Advancements in Low-Chill Blueberry Vaccinium corymbosum L. Tissue Culture Practices



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Introduction to Tissue Culture from Field Conditions
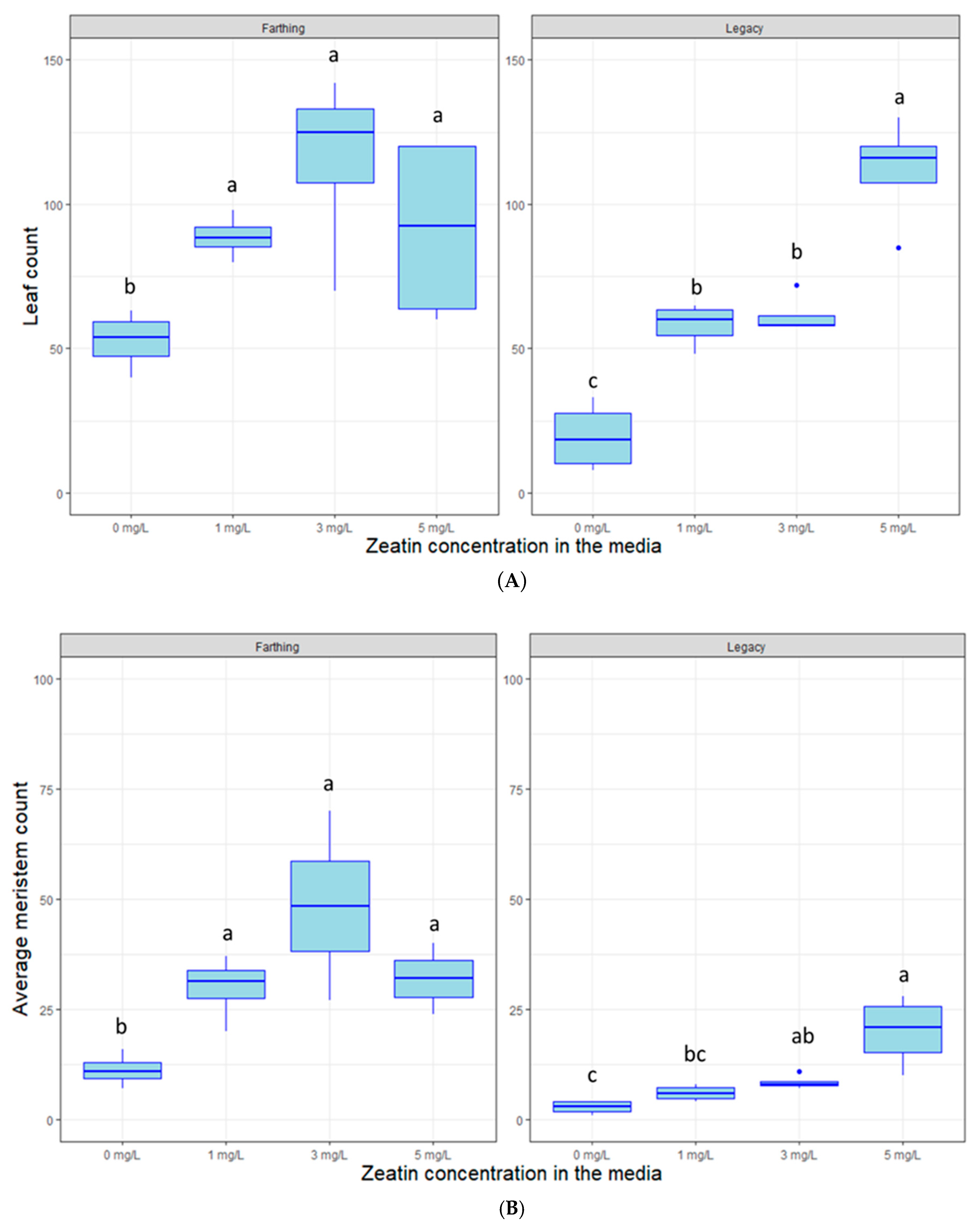
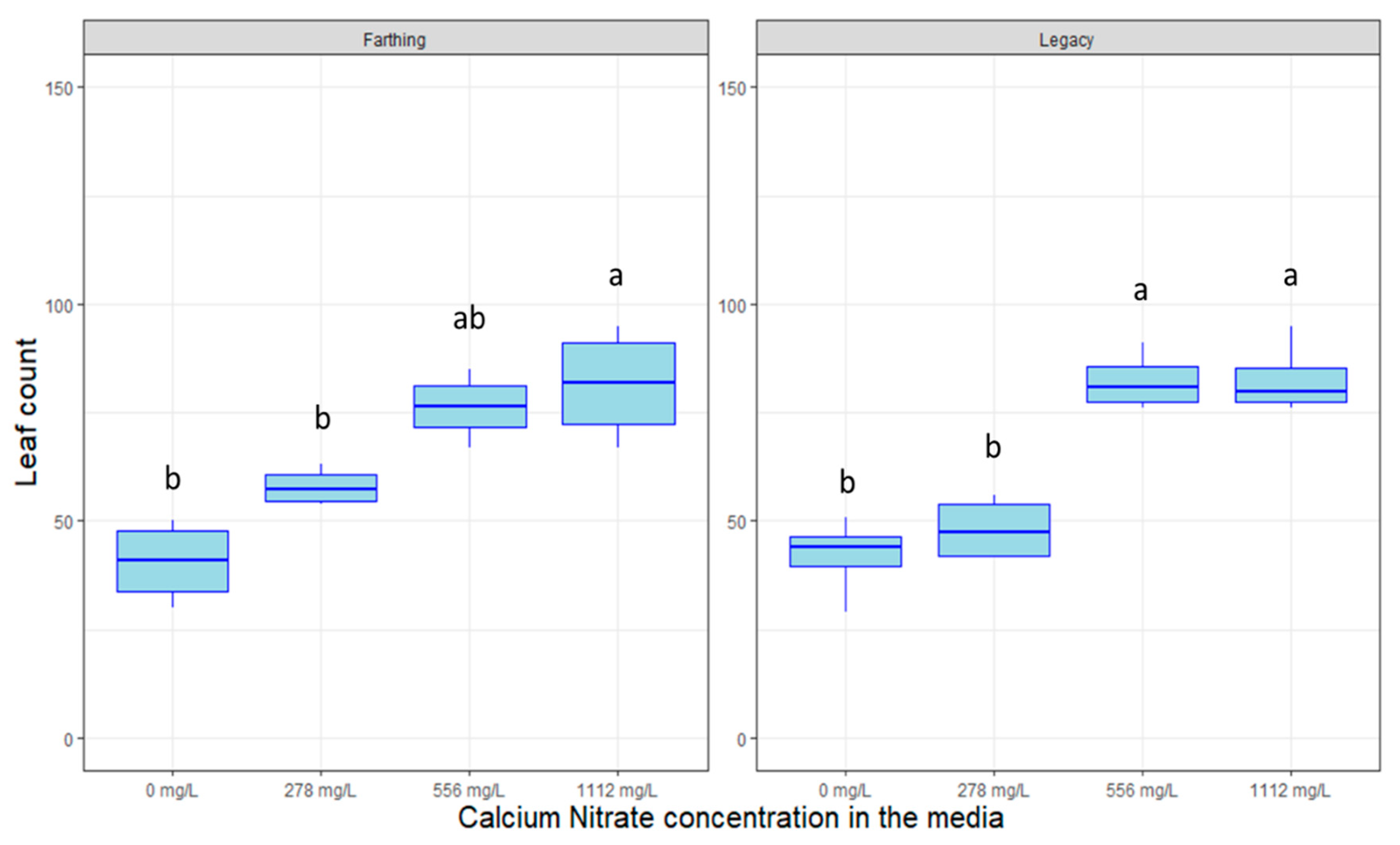
2.2. Hormonal Dosage and Nutrient Optimization
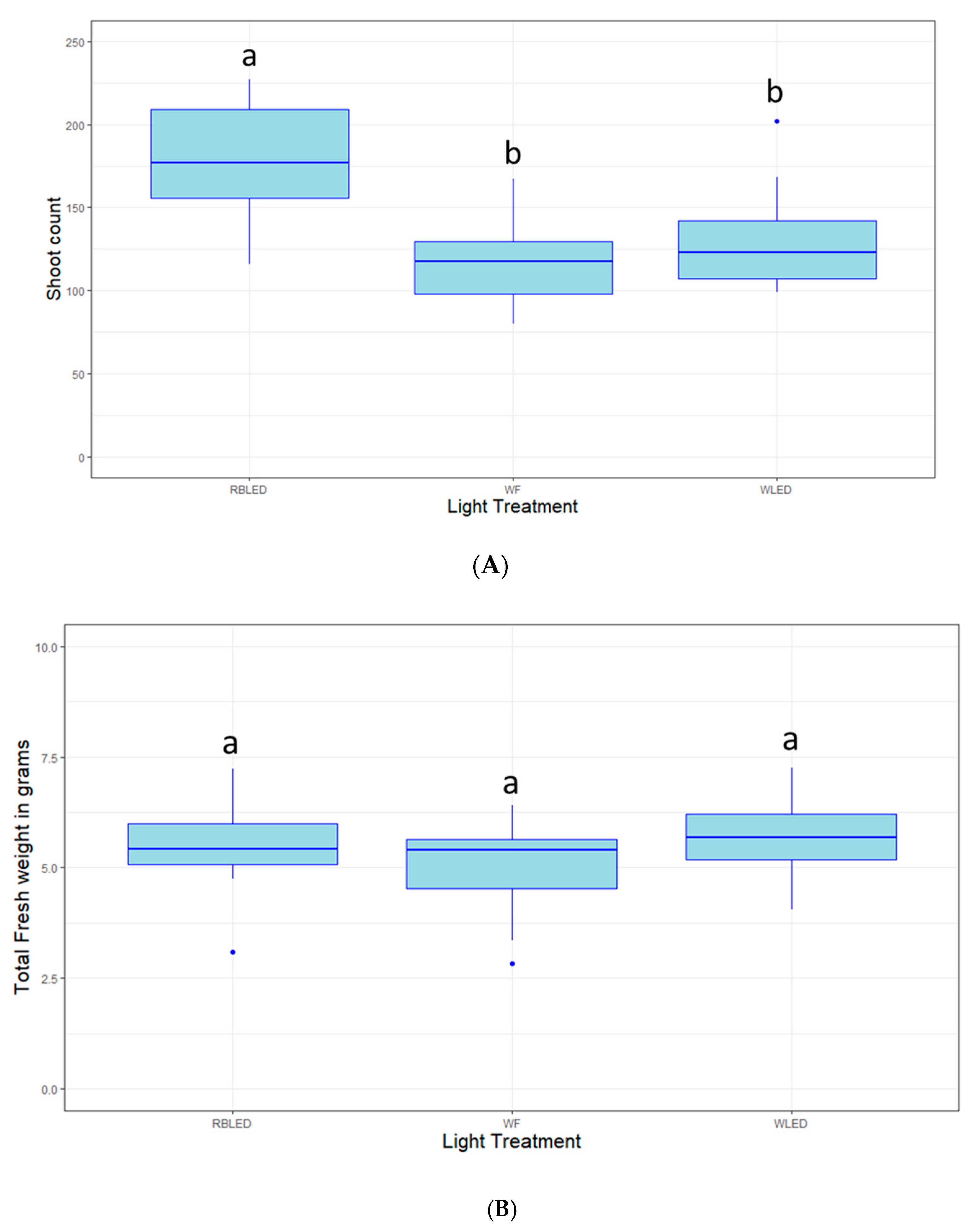
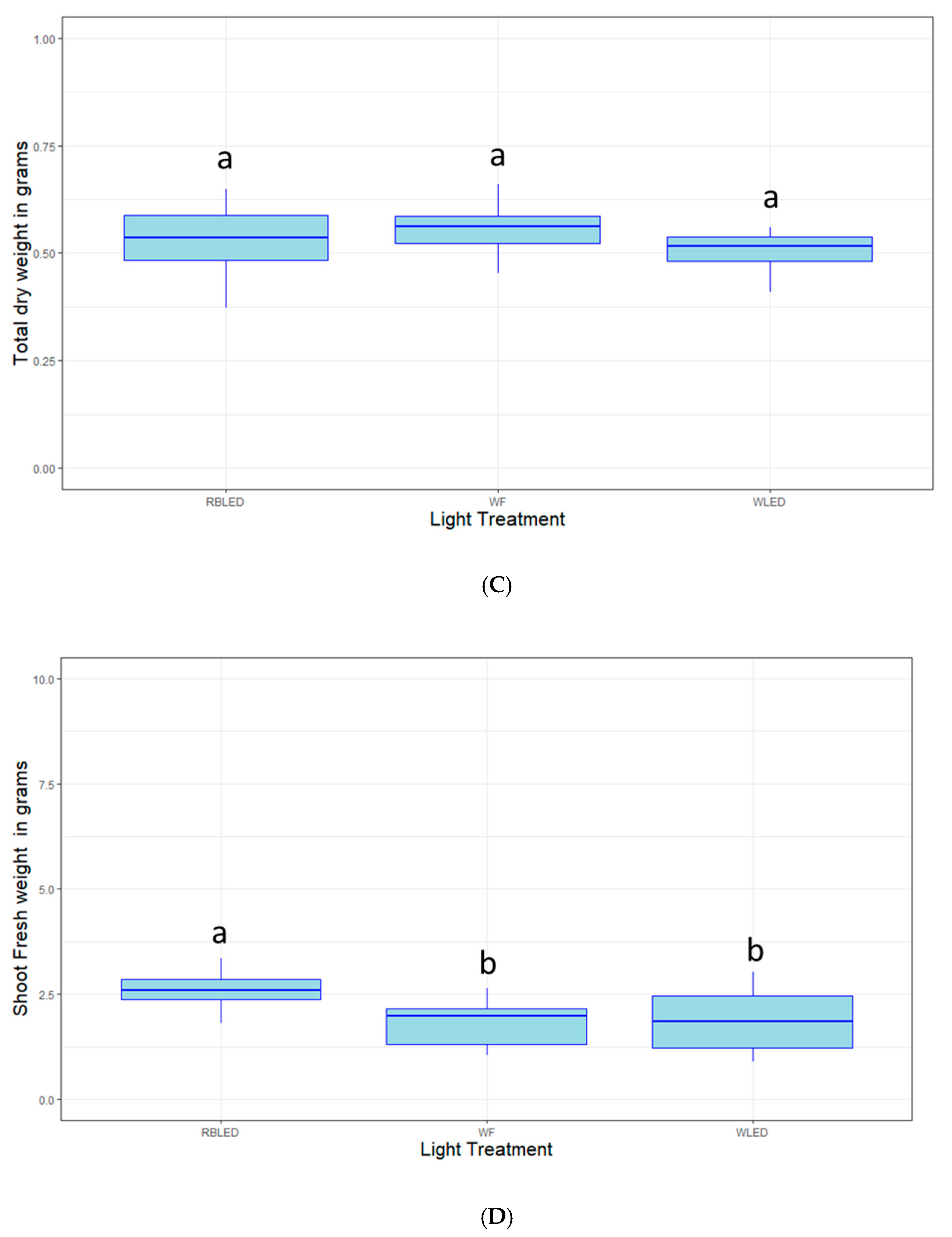
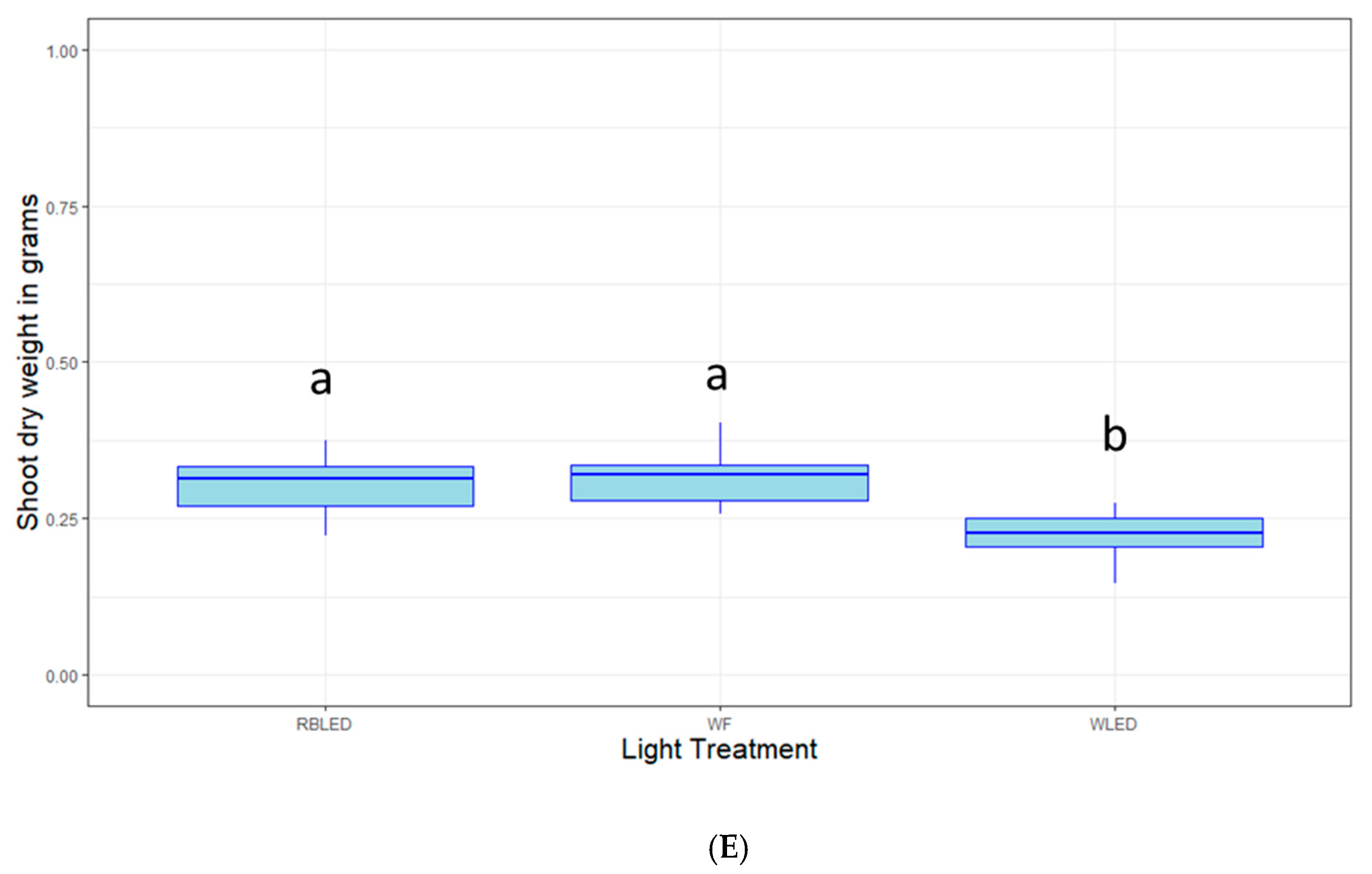
2.3. Light Quality Optimization
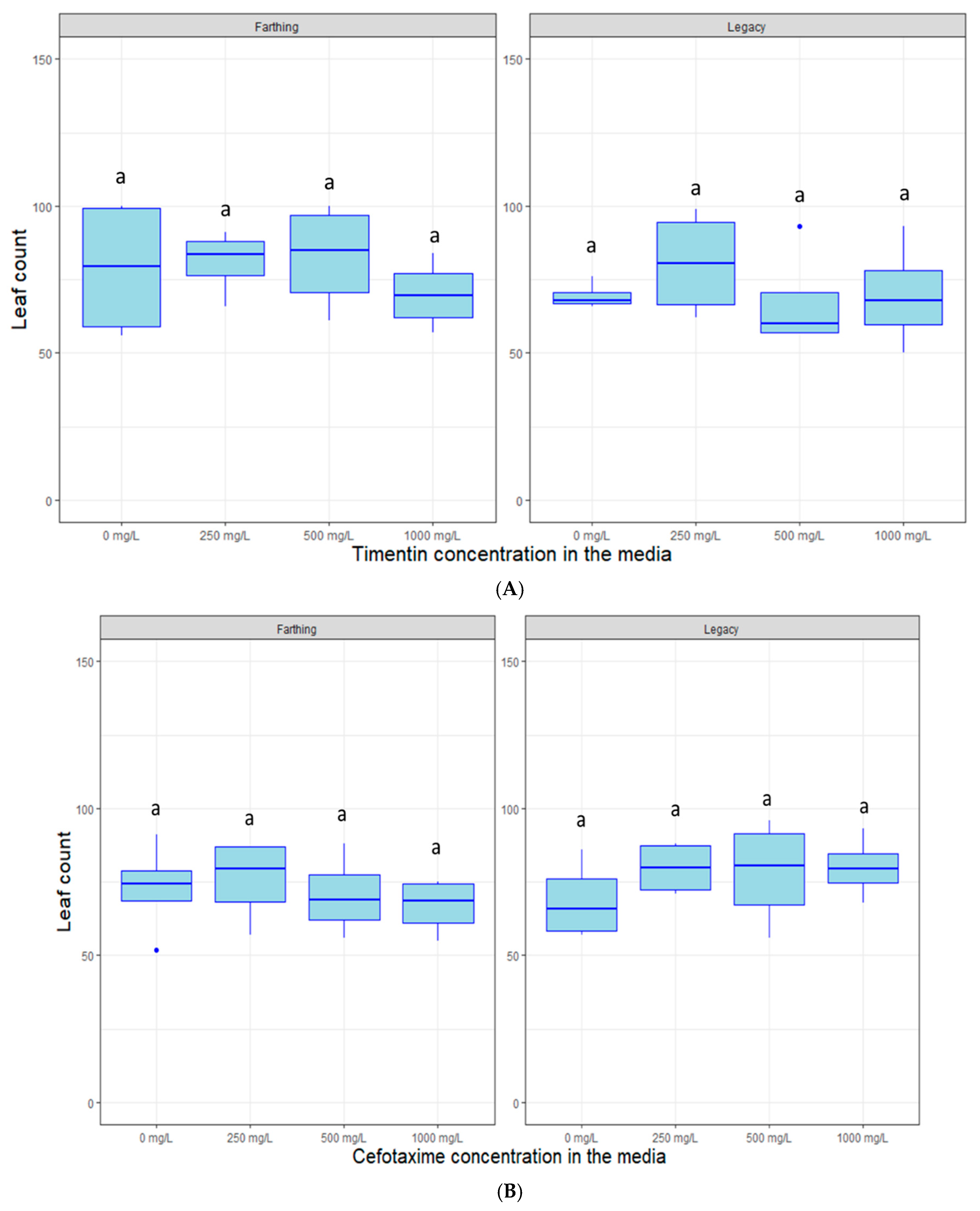
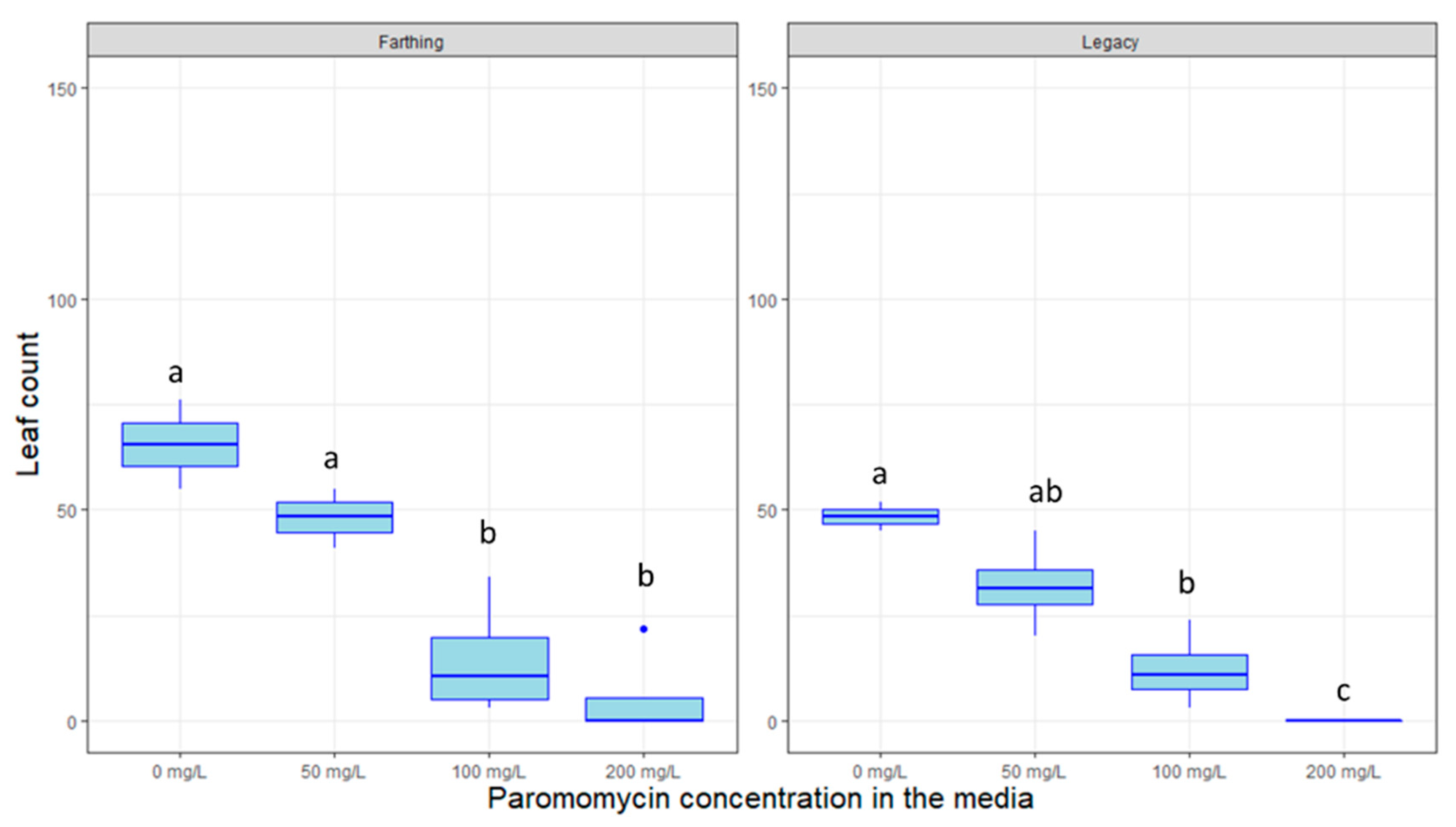
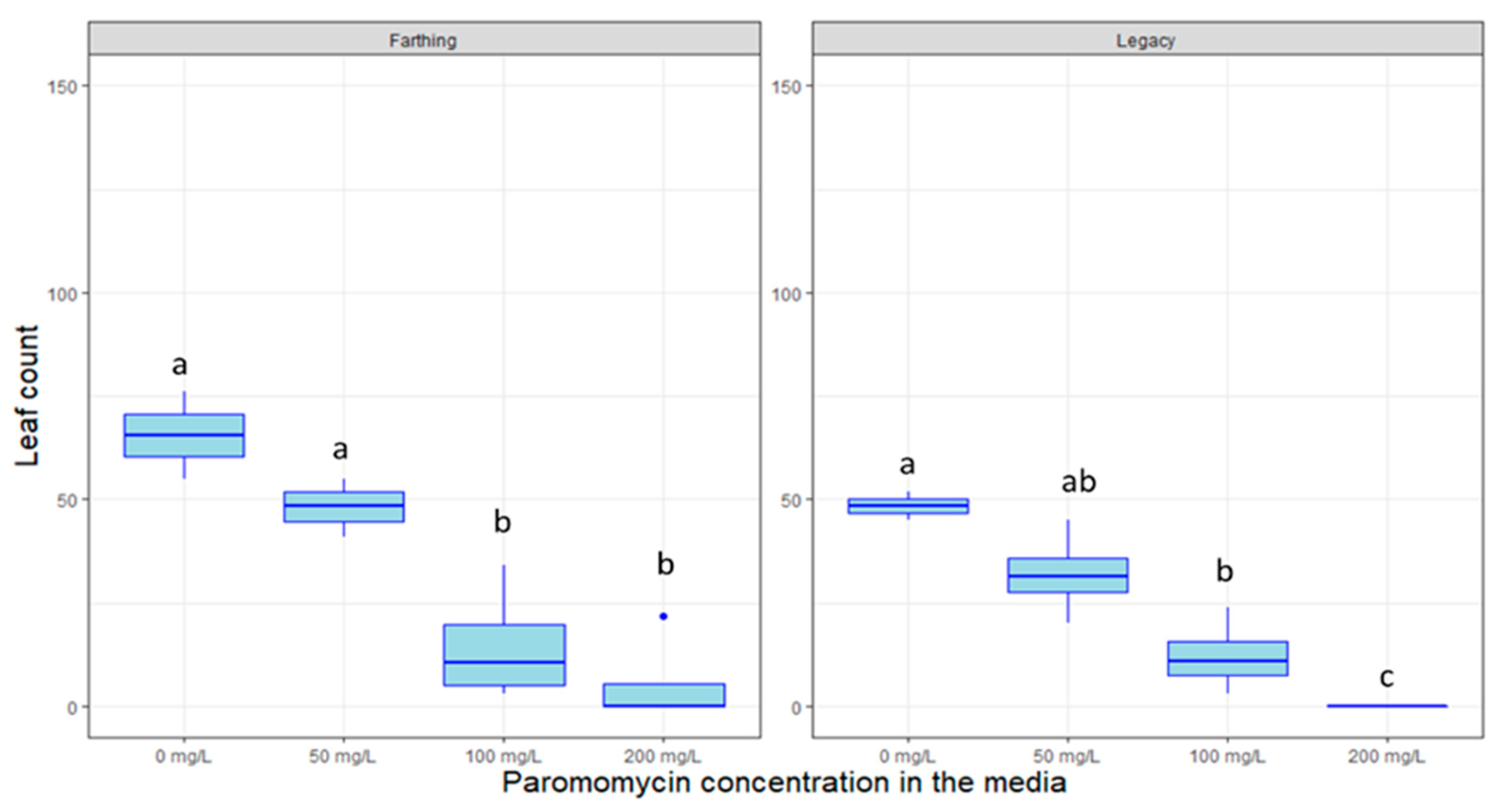
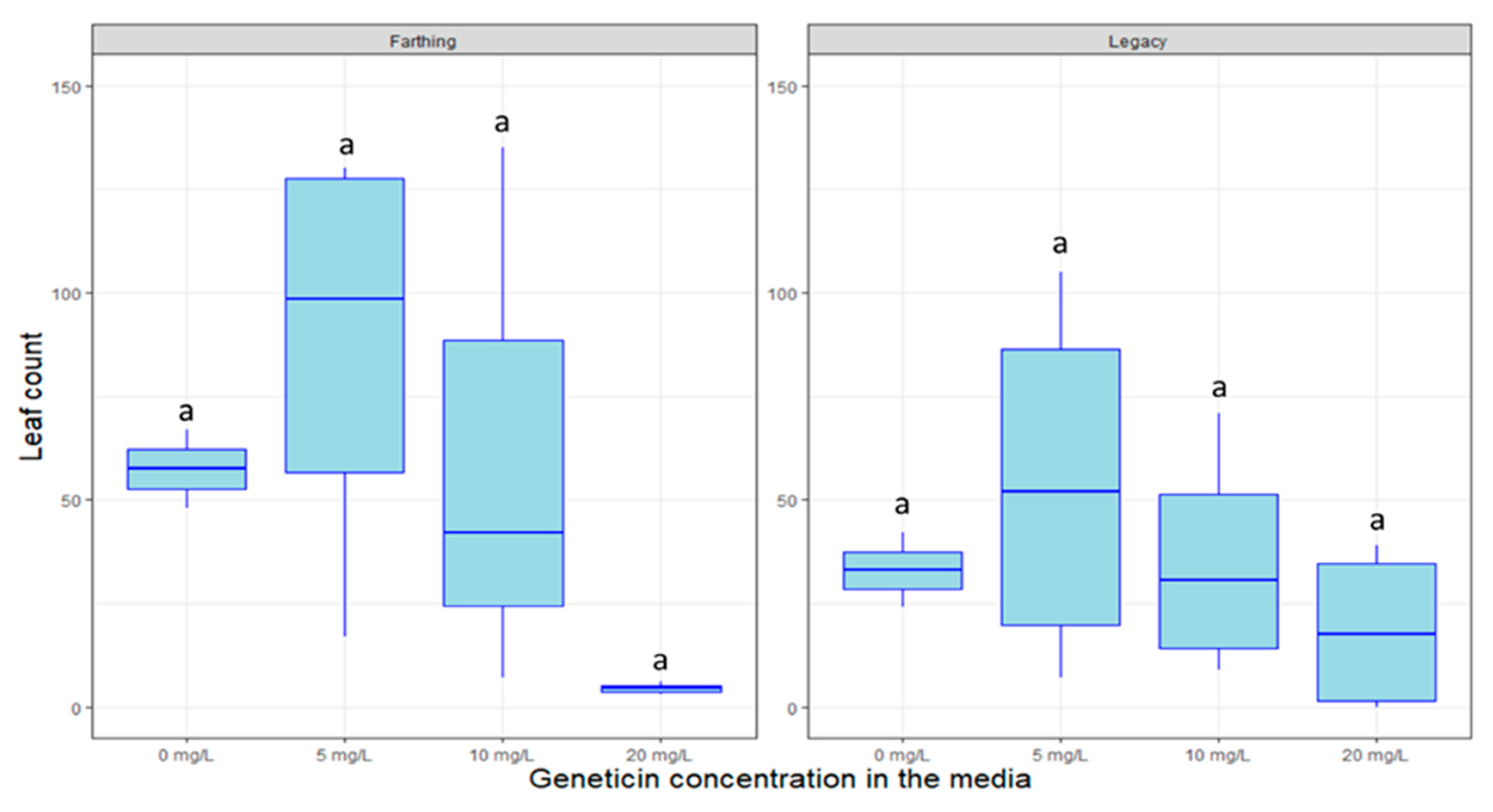
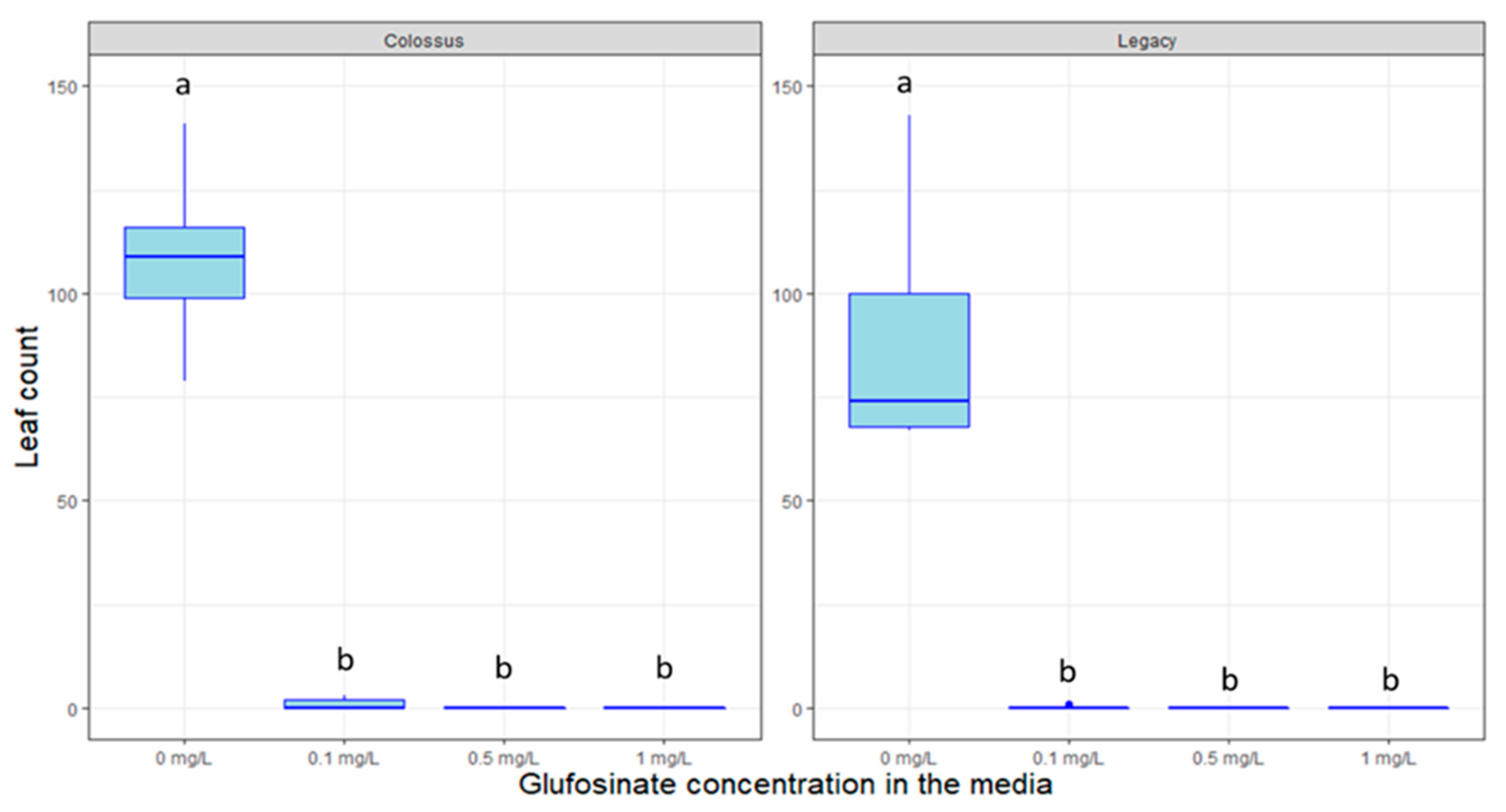
2.4. Response to Decontamination Agents and Selectable Markers
3. Discussion
4. Materials and Methods
4.1. Plant Genotypes and Growth Conditions
4.2. Introduction to Tissue Culture from Field Conditions
4.3. Hormonal Dosage and Nutrient Optimization
4.4. Light Quality Optimization
4.5. Response to Selectable Markers and Decontamination Agents
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krewer, G.; Cline, B. Blueberry Propagation Suggestions; Southern Region Small Fruit Consortium: Tifton, GA, USA, 2003. [Google Scholar]
- Spiers, J.M.; Braswell, J.H.; Hegwood, C.P., Jr. Establishment and Maintenance of Rabbiteye Blueberries; Mississippi Agricultural and Forestry Experiment Station in Cooperation with Mississippi Cooperative Extension Service and U.S. Department of Agriculture, Agricultural Research Service: Mississippi State, MS, USA, 1985. [Google Scholar]
- Polashock, J.; Vorsa, N. Cranberry transformation and regeneration. In Transgenic Plants and Crops; Khachatourians, G.G., McHughen, A., Scorza, R., Nip, W.-K., Hui, Y.H., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 2002; pp. 383–396. [Google Scholar]
- Song, G.-Q.; Sink, K.C. Blueberry (Vaccinium corymbosum L.). In Agrobacterium Protocols; Wang, K., Ed.; Springer: New York, NY, USA, 2006; Volume 2, pp. 263–272. [Google Scholar]
- Gaspar, T.; Kevers, C.; Penel, C.; Greppin, H.; Reid, D.M.; Thorpe, T.A. Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell. Dev. Biol. Plant 1996, 32, 272–289. [Google Scholar] [CrossRef]
- Valvekens, D.; Van Montagu, M.; Van Lijsebettens, M. Agrobacterium tumefaciens-mediated transformation of Arabidopsis thaliana root explants by using kanamycin selection. Proc. Natl. Acad. Sci. USA 1988, 85, 5536–5540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.-M.; Schnurr, J.A.; Kapaun, J.A. Timentin as an alternative antibiotic for suppression of Agrobacterium tumefaciens in genetic transformation. Plant Cell Rep. 1998, 17, 646–649. [Google Scholar] [CrossRef] [PubMed]
- Cappelletti, R.; Mezzetti, B. TDZ, 2iP and zeatin in blueberry (Vaccinium corymbosum L. ‘Duke’) in vitro proliferation and organogenesis. Acta Hortic. 2016, 1117, 321–324. [Google Scholar] [CrossRef] [Green Version]
- Reed, B.M.; Abdelnour-Esquivel, A. The Use of Zeatin to Initiate in Vitro Cultures of Vaccinium Species and Cultivars. HortScience 1991, 26, 1320–1322. [Google Scholar] [CrossRef] [Green Version]
- Eccher, T.; Noé, N. Comparison between 2iP and zeatin in the micropropagation of highbush blueberry (Vaccinium corymbosum). Acta Hortic. 1989, 241, 185–190. [Google Scholar] [CrossRef]
- Ružić, D.; Vujović, T.; Libiakova, G.; Cerović, R.; Gajdošova, A. Micropropagation in vitro of highbush blueberry (Vaccinium corymbosum L.). J. Berry Res. 2012, 2, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Sedlak, J.; Paprštein, F. In vitro multiplication of highbush blueberry (Vaccinium corymbosum L.) cultivars. Acta Hortic. 2009, 810, 575–580. [Google Scholar] [CrossRef]
- Kieber, J.J.; Schaller, G.E. Cytokinins. Arab. Book 2014, 12, e0168. [Google Scholar] [CrossRef] [Green Version]
- Von Arnim, A.; Deng, X.W. Light control of seedling development. Annu. Rev. Plant Biol. 1996, 47, 215–243. [Google Scholar] [CrossRef]
- Neff, M.M.; Fankhauser, C.; Chory, J. Light: An indicator of time and place. Genes Dev. 2000, 14, 257–271. [Google Scholar] [PubMed]
- Lloyd, G.; McCown, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Comb. Proc. Int. Plant Propagators Soc. 1980, 30, 421–427. [Google Scholar]
- Rowland, L.J.; Ogden, E.L. Use of a Cytokinin Conjugate for Efficient Shoot Regeneration from Leaf Sections of Highbush Blueberry. HortScience 1992, 27, 1127–1129. [Google Scholar] [CrossRef] [Green Version]
- Song, G.-Q.; Sink, K.C. Agrobacterium tumefaciens-mediated transformation of blueberry (Vaccinium corymbosum L.). Plant Cell Rep. 2004, 23, 475–484. [Google Scholar] [CrossRef]
- Cao, X.; Liu, Q.; Rowland, L.J.; Hammerschlag, F.A. GUS expression in blueberry (Vaccinium spp.): Factors influencing Agrobacterium-mediated gene transfer efficiency. Plant Cell Rep. 1998, 18, 266–270. [Google Scholar] [CrossRef]
- Chetty, V.J.; Ceballos, N.; Garcia, D.; Narváez-Vásquez, J.; Lopez, W.; Orozco-Cárdenas, M.L. Evaluation of four Agrobacterium tumefaciens strains for the genetic transformation of tomato (Solanum lycopersicum L.) cultivar Micro-Tom. Plant Cell Rep. 2013, 32, 239–247. [Google Scholar] [CrossRef]
- Walworth, A.E.; Rowland, L.J.; Polashock, J.J.; Hancock, J.F.; Song, G.-Q. Overexpression of a blueberry-derived CBF gene enhances cold tolerance in a southern highbush blueberry cultivar. Mol. Breed. 2012, 30, 1313–1323. [Google Scholar] [CrossRef]
- Song, G.-Q.; Roggers, R.; Sink, K.; Particka, M.; Zandstra, B. Production of herbicide-resistant highbush blueberry ’legacy’ by Agrobacterium-mediated transformation of the bar gene. Acta Hortic. 2007, 738, 397–407. [Google Scholar] [CrossRef]
- Okada, Y.; Saito, A.; Nishiguchi, M.; Kimura, T.; Mori, M.; Hanada, K.; Sakai, J.; Miyazaki, C.; Matsuda, Y.; Murata, T. Virus resistance in transgenic sweetpotato [Ipomoea batatas L. (Lam)] expressing the coat protein gene of sweet potato feathery mottle virus. Theor. Appl. Genet. 2001, 103, 743–751. [Google Scholar] [CrossRef]
- Figuera Filho, E.S.; Figueiredo, L.F.A.; Monte-Neshich, D.C. Transformation of potato (Solanum tuberosum) cv. Mantiqueira using Agrobacterium tumefaciens and evaluation of herbicide resistance. Plant Cell Rep. 1994, 13, 666–670. [Google Scholar] [CrossRef]
- Spencer, T.M.; Gordon-Kamm, W.J.; Daines, R.J.; Start, W.G.; Lemaux, P.G. Bialaphos selection of stable transformants from maize cell culture. Theor. Appl. Genet. 1990, 79, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Gordon-Kamm, W.J.; Spencer, T.M.; Mangano, M.L.; Adams, T.R.; Daines, R.J.; Start, W.G.; O’Brien, J.V.; Chambers, S.A.; Adams, W.R.; Willetts, N.G.; et al. Transformation of maize cells and regeneration of fertile transgenic plants. Plant Cell 1990, 2, 603–618. [Google Scholar] [PubMed] [Green Version]
- Hauptmann, R.M.; Vasil, V.; Ozias-Akins, P.; Tabaeizadeh, Z.; Rogers, S.G.; Fraley, R.T.; Horsch, R.B.; Vasil, I.K. Evaluation of Selectable Markers for Obtaining Stable Transformants in the Gramineae. Plant Physiol. 1988, 86, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Curtis, I.S.; Power, J.B.; Davey, M.R.; Jones, H. NPTII Assays for measuring gene expression and enzyme activity in transgenic plants. In Plant Gene Transfer and Expression Protocols; Springer: Totowa, NJ, USA, 1995; Volume 49, pp. 149–159. [Google Scholar]
- Shin, Y.-M.; Choe, G.; Shin, B.; Yi, G.; Yun, P.-Y.; Yang, K.; Lee, J.S.; Kwak, S.-S.; Kim, K.-M. Selectionof nptll transgenic sweetpotato plants using G418 and paromomycin. J. Plant Biol. 2007, 50, 206–212. [Google Scholar] [CrossRef]
- Tabei, Y.; Nishio, T.; Kurihara, K.; Kanno, T. Selection of Transformed Callus in a Liquid Medium and Regeneration of Transgenic Plants in Cucumber (Cucumis sativus L.). Jpn. J. Breed. 1994, 44, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Maziah, M.; Sariah, M.; Sreeramanan, S. Transgenic banana Rastali (AAB) with β-1, 3-glucanase gene for tolerance to Fusarium wilt race 1 disease via Agrobacterium-mediated transformation system. Plant Pathol. J. 2007, 6, 271–282. [Google Scholar]
- Collier, R.; Bragg, J.; Hernandez, B.T.; Vogel, J.P.; Thilmony, R. Use of Agrobacterium rhizogenes strain 18r12v and paromomycin selection for transformation of Brachypodium distachyon and Brachypodium sylvaticum. Front. Plant Sci. 2016, 7, 716. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, P.; Hanania, U.; Sahar, N.; Mawassi, M.; Gafny, R.; Sela, I.; Tanne, E.; Perl, A. Improvement of Agrobacterium-mediated transformation efficiency and transgenic plant regeneration of Vitis vinifera L. by optimizing selection regimes and utilizing cryopreserved cell suspensions. Plant Sci. 2005, 168, 565–571. [Google Scholar] [CrossRef]
- Cho, M.-A.; Moon, C.-Y.; Liu, J.-R.; Choi, P.-S. Agrobacterium-mediated transformation in Citrullus lanatus. Biol. Plant. 2008, 52, 365–369. [Google Scholar] [CrossRef]
- Retamales, J.B.; Hancock, J.F. Blueberries, 2nd ed.; Cabi: Wallingford, UK, 2018. [Google Scholar]
- Ehlenfeldt, M.K.; Martin, R.B. A Survey of Fruit Firmness in Highbush Blueberry and Species-introgressed Blueberry Cultivars. HortScience 2002, 37, 386–389. [Google Scholar] [CrossRef] [Green Version]
- Cüce, M.; Bektaş, E.; Sökmen, A. Micropropagation of Vaccinium arctostaphylos L. via lateral-bud culture. Turkish J. Agric. For. 2013, 37, 40–44. [Google Scholar]
- Sathyanarayana, B.N.; Verghese, D.B. Plant Tissue Culture: Practices and New Experimental Protocols; IK International Pvt Ltd.: New Delhi, India, 2007; ISBN 8189866117. [Google Scholar]
- Smith, R.H. Plant Tissue Culture: Techniques and Experiments; Academic Press: College Station, TX, USA, 2012; ISBN 0124159206. [Google Scholar]
- Smith, E.D. The effect of foliar calcium treatments on fruit weight and firmness of rabbiteye blueberry (Vaccinium virgatum Aiton). J. Am. Pomol. Soc. 2016, 70, 74–81. [Google Scholar]
- Blodgett, A.B.; Caldwell, R.W.; McManus, P.S. Effects of Calcium Salts on the Cranberry Fruit Rot Disease Complex. Plant Dis. 2002, 86, 747–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, C.D.; Hong, C.-H.; Kim, S.-K.; Lee, K.-H.; Park, J.-Y.; Nam, M.-W.; Choi, D.-H.; Lee, H.-I. LED light for in vitro and ex vitro efficient growth of economically important highbush blueberry (Vaccinium corymbosum L.). Acta Physiol. Plant. 2016, 38, 1–9. [Google Scholar] [CrossRef]
- Heo, J.W.; Shin, K.S.; Kim, S.K.; Paek, K.Y. Light quality affectsin Vitro growth of grape ‘Teleki 5BB’. J. Plant Biol. 2006, 49, 276–280. [Google Scholar] [CrossRef]
- Norberto, S.; Silva, S.; Meireles, M.; Faria, A.C.; Pintado, M.; Calhau, C. Blueberry anthocyanins in health promotion: A metabolic overview. J. Funct. Foods 2013, 5, 1518–1528. [Google Scholar] [CrossRef]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent Research on the Health Benefits of Blueberries and Their Anthocyanins. Adv. Nutr. 2020, 11, 224–236. [Google Scholar] [CrossRef]
- González-Villagra, J.; Reyes-Díaz, M.; Alberdi, M.; Acevedo, P.; Loyola, R.; Tighe-Neira, R.; Arce-Johnson, P.; Inostroza-Blancheteau, C. Solar UV irradiation effects on photosynthetic performance, biochemical markers, and gene expression in highbush blueberry (Vaccinium corymbosum L.) cultivars. Sci. Hortic. 2020, 259, 108816. [Google Scholar] [CrossRef]
- Perkins-Veazie, P.; Collins, J.K.; Howard, L. Blueberry fruit response to postharvest application of ultraviolet radiation. Postharvest Biol. Technol. 2008, 47, 280–285. [Google Scholar] [CrossRef]
- Dias, M.I.; Sousa, M.J.; Alves, R.C.; Ferreira, I.C.F.R. Exploring plant tissue culture to improve the production of phenolic compounds: A review. Ind. Crop. Prod. 2016, 82, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Song, G.-Q.; Sink, K.C.; Callow, P.W.; Baughan, R.; Hancock, J.F. Evaluation of a Herbicide-resistant Trait Conferred by the Bar Gene Driven by Four Distinct Promoters in Transgenic Blueberry Plants. J. Am. Soc. Hortic. Sci. 2008, 133, 605–611. [Google Scholar] [CrossRef] [Green Version]
- Ciaburro, G.; Venkateswaran, B. Neural Networks with R: Smart Models Using CNN, RNN, Deep Learning, and Artificial Intelligence Principles; Packt Publishing Ltd.: Birmingham, UK, 2017; ISBN 1788399412. [Google Scholar]
- Hesami, M.; Naderi, R.; Yoosefzadeh-Najafabadi, M.; Rahmati, M. Data-driven modeling in plant tissue culture. J. Appl. Environ. Biol. Sci. 2017, 7, 37–44. [Google Scholar]
- Hesami, M.; Condori-Apfata, J.A.; Valencia, M.V.; Mohammadi, M. Application of Artificial Neural Network for Modeling and Studying In Vitro Genotype-Independent Shoot Regeneration in Wheat. Appl. Sci. 2020, 10, 5370. [Google Scholar] [CrossRef]
- Hesami, M.; Naderi, R.; Tohidfar, M.; Yoosefzadeh-Najafabadi, M. Development of support vector machine-based model and comparative analysis with artificial neural network for modeling the plant tissue culture procedures: Effect of plant growth regulators on somatic embryogenesis of chrysanthemum, as a case study. Plant Methods 2020, 16, 1–15. [Google Scholar] [CrossRef]
- Hesami, M.; Naderi, R.; Tohidfar, M. Modeling and Optimizing Medium Composition for Shoot Regeneration of Chrysanthemum via Radial Basis Function-Non-dominated Sorting Genetic Algorithm-II (RBF-NSGAII). Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kaur, P.; Gupta, R.C.; Dey, A.; Malik, T.; Pandey, D.K. Optimization of salicylic acid and chitosan treatment for bitter secoiridoid and xanthone glycosides production in shoot cultures of Swertia paniculata using response surface methodology and artificial neural network. BMC Plant Biol. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- García-Pérez, P.; Lozano-Milo, E.; Landín, M.; Gallego, P.P. Combining Medicinal Plant In Vitro Culture with Machine Learning Technologies for Maximizing the Production of Phenolic Compounds. Antioxidants 2020, 9, 210. [Google Scholar] [CrossRef] [Green Version]
- Arab, M.M.; Yadollahi, A.; Eftekhari, M.; Ahmadi, H.; Akbari, M.; Khorami, S.S. Modeling and Optimizing a New Culture Medium for In Vitro Rooting of G×N15 Prunus Rootstock using Artificial Neural Network-Genetic Algorithm. Sci. Rep. 2018, 8, 1–18. [Google Scholar] [CrossRef]
- Arab, M.M.; Yadollahi, A.; Shojaeiyan, A.; Ahmadi, H. Artificial Neural Network Genetic Algorithm as Powerful Tool to Predict and Optimize In vitro Proliferation Mineral Medium for G × N15 Rootstock. Front. Plant Sci. 2016, 7, 1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Deng, D.; Dai, W.; Li, J.; Jin, X. Optimization of culture conditions for differentiation of melon based on artificial neural network and genetic algorithm. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https//www.R-project.org (accessed on 8 August 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Dordrecht, The Netherlands, 2016. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappai, F.; Garcia, A.; Cullen, R.; Davis, M.; Munoz, P.R. Advancements in Low-Chill Blueberry Vaccinium corymbosum L. Tissue Culture Practices. Plants 2020, 9, 1624. https://doi.org/10.3390/plants9111624
Cappai F, Garcia A, Cullen R, Davis M, Munoz PR. Advancements in Low-Chill Blueberry Vaccinium corymbosum L. Tissue Culture Practices. Plants. 2020; 9(11):1624. https://doi.org/10.3390/plants9111624
Chicago/Turabian StyleCappai, Francesco, Alexandria Garcia, Ryan Cullen, Matthew Davis, and Patricio R. Munoz. 2020. "Advancements in Low-Chill Blueberry Vaccinium corymbosum L. Tissue Culture Practices" Plants 9, no. 11: 1624. https://doi.org/10.3390/plants9111624