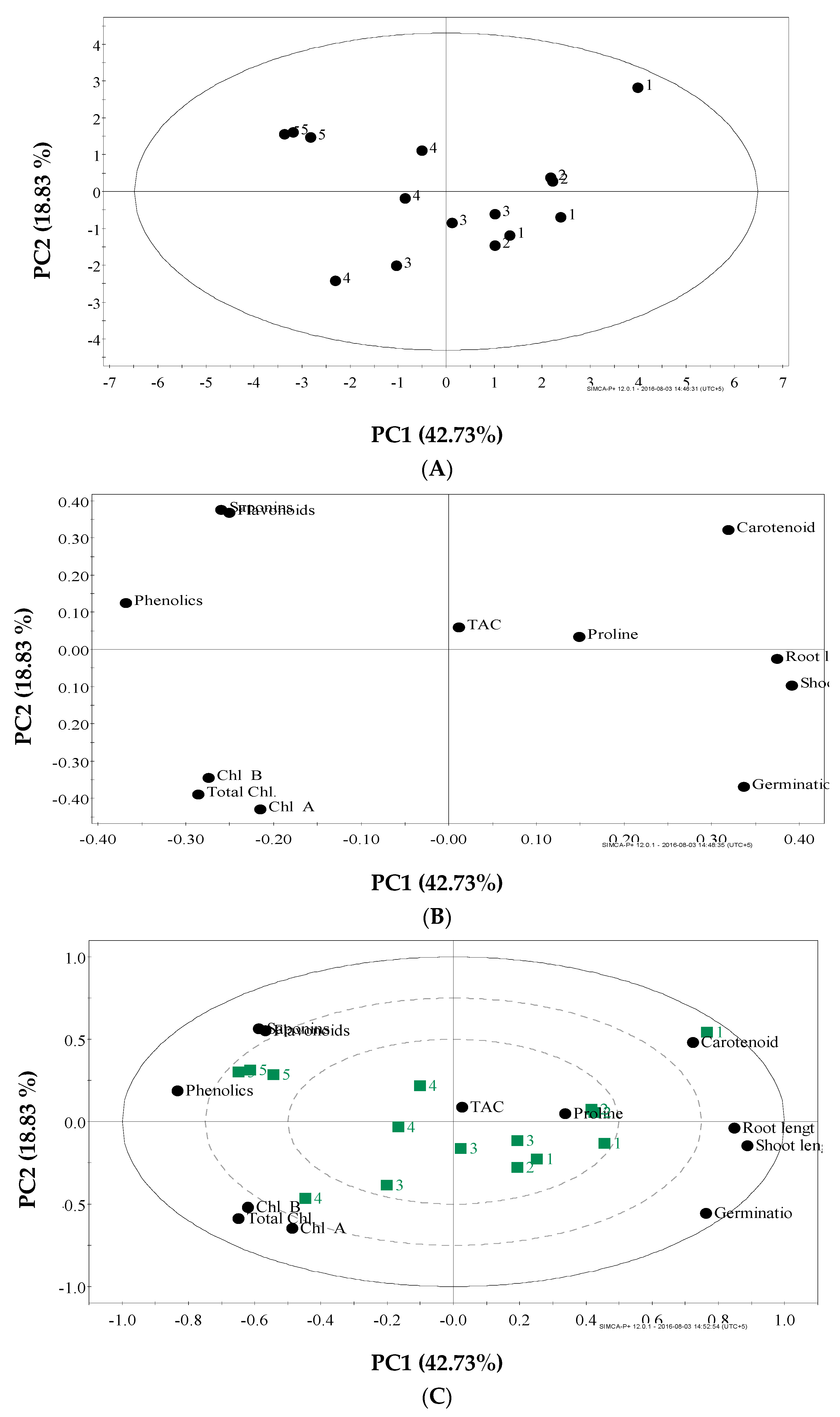
2.6. Dissecting the Effect of Salinity Stress on Tomato Using Metabolomic and Multivariate Data Analysis
3. Discussion
4. Materials and Methods
4.1. Germination Experiment
4.2. Pots Experiments
4.3. Plant Analysis
4.3.1. Determination of Total Phenolics (TP)
4.3.2. Determination of Total Flavonoid (TF)
4.3.3. Determination of Total Saponins
4.3.4. Determination of Total Antioxidant Capacity (TAC)
4.3.5. Determination of Free Proline
4.3.6. Determination of Photosynthetic Pigments
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Maas, E.V.; Hoffman, G.J. Crop salt tolerance-current assessment. J. Irrig. Drain. Div. 1977, 103, 115–134. [Google Scholar]
- Ali, Y.; Aslam, Z.; Ashraf, M.Y.; Tahir, G.R. Effect of salinity on chlorophyll concentration, leaf area, yield and yield components of rice genotypes grown under saline environment. Int. J. Environ. Sci. Technol. 2004, 1, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.T. An overview of hazardous impacts of soil salinity in crops, tolerance mechanisms, and amelioration through selenium supplementation. Int. J. Mol. Sci. 2020, 21, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonar, B.A.; Desai Nivas, D.; Gaikwad, D.K.; Chanven, P.D. Assessment of salinity-induced antioxidative defense system in Colubrina asiatica Brong. J. Stress Physiol. Biochem. 2011, 7, 193–200. [Google Scholar]
- Movafegh, S.; Jadid, R.R.; Kiabi, S. Effect of salinity stress on chlorophyll content, proline, water soluble carbohydrate, germination, growth and dry weight of three seedling barley (Hordeum vulgare L.) cultivars. J. Stress Physiol. Biochem. 2012, 8, 157–168. [Google Scholar]
- Miljuš-Djukić, J.; Stanisavljević, N.; Radović, S.; Jovanović, Ž.; Mikić, A.; Maksimović, V. Differential response of three contrasting pea (Pisum arvense, P. sativum and P. fulvum) species to salt stress: Assessment of variation in antioxidative defence and miRNA expression. Aust. J. Crop Sci. 2013, 7, 2145–2153. [Google Scholar]
- Singh, P.K.; Shahi, S.K.; Singh, A.P. Effects of salt stress on physic-chemical changes in maize (Zea mays L.) plants in response to salicylic acid. Indian J. Plant Sci. 2015, 4, 69–77. [Google Scholar]
- Saleh, B. Salt stress alters physiological indicators in cotton (Gossypium hirsutum L.). Soil Environ. 2012, 31, 113–118. [Google Scholar]
- Jamil, M.; Shik Rha, E. NaCl stress–induced reduction in growth, photosynthesis and protein in mustard. J. Agric. Sci. 2013, 5, 114–127. [Google Scholar] [CrossRef]
- Chandramohanan, K.T.; Radhakrishnan, V.V.; Abhilash Joseph, E.; Mohanan, K.V. A study on the effect of salinity stress on the chlorophyll content of certain rice cultivars of Kerala state of India. Agric. For. Fish. 2014, 3, 67–70. [Google Scholar] [CrossRef]
- Dhanapackiam, S.; Ilyas, M.H.M. Effect of salinity on chlorophyll and carbohydrate contents of Sesbania grandiflora seedlings. Indian J. Sci. Technol. 2010, 3, 64–66. [Google Scholar] [CrossRef]
- Abdul Qados, A.M.S. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba (L.). J. Saudi Soc. Agric. Sci. 2011, 10, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Heidari, M. Effects of salinity stress on growth, chlorophyll content and osmotic components of two basil (Ocimum basilicum L.) genotypes. Afr. J. Biotechnol. 2012, 11, 379–384. [Google Scholar] [CrossRef]
- Santos, C.V. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci. Hortic. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Masoumzadeh, B.M.; Imani, A.A.; Khayamaim, S. Salinity stress effect on proline and chlorophyll rate in four beet cultivars. Ann. Biol. Res. 2012, 3, 5453–5456. [Google Scholar] [CrossRef] [Green Version]
- Joshi, B.; Pandey, S.N.; Rao, P.B. Allelopathic effects of weeds extracts on germination, growth and biochemical aspects in different varieties of wheat (Triticum aestivum). Indian J. Agric. Res. 2009, 43, 79–87. [Google Scholar]
- Das, C.R.; Mondal, N.K.; Aditya, P.; Datta, J.K.; Banerjee, A.; Das, K. Allelopathic potentialities of leachates of leaf litter of some selected tree species on gram seeds under laboratory conditions. Asian J. Exp. Biol. Sci. 2012, 3, 59–65. [Google Scholar]
- Abdel-Farid, I.B.; El-Sayed, M.A.; Mohamed, E.A. Allelopathic potential of Calotropis procera and Morettia philaeana. Int. J. Agric. Biol. 2013, 15, 120–134. [Google Scholar]
- Hussain, M.; Park, H.W.; Farooq, M.; Jabran, K.; Lee, D.J. Morphological and physiological basis of salt resistance in different rice genotypes. Int.J. Agric. Biol. 2013, 15, 113–118. [Google Scholar]
- Abdel-Farid, I.B.; Jahangir, M.; van den Hondel, C.A.M.J.J.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Fungal infection-induced metabolites in Brassica rapa. Plant Sci. 2009, 176, 608–615. [Google Scholar] [CrossRef]
- Haghighi, Z.; Karimi, N.; Modarresi, M.; Mollayi, S. Enhancement of compatible solute and secondary metabolites production in Plantago ovata Forsk. by salinity stress. J. Med. Plants Res. 2012, 6, 3495–3500. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Ashraf, M.; Ali, Q. Response of two genetically diverse wheat cultivars to salt stress at different growth stages: Leaf lipid peroxidation and phenolic contents. Pak. J. Bot. 2010, 42, 559–565. [Google Scholar]
- Odjegba, V.J.; Alokolaro, A.A. Simulated drought and salinity modulates the production of phytochemicals in Acalypha wilkesiana. J. Plant Stud. 2013, 2, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Zapata, P.J.; Serrano, M.; Pretel, M.T.; Amorós, A.; Botella, M.Á. Polyamines and ethylene changes during germination of different plant species under salinity. Plant Sci. 2004, 167, 781–788. [Google Scholar] [CrossRef]
- Jamil, M.; Ur Rehman, S.; Lee, K.J.; Kim, J.M.; Kim, H.-S.; Shik Rha, E. Salinity reduced growth PS2 photochemistry and chlorophyll content in radish. Sci. Agric. 2007, 64, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Bybordi, A. The influence of salt stress on seed germination, growth and yield of canola cultivars. Not. Bot. Horti Agrobot. Cluj-Napoca 2010, 38, 128–133. [Google Scholar] [CrossRef]
- Turhan, A.; Kuşçu, H.; Şeniz, V. Effects of different salt concentrations (NaCl) on germination of some spinach cultivars. J. Agric. Fac. Uludag Univ. 2011, 25, 65–77. [Google Scholar]
- Foolad, M.R.; Lin, G.Y. Relationships between cold-and salt-tolerance during seed germination in tomato: Germplasm evaluation. Plant Breed. 1999, 118, 45–48. [Google Scholar] [CrossRef]
- Jones, R.A. High salt tolerance potential in Lycopersicon species during germination. Euphytica 1986, 35, 575–582. [Google Scholar] [CrossRef]
- Doğan, M.; Avu, A.; Can, E.N.; Aktan, A. Farklı domates tohumlarının çimlenmesiüzerine tuz stresinin etkisi. SDÜ Fen Edebiyat Fakültesi Fen Dergisi (E-DERGİ) 2008, 3, 174–182. (In Turkish) [Google Scholar]
- Passam, H.C.; Kakouriotis, D. The effects of osmoconditioning on the germination emergence and early plant growth of cucumber under saline conditions. Sci. Hortic. 1994, 57, 233–240. [Google Scholar] [CrossRef]
- Roussos, P.A.; Pontikis, C.A. Long term effects of sodium chloride salinity on growing in vitro, proline and phenolic compound content of jojoba explants. Eur. J. Hortic. Sci. 2003, 68, 38–44. [Google Scholar]
- Gharineh, M.H.; Khoddami, H.R.; Rafieian-kopaei, M. The influence of different levels of salt stress on germination of marigold (Calendula officinalis L.). Int. J. Agric. Crop Sci. 2013, 5, 1851–1854. [Google Scholar]
- Andriolo, J.L.; Luz, G.L.; Witter, M.H.; Godoi, R.S.; Barros, G.T.; Bortolotto, O.C. Growth and yield of lettuce plants under salinity. Hortic. Bras. 2005, 23, 931–934. [Google Scholar] [CrossRef] [Green Version]
- Dantus, B.F.; Ribeiro, L.; Aragao, C.A. Physiological response of cowpea seeds to salinity stress. Rev. Bras.Sem. 2005, 27, 144–148. [Google Scholar] [CrossRef] [Green Version]
- Niaz, B.H.; Athar, M.; Salim, M.; Rozema, J. Growth and ionic relations of fodder beet and sea beet under saline. Int. J. Environ. Sci. Technol. 2005, 2, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Nedjimi, B.; Daoud, Y.; Touati, M. Growth, water relations, proline and ion content of in vitro cultured Atriplex halimus sub sp. Schweinfurthii as affected by CaCl2. Commun. Biom. Crop Sci. 2006, 1, 79–89. [Google Scholar]
- Memon, S.A.; Hou, X.; Wang, L.J. Morphological analysis of salt stress response of pak Choi. Electron. J. Environ. Agric. Food Chem. 2010, 9, 248–254. [Google Scholar]
- Sevengor, S.; Yaşar, F.; Kusvuran, S.; Ellialtıoğlu, S. The effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidative enzymes of pumpkin seedling. Afr. J. Agric. Res. 2011, 6, 4920–4924. [Google Scholar] [CrossRef]
- Al Hassan, M.; Fuertes, M.M.; Sánchez, F.J.R.; Vicente, O.; Boscaiu, M. Effects of salt and water stress on plant growth and on accumulation of osmolytes and antioxidant compounds in cherry tomato. Not. Bot. Horti Agrobot. Cluj-Napoca 2015, 43, 1–11. [Google Scholar] [CrossRef]
- Gómez-Caravaca, A.M.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Caboni, M.F. Simultaneous determination of phenolic compounds and saponins in quinoa (Chenopodium quinoa Willd) by a liquid chromatography-diode array detection-electrospray ionization-time-of-flight mass spectrometry methodology. J. Agric. Food Chem. 2011, 59, 10815–10825. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.-H.; Park, K.-J.; Kim, B.-K.; Jeong, J.W.; Kim, H.-J. Effect of salinity stress on phenolic compounds and carotenoids in buckwheat (Fagopyrum esculentum M.) sprout. Food Chem. 2012, 135, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Radi, A.A.; Farghaly, F.A.; Hamada, A.M. Physiological and biochemical responses of salt–tolerant and salt–sensitive wheat and bean cultivars to salinity. J. Biol. Earth Sci. 2013, 3, B72–B88. [Google Scholar]
- Reginato, M.A.; Castagna, A.; Furlán, A.; Castro, S.; Ranieri, A.; Luna, V. Physiological responses of a halophytic shrub to salt stress by Na2SO4 and NaCl: Oxidative damage and the role of polyphenols in antioxidant protection. AOB Plants 2014, 6, 1–13. [Google Scholar] [CrossRef]
- Attaran, E. Canadian Society of Plant Physiologists/La Société Canadiene de Physiologie Végétale, Annual Meeting. In Proceedings of the Evaluation of the Effect of Salinity Stress on Saponin Contents in Bellis Perrenis L., Guelph, ON, Canada, 19–22 June 2004. [Google Scholar]
- Docimo, T.; De Stefano, R.; Cappetta, E.; Piccinelli, A.L.; Celano, R.; De Palma, M.; Tucci, M. Physiological, biochemical, and metabolic responses to short and prolonged saline stress in two cultivated cardoon genotypes. Plants 2020, 9, 554. [Google Scholar] [CrossRef]
- Abdel-Farid, I.B.; Sheded, M.G.; Mohamed, E.A. Metabolomic profiling and antioxidant activity of some Acacia species. Saudi J. Biol. Sci. 2014, 21, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Farid, I.B.; Mahalel, U.A.; Jahangir, M.; Elgebaly, H.A.; El-Naggar, S.A. Metabolomic profiling and antioxidant activity of Opophytum forsskalii. Aljouf Univ. Sci. Eng. J. 2016, 3, 19–24. [Google Scholar] [CrossRef]
- El-Naggar, S.; Abdel-Farid, I.B.; Elgebaly, H.A.; Germoush, M.O. Metabolomic profiling, antioxidant capacity and in vitro anticancer activity of some compositae plants growing in Saudi Arabia. Afr. J. Pharm. Pharmacol. 2015, 9, 764–774. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Kasim, W.A.; Dowidar, S. Amino acids and soluble protein profile of radish seedlings under salt stress as affected by GA3 priming. Indian J. Plant Physiol. 2006, 11, 75–82. [Google Scholar] [CrossRef]
- Misra, A.; Sahu, A.N.; Misra, M.; Singh, P.; Meera, I.; Das, N.; Kar, M.; Sahu, P. Sodium chloride induced changes in leaf growth, and pigment and protein contents in two rice cultivars. Biol. Plant. 1997, 39, 257–262. [Google Scholar] [CrossRef]
- Bai, Y.; Kissoudis, C.; Yan, Z.; Visser, R.G.; van der Linden, G. Plant behaviour under combined stress: Tomato responses to combined salinity and pathogen stress. Plant J. 2018, 93, 781–793. [Google Scholar] [CrossRef] [Green Version]
- El-Khatib, A.A.; Barakat, N.A.; Nazeir, H. Growth and physiological response of some cultivated species under allelopathic stress of Calotropis procera (Aiton) W.T. Appl. Sci. Rep. 2016, 14, 237–246. [Google Scholar] [CrossRef] [Green Version]
- El-Khatib, A.A.; Abd-Elaah, G.A. Allelopathic potential of Zilla spinosa on growth of associate flowering plants and some rhizosphere fungi. Biol. Plant. 1998, 41, 461–467. [Google Scholar] [CrossRef]
- Grange, S.L.; Leskovar, D.I.; Pike, L.M.; Cobb, B.G. Excess moisture and seed coat nicking influence germination of triploid watermelon. HortScience 2000, 35, 1355–1356. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthifer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-ciocalteau reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Ebrahimzadeh, H.; Niknam, V. A revised spectrophotometric method for determination of triterpenoid saponins. Indian Drugs 1998, 35, 379–381. [Google Scholar]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, T.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determination of total carotenoids and chlorophylls a and b of leaf in different solvents. Biochem. Soc. Trans. 1985, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Dere, S.; Güneş, T.; Sivaci, R. Spectrophotometric determination of chlorophyll A, B and total carotenoid contents of some algae species using different solvents. Tur. J. Bot. 1998, 22, 13–17. [Google Scholar]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Farid, I.B.; Marghany, M.R.; Rowezek, M.M.; Sheded, M.G. Effect of Salinity Stress on Growth and Metabolomic Profiling of Cucumis sativus and Solanum lycopersicum. Plants 2020, 9, 1626. https://doi.org/10.3390/plants9111626
Abdel-Farid IB, Marghany MR, Rowezek MM, Sheded MG. Effect of Salinity Stress on Growth and Metabolomic Profiling of Cucumis sativus and Solanum lycopersicum. Plants. 2020; 9(11):1626. https://doi.org/10.3390/plants9111626
Chicago/Turabian StyleAbdel-Farid, Ibrahim Bayoumi, Marwa Radawy Marghany, Mohamed Mahmoud Rowezek, and Mohamed Gabr Sheded. 2020. "Effect of Salinity Stress on Growth and Metabolomic Profiling of Cucumis sativus and Solanum lycopersicum" Plants 9, no. 11: 1626. https://doi.org/10.3390/plants9111626