Effect of Adjuvants on Herbicidal Activity and Selectivity of Three Phytotoxins Produced by the Fungus, Stagonospora cirsii



Abstract
:1. Introduction
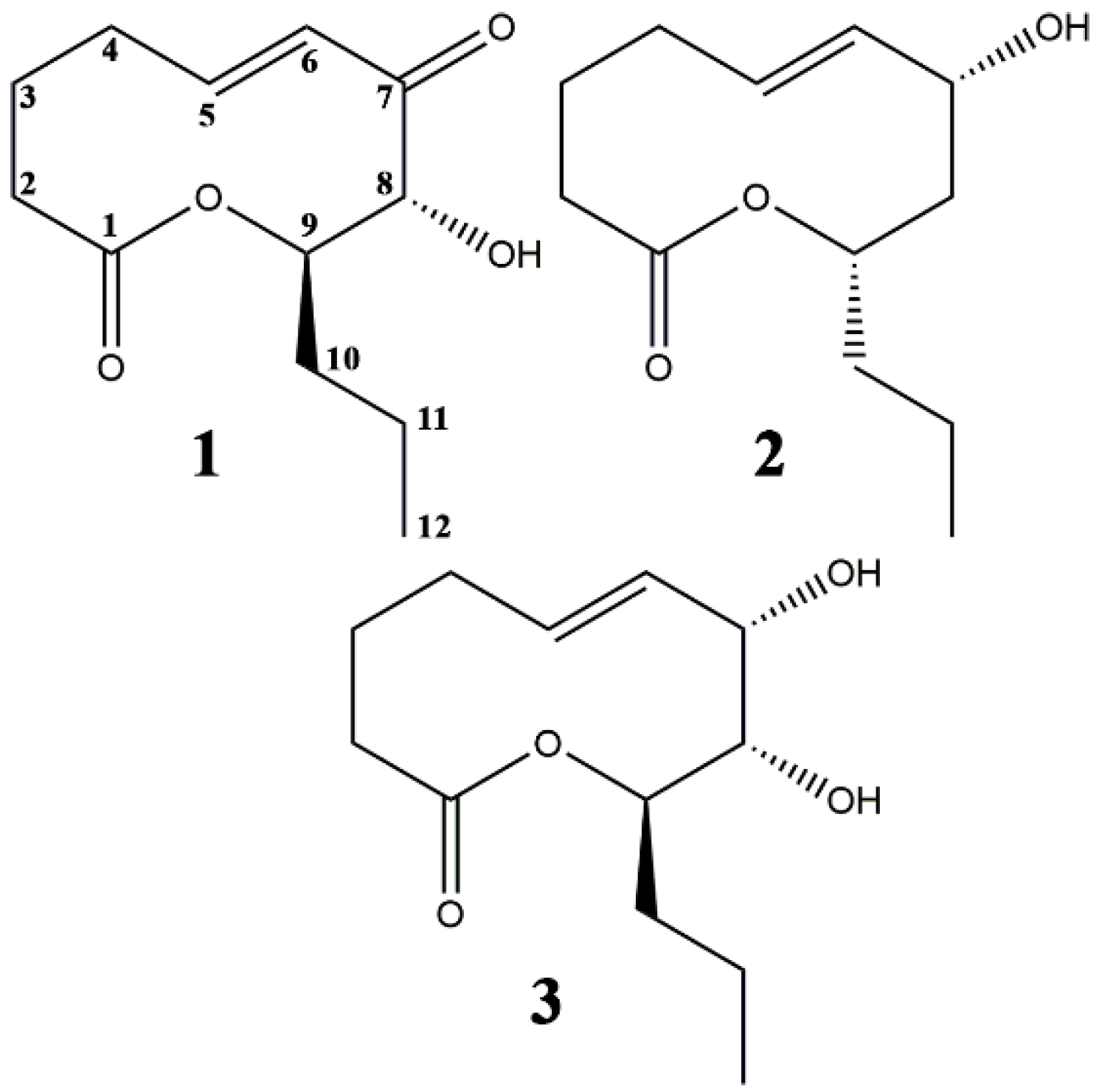
2. Results
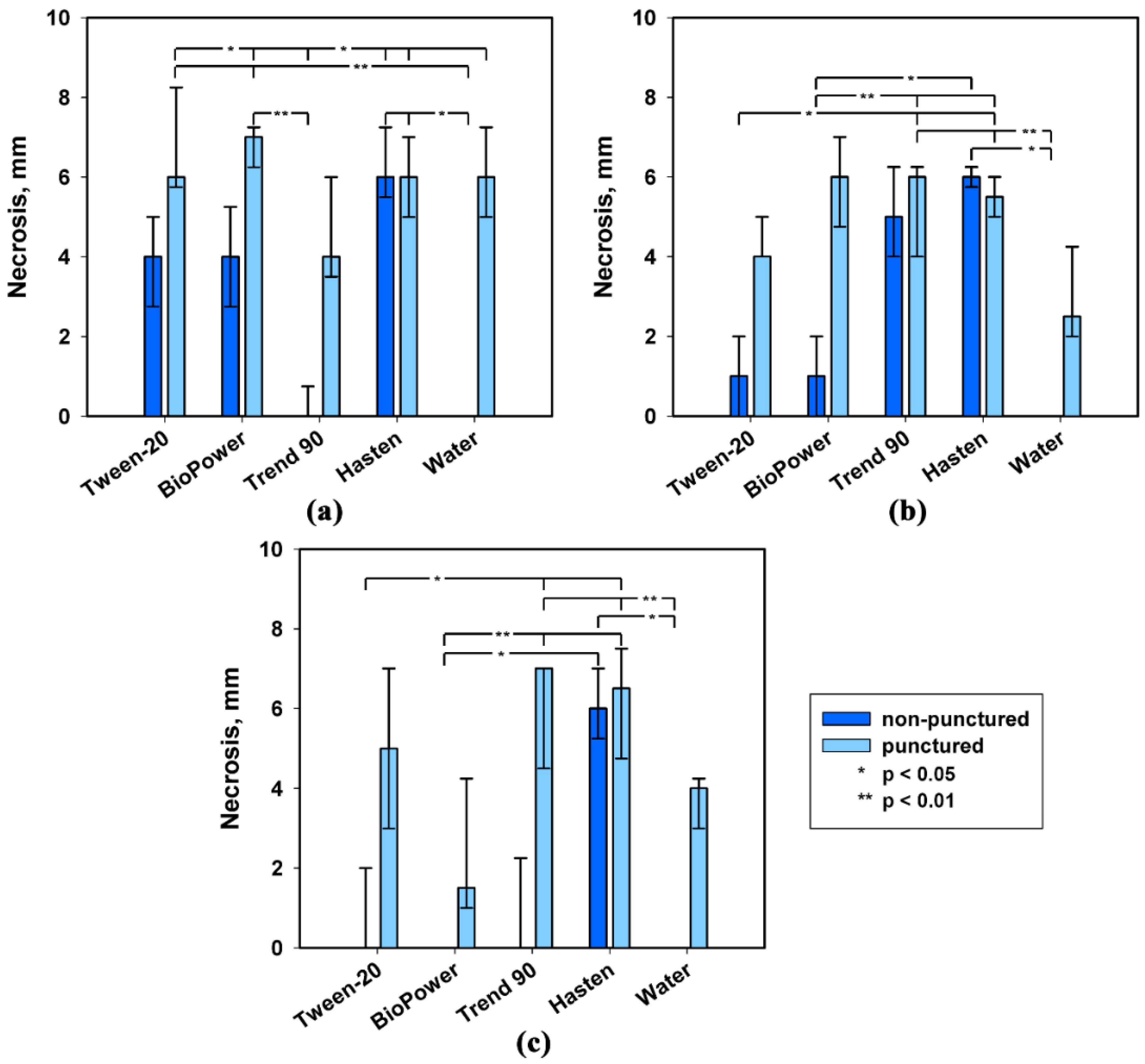
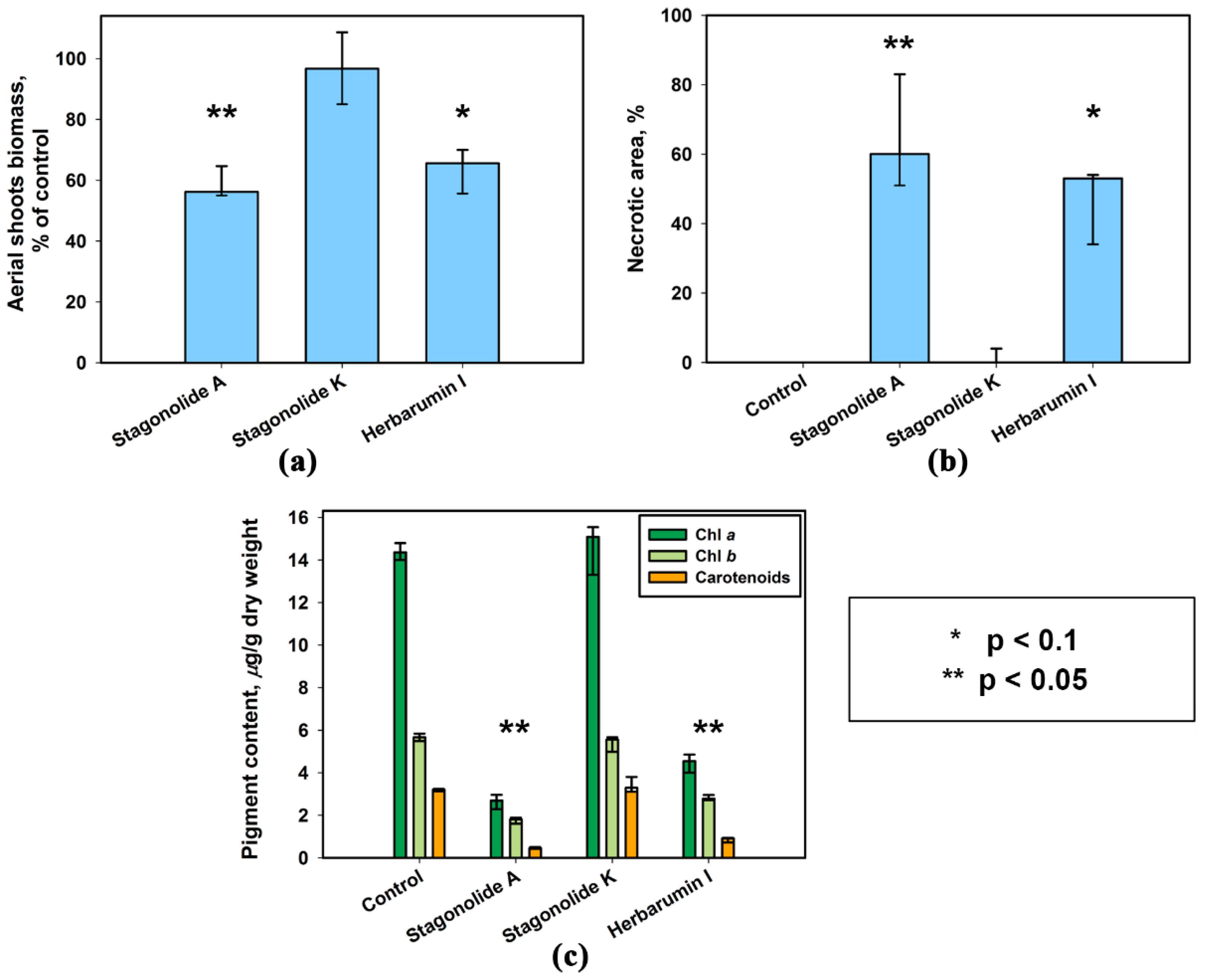
2.1. Effect of Leaf Damage and Adjuvants on Phytotoxicity of S. cirsii Toxins
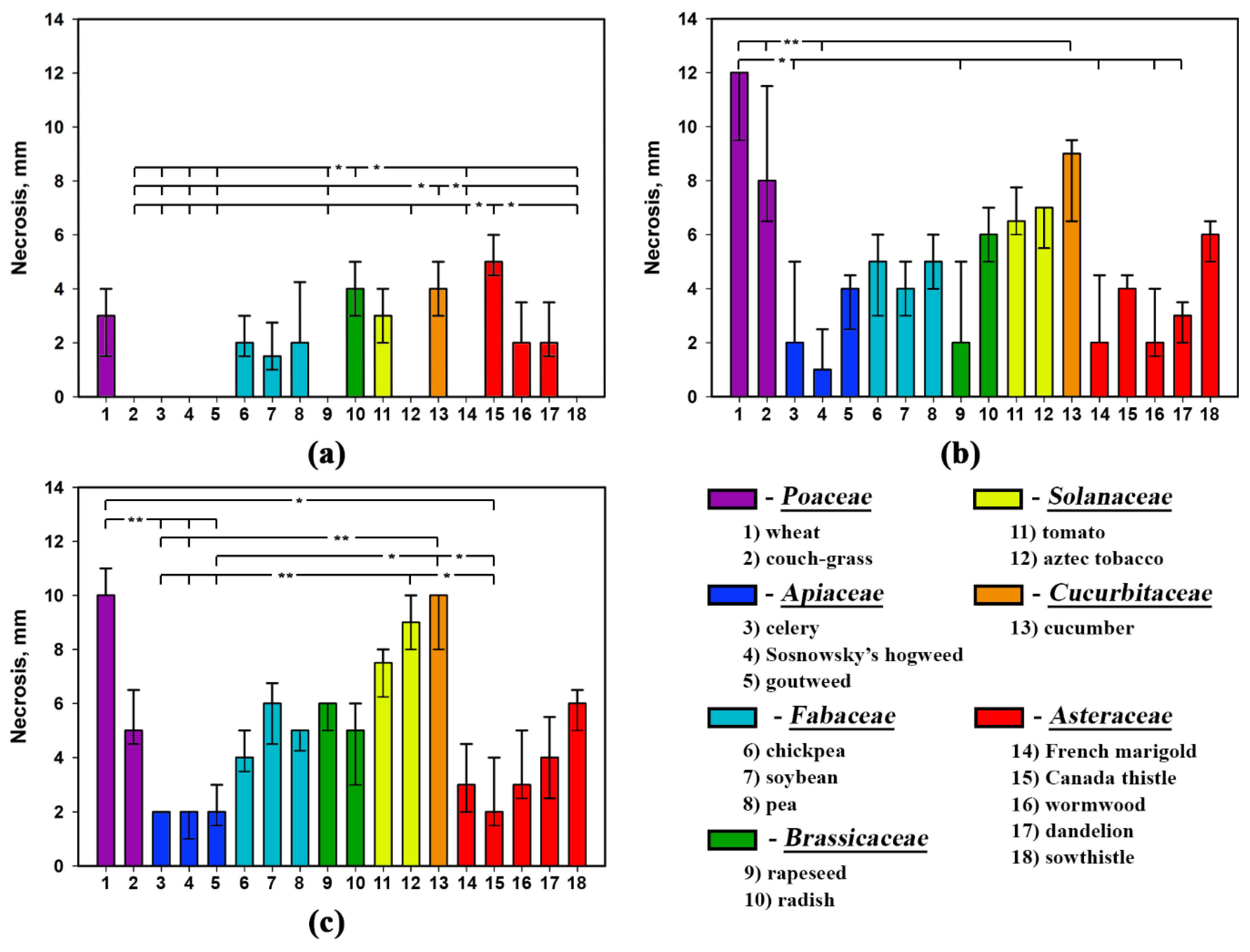
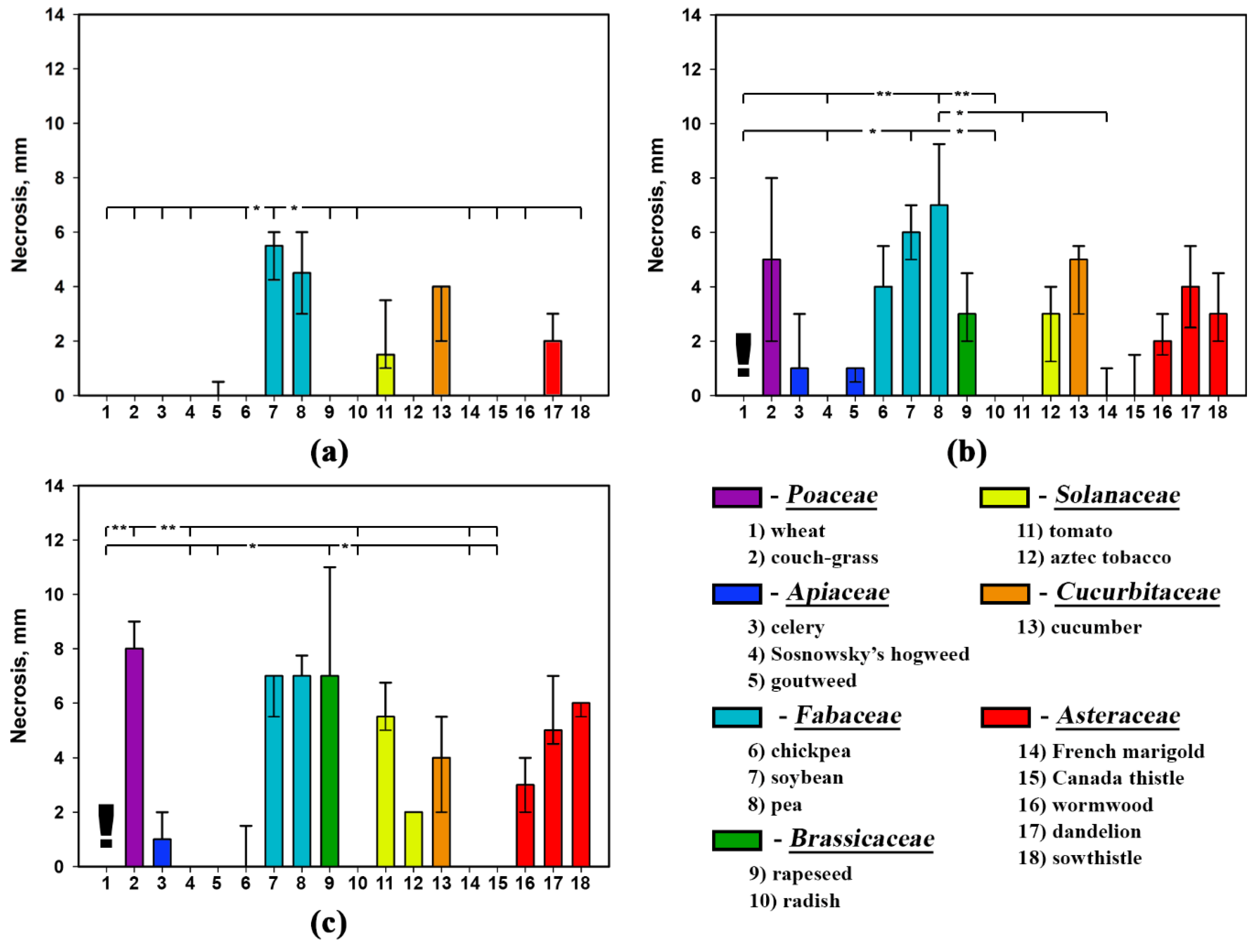
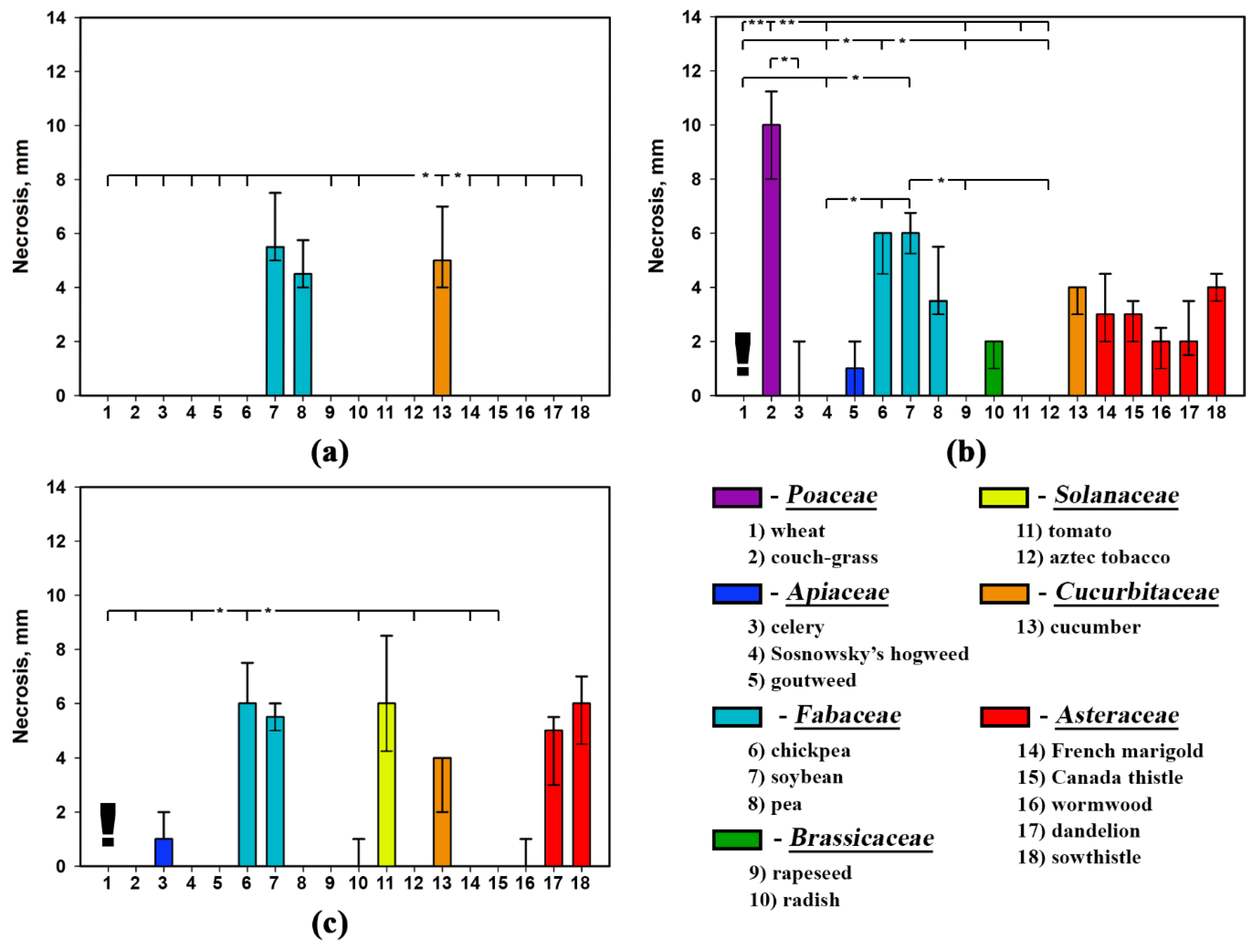
2.2. Phytotoxic Selectivity of S. cirsii Toxins
2.3. Contact Herbicidal Activity of S. cirsii Phytotoxins
3. Discussion
4. Materials and Methods
4.1. Fungal Strain and Toxin Production
4.2. Effect of Adjuvants on Phytotoxic Activity
4.3. Phytotoxic Selectivity of S. cirsii Toxins
4.4. Herbicidal Activity of Phytotoxins
4.5. Quantification of Photosynthetic Pigments
4.6. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Westwood, J.H.; Charudattan, R.; Duke, S.O.; Fennimore, S.A.; Marrone, P.; Slaughter, D.C.; Swanton, C.; Zollinger, R. Weed management in 2050: Perspectives on the future of weed science. Weed Sci. 2018, 66, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Glare, T.; Caradus, J.; Gelernter, W.; Jackson, T.; Keyhani, N.; Köhl, J.; Marrone, P.; Morin, L.; Stewart, A. Have biopesticides come of age? Trends Biotechnol. 2012, 30, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O.; Owens, D.K.; Dayan, F.E. Natural Product-Based Chemical Herbicides. In Weed Control: Sustainability, Hazards, and Risks in Cropping Systems Worldwide, 1st ed.; Korres, N.E., Burgos, N.R., Duke, S.O., Eds.; CRC Press: Boca Raton, FL, USA, 2019; Chapter 8; pp. 153–165. [Google Scholar] [CrossRef]
- Duke, S.O.; Dayan, F.E. Discovery of New Herbicide Modes of Action with Natural Phytotoxins. In Discovery and Synthesis of Crop Protection Products; Maienfisch, P., Stevenson, T.M., Eds.; ACS Symposium Series eBooks: Washington, DC, USA, 2015; pp. 79–92. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Cutignano, A.; Taglialatela-Scafati, O.; Vurro, M.; Chiara Zonno, M.; Motta, A. Ascaulitoxin, a phytotoxic bis-amino acid N-glucoside from Ascochyta caulina. Phytochemistry 1998, 48, 1131–1137. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Evidente, M.; Evidente, A. Fungal phytotoxins with potential herbicidal activity to control Chenopodium album. Nat. Prod. Commun. 2015, 10, 1119–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graupner, P.R.; Carr, A.; Clancy, E.; Gilbert, J.; Bailey, K.L.; Derby, J.A.; Gerwick, B.C. The Macrocidins: Novel cyclic tetramic acids with herbicidal activity produced by Phoma macrostoma. J. Nat. Prod. 2003, 66, 1558–1561. [Google Scholar] [CrossRef]
- Hubbard, M.; Hynes, R.K.; Bailey, K.L. Impact of macrocidins, produced by Phoma macrostoma, on carotenoid profiles of plants. Biol. Control 2015, 89, 11–22. [Google Scholar] [CrossRef]
- Hubbard, M.; Taylor, W.G.; Bailey, K.L.; Hynes, R.K. The dominant modes of action of macrocidins, bioherbicidal metabolites of Phoma macrostoma, differ between susceptible plant species. Environ. Exp. Bot. 2016, 132, 80–91. [Google Scholar] [CrossRef]
- Zhou, B.; Wang, H.; Meng, B.; Wei, R.; An, C.; Chen, S.; Yang, C.; Qiang, S. An evaluation of tenuazonic acid, a potential biobased herbicide in cotton. Pest. Manag. Sci. 2019, 75, 2482–2489. [Google Scholar] [CrossRef]
- Chen, S.; Qiang, S. Recent advances in tenuazonic acid as a potential herbicide. Pestic. Biochem. Phys. 2017, 143, 252–257. [Google Scholar] [CrossRef]
- Duke, S.O.; Cantrell, C.L.; Meepagala, K.M.; Wedge, D.E.; Tabanca, N.; Schrader, K.K. Natural toxins for use in pest management. Toxins 2010, 2, 1943–1962. [Google Scholar] [CrossRef] [Green Version]
- Dayan, F.E.; Owens, D.K.; Duke, S.O. Rationale for a natural products approach to herbicide discovery. Pest Manag. Sci. 2012, 68, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Kastanias, M.A.; Chrysayi-Tokousbalides, M. Herbicidal potential of pyrenophorol isolated from a Drechslera avenae pathotype. Pest Manag. Sci. 2000, 56, 227–232. [Google Scholar] [CrossRef]
- Jiang, S.J.; Qiang, S.; Zhu, Y.Z.; Dong, Y.F. Isolation and phytotoxicity of a metabolite from Curvularia eragrostidis and characterisation of its modes of action. Ann. Appl. Biol. 2008, 152, 103–111. [Google Scholar] [CrossRef]
- Hignett, R.C.; Quirk, A.V. Properties of phytotoxic cell-wall components of plant pathogenic pseudomonads. J. Gen. Microbiol. 1979, 110, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Frías, M.; Brito, N.; González, M.; González, C. The phytotoxic activity of the cerato-platanin BcSpl1 resides in a two-peptide motif on the protein surface. Mol. Plant Pathol. 2013, 15, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Freda, F.; Sangermano, F.; Calabrò, V.; Cimmino, A.; Cristofaro, M.; Meyer, S.; Evidente, A. radicinin, a fungal phytotoxin as a target-specific bioherbicide for invasive buffelgrass (Cenchrus ciliaris) control. Molecules 2019, 24, 1086. [Google Scholar] [CrossRef] [Green Version]
- Qiang, S.; Wang, L.; Wei, R.; Zhou, B.; Chen, S.; Zhu, Y.; Dong, Y.; An, C. Bioassay of the herbicidal activity of AAC-toxin produced by Alternaria alternata isolated from Ageratina adenophora. Weed Technol. 2010, 24, 197–201. [Google Scholar] [CrossRef]
- Zollinger, R.K. Influence of adjuvants on weed control from tribenuron. J. ASTM Int. 2005, 2, 1–7. [Google Scholar] [CrossRef]
- Lockett, J.; Morgan, C. Herbicide Composition. EP 1981339B1, 29 February 2012. Available online: https://patents.google.com/patent/EP1981339B1/zh-cn (accessed on 22 October 2018).
- McWhorter, C.G. The Physiological Effects of Adjuvants on Plants. In Weed Physiology. Volume II: Herbicide Physiology, 1st ed.; Duke, S.O., Ed.; CRC Press: Boca Raton, FL, USA, 2017; Chapter 6; pp. 141–155. [Google Scholar] [CrossRef]
- McDaniel, G.L.; Klingeman, W.E.; Witte, W.T.; Flanagan, P.C. Choice of adjuvant with halosulfuron affects purple nutsedge control and nursery crop tolerance. Hort. Sci. 2001, 36, 1085–1088. [Google Scholar] [CrossRef] [Green Version]
- Frihauf, J.C.; Miller, S.D.; Alford, C.M. Imazamox Rates, Timings, and Adjuvants Affect Imidazolinone-tolerant winter wheat cultivars. Weed Technol. 2005, 19, 599–607. [Google Scholar] [CrossRef]
- Pacanoski, Z. Herbicides and Adjuvants. In Herbicides, Physiology of Action, and Safety; Price, A., Kelton, J., Sarunaite, L., Eds.; IntechOpen: Rijeka, Croatia, 2015; pp. 125–148. [Google Scholar] [CrossRef] [Green Version]
- Yuzikhin, O.; Mitina, G.; Berestetskiy, A. Herbicidal potential of stagonolide, a new phytotoxic nonenolide from Stagonospora cirsii. J. Agric. Food Chem. 2007, 55, 7707–7711. [Google Scholar] [CrossRef] [PubMed]
- Fausto Rivero-Cruz, J.; García-Aguirre, G.; Cerda-García-Rojas, C.M.; Mata, R. Conformational behavior and absolute stereostructure of two phytotoxic nonenolides from the fungus Phoma herbarum. Tetrahedron 2000, 56, 5337–5344. [Google Scholar] [CrossRef]
- Evidente, A.; Lanzetta, R.; Capasso, R.; Vurro, M.; Botralico, A. Pinolidoxin, a phytotoxic nonenolide from Ascochyta pinodes. Phytochemistry 1993, 34, 999–1003. [Google Scholar] [CrossRef]
- Evidente, A.; Lanzetta, R.; Capasso, R.; Andolfi, A.; Botralico, A.; Vurro, M.; Zonno, M.C. Putaminoxin, a phytotoxic nonenolide from Phoma putaminum. Phytochemistry 1995, 40, 1637–1641. [Google Scholar] [CrossRef]
- Evidente, A.; Cimmino, A.; Berestetskiy, A.; Mitina, G.; Andolfi, A.; Motta, A. Stagonolides B-F, nonenolides produced by Stagonospora cirsii, a potential mycoherbicide of Cirsium arvense. J. Nat. Prod. 2008, 71, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Cimmino, A.; Berestetskiy, A.; Andolfi, A.; Motta, A. Stagonolides G-I and Modiolide A, nonenolides produced by Stagonospora cirsii, a potential mycoherbicide for Cirsium arvense. J. Nat. Prod. 2008, 71, 1897–1901. [Google Scholar] [CrossRef]
- Dalinova, A.; Dubovik, V.; Chisty, L.; Kochura, D.; Ivanov, A.; Smirnov, S.; Petrova, M.; Zolatarev, A.; Evidente, A.; Berestetskiy, A. Stagonolides J and K and Stagochromene A, two new natural substituted nonenolides and a new disubstituted chromene-4,5-dione isolated from Stagonospora cirsii S-47 proposed for the biocontrol of Sonchus arvensis. J. Agric. Food Chem. 2019, 67, 13040–13050. [Google Scholar] [CrossRef]
- Holloway, P.J.; Butler Ellis, M.C.; Webb, D.A.; Western, N.M.; Tuck, C.R.; Hayes, A.L.; Miller, P.C. Effects of some agricultural tank-mix adjuvants on the deposition efficiency of aqueous sprays on foliage. Crop Prot. 2000, 19, 27–37. [Google Scholar] [CrossRef]
- Green, J.M.; Beestman, G.B. Recently patented and commercialized formulation and adjuvant technology. Crop Prot. 2007, 26, 320–327. [Google Scholar] [CrossRef]
- Poluektova, E.; Tokarev, Y.; Sokornova, S.; Chisty, L.; Evidente, A.; Berestetskiy, A. Curvulin and Phaeosphaeride A from Paraphoma sp. VIZR 1.46 isolated from Cirsium arvense as Pote##ntial H##erbicides. Molecules 2018, 23, 2795. [Google Scholar] [CrossRef] [Green Version]
- Somervaille, A.; Betts, G.; Gordon, B.; Green, V.; Burgis, M.; Henderson, R. Adjuvants—Oils, Surfactants and Other Additives for Farm Chemicals—Revised 2012 Edition; GRDC: Barton, Australia, 2012; pp. 1–48. [Google Scholar]
- Qiang, S.; Dong, Y.; An, C.; Zhou, B.; Zhu, Y.; Chen, S.; Dai, X.; Dai, B.; Cai, J. Biological control of weeds using the metabolites of Alternaria Alternata. EP 1849361A1, 2008. Available online: https://patents.google.com/patent/EP1849361A1/en (accessed on 19 October 2020).
- Nalewaja, J.D. Oils as and with herbicides. In Spray Oils Beyond 2000. Sustainable Pest and Disease Management, 1st ed.; Beattie, G.A.C., Watson, D.M., Stevens, M.L., Rae, D.J., Spooner-Hart, R.N., Eds.; Univ. Western: Sydney, Australia, 2002; pp. 290–300. [Google Scholar]
- Cimmino, A.; Andolfi, A.; Fondevilla, S.; Abouzeid, M.A.; Rubiales, D.; Evidente, A. Pinolide, a new nonenolide produced by Didymella pinodes, the causal agent of ascochyta blight on Pisum sativum. J. Agric. Food Chem. 2012, 60, 5273–5278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robeson, D.J.; Strobel, G.A. Zinniol induces chlorophyll retention in barley leaves: The selective action of a nonhost-specific phytotoxin. Phytochemistry 1984, 23, 1597–1599. [Google Scholar] [CrossRef]
- Durbin, R.D.; Uchytil, T.F. A survey of plant insensitivity to tentoxin. Phytopathology 1977, 67, 602–603. [Google Scholar] [CrossRef]
- Chow, P.N.P.; Grant, C.A.; Hinshalwood, A.M. Adjuvants and Agrochemicals. Volume 1: Mode of Action and Physiological Activity, 1st ed.; CRC Press: Boca Raton, FL, USA, 1989; p. 222. [Google Scholar] [CrossRef]
- Foy, C.L. Adjuvants-current trends and technology. In Pesticide Formulation and Adjuvant Technology; Foy, C.L., Pritchard, D.W., Eds.; CRC Press: Boca Raton, FL, USA, 1996; pp. 323–352. [Google Scholar] [CrossRef]
- Li, J.; Chen, W.; Xu, Y.; Wu, X. Comparative effects of different types of tank-mixed adjuvants on the efficacy, absorption and translocation of cyhalofop-butyl in barnyardgrass (Echinochloa crus-galli [L.] Beauv.). Weed Biol. Manag. 2016, 16, 80–89. [Google Scholar] [CrossRef]
- Xiong, Z.T. Bioaccumulation and physiological effects of excess lead in a roadside pioneer species Sonchus oleraceus L. Environ. Pollut. 1997, 97, 275–279. [Google Scholar] [CrossRef]
- El-Khatib, A.A. The response of some common Egyptian plants to ozone and their use as biomonitors. Environ. Pollut. 2003, 124, 419–428. [Google Scholar] [CrossRef]
- Fraga, M.I.; Tasende, M.G. Mechanisms of resistance to simazine in Sonchus oleraceus. Weed Res. 2003, 43, 333–340. [Google Scholar] [CrossRef]
- Jia, P.Y.; Hu, Y.; Zhang, L.X.; Ze, H.; Tian, F.P.; Hu, Y.; Wu, G.L. Physiological characteristics of three wild Sonchus species to prolonged drought tolerance in arid regions. Pak. J. Bot. 2018, 50, 9–17. [Google Scholar]
- Halloin, J.M.; de Zoeten, G.A.; Gaard, G.; Walker, J.C. The effects of tentoxin on chlorophyll synthesis and plastid structure in cucumber and cabbage. Plant Physiol. 1970, 45, 310–314. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Strasser, R.J.; Qiang, S. In vivo assessment of effect of phytotoxin tenuazonic acid on PSII reaction centers. Plant Physiol. Biochem. 2014, 84, 10–21. [Google Scholar] [CrossRef]
- Kshirsagar, A.; Reid, A.J.; McColl, S.M.; Saunders, V.A.; Whalley, A.J.S.; Evans, E.H. The effect of fungal metabolites on leaves as detected by chlorophyll fluorescence. New Phytol. 2001, 151, 451–457. [Google Scholar] [CrossRef]
- Kastanias, M.A.; Chrysayi-Tokousbalides, M. Bioactivity of the fungal metabolite (8r,16r)-(-)-pyrenophorin on Graminaceous Plants. J. Agric. Food Chem. 2005, 53, 5943–5947. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-S.; Bao, W.; Liu, D.-Y.; Ling, N.; Ying, R.-R.; Raza, W.; Shen, Q.-R. Effect of fusaric acid on biomass and photosynthesis of watermelon seedlings leaves. Caryologia 2008, 61, 258–268. [Google Scholar] [CrossRef]
- Berestetskiy, A.; Dmitriev, A.; Mitina, G.; Lisker, I.; Andolfi, A.; Evidente, A. Nonenolides and cytochalasins with phytotoxic activity against Cirsium arvense and Sonchus arvensis: A structure–activity relationships study. Phytochemistry 2008, 69, 953–960. [Google Scholar] [CrossRef]
- Duke, S.O.; Dayan, F.E. Modes of action of microbially-produced phytotoxins. Toxins 2011, 3, 1038–1064. [Google Scholar] [CrossRef] [Green Version]
- Berestetskiy, A.O.; Yuzikhin, O.S.; Katkova, A.S.; Dobrodumov, A.V.; Sivogrivov, D.E.; Kolombet, L.V. Isolation, identification, and characteristics of the phytotoxin produced by the fungus Alternaria cirsinoxia. Appl. Biochem. Microbiol. 2010, 46, 75–79. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Extraction of photosynthetic tissues: Chlorophylls and carotenoids. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.2.1–F4.2.6. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Wounding | Phytotoxin | ||
|---|---|---|---|
| Stagonolide A | Stagonolide K | Herbarumin I | |
| punctured | H = 6.34, p = 0.175 | H = 12.58, p = 0.014 * | H = 9.76, p = 0.045 * |
| non-punctured | H = 16.81, p = 0.002 * | H = 22.85, p = 0.000 * | H = 21.15, p = 0.000 * |
| Adjuvant | Phytotoxin | ||
|---|---|---|---|
| Stagonolide A | Stagonolide K | Herbarumin I | |
| Tween®-20 | H = 7.37, p = 0.007 * | H = 4.95, p = 0.026 * | H = 6.53, p = 0.012 * |
| BioPower® | H = 6.02, p = 0.014 * | H = 6.88, p = 0.009 * | H = 3.58, p = 0.059 |
| Trend® 90 | H = 2.80, p = 0.094 | H = 0.11, p = 0.742 | H = 8.47, p = 0.004 * |
| Hasten™ | H = 0.11, p = 0.737 | H = 1.88, p = 0.171 | H = 0.06, p = 0.803 |
| Water | H = 9.54, p = 0.002 * | H = 7.21, p = 0.007 * | H = 9.66, p = 0.001 * |
| Leaf Wounding | Adjuvant | ||||
|---|---|---|---|---|---|
| Tween®-20 | BioPower® | Trend® 90 | Hasten™ | Water | |
| punctured | H = 5.78, p = 0.056 | H = 10.28, p = 0.006 * | H = 2.25, p = 0.325 | H = 1.43, p = 0.490 | H = 11.19, p = 0.037 * |
| non-punctured | H = 6.47, p = 0.039 * | H = 8.72, p = 0.013 * | H = 9.21, p = 0.010 * | H = 0.26, p = 0.878 | H = 0.00, p = 1.000 |
| Formulation/Wounding | Phytotoxin | ||
|---|---|---|---|
| Stagonolide A | Stagonolide K | Herbarumin I | |
| Water/ non-punctured | H = 77.61, p = 0.000 * | H = 82.39, p = 0.000 * | H = 82.73, p = 0.000 * |
| Water/ punctured | H = 67.55, p = 0.000 * | H = 73.33, p = 0.000 * | H = 77.34, p = 0.000 * |
| 0.1% Hasten™/ non-punctured | H = 71.01, p = 0.000 * | H = 82.40, p = 0.000 * | H = 80.43, p = 0.000 * |
| Plant Species | Phytotoxin | ||
|---|---|---|---|
| Stagonolide A | Stagonolide K | Herbarumin I | |
| wheat | H = 14.60, p = 0.002 * | H = 0.00, p = 1.000 | H = 0.00, p = 1.000 |
| couch-grass | H = 15.41, p = 0.002 * | H = 12.63, p = 0.006 * | H = 16.00, p = 0.001 * |
| celery | H = 15.59, p = 0.001 * | H = 16.52, p = 0.001 * | H = 15.26, p = 0.002 * |
| Sosnowsky’s hogweed | H = 12.55, p = 0.006 * | H = 0.00, p = 1.000 | H = 0.00, p = 1.000 |
| goutweed | H = 14.48, p = 0.002 * | H = 6.65, p = 0.084 | H = 12.44, p = 0.006 * |
| chickpea | H = 8.36, p = 0.039 * | H = 15.71, p = 0.001 * | H = 14.89, p = 0.002 * |
| soybean | H = 12.39, p = 0.006 * | H = 4.69, p = 0.196 | H = 1.67, p = 0.644 |
| pea | H = 4.00, p = 0.261 | H = 8.57, p = 0.036 * | H = 12.46, p = 0.006 * |
| rapeseed | H = 15.16, p = 0.002 * | H = 16.76, p = 0.001 * | H = 6.32, p = 0.097 |
| radish | H = 9.51, p = 0.023 * | H = 0.00, p = 1.000 | H = 11.29, p = 0.010 * |
| tomato | H = 5.91, p = 0.116 | H = 13.45, p = 0.004 * | H = 13.26, p = 0.004 * |
| aztec tobacco | H = 17.15, p = 0.001 * | H = 12.55, p = 0.006 * | H = 18.55, p = 0.000 * |
| cucumber | H = 9.93, p = 0.019 * | H = 6.17, p = 0.104 | H = 12.42, p = 0.006 * |
| French marigold | H = 10.97, p = 0.012 * | H = 4.75, p = 0.191 | H = 16.75, p = 0.001 * |
| Canada thistle | H = 7.70, p = 0.053 | H = 14.35, p = 0.003 * | H = 16.65, p = 0.001 * |
| wormwood | H = 10.44, p = 0.015 * | H = 15.64, p = 0.001 * | H = 15.65, p = 0.001 * |
| dandelion | H = 3.60, p = 0.309 | H = 5.98, p = 0.112 | H = 13.80, p = 0.003 * |
| perennial sowthistle | H = 11.53, p = 0.009 * | H = 16.62, p = 0.001 * | H = 14.94, p = 0.002 * |
| Plant Species | Formulation/Wounding | ||
|---|---|---|---|
| Water/ Non-Punctured | Water/ Punctured | 0.1% Hasten™/ Non-Punctured | |
| wheat | H = 13.32, p = 0.001 * | H = 13.43, p = 0.001 * | H = 13.46, p = 0.001 * |
| couch-grass | H = 10.22, p = 0.006 * | H = 5.58, p = 0.061 | H = 13.23, p = 0.001 * |
| celery | H = 0.00, p = 1.000 | H = 5.41, p = 0.067 | H = 4.67, p = 0.097 |
| Sosnowsky’s hogweed | H = 0.00, p = 1.000 | H = 13.43, p = 0.001 * | H = 13.46, p = 0.001 * |
| goutweed | H = 2.00, p = 0.368 | H = 9.20, p = 0.010 * | H = 13.43, p = 0.001 * |
| chickpea | H = 13.36, p = 0.001 * | H = 1.07, p = 0.585 | H = 12.73, p = 0.002 * |
| soybean | H = 7.83, p = 0.020 * | H = 5.37, p = 0.068 | H = 2.59, p = 0.274 |
| pea | H = 2.14, p = 0.342 | H = 8.38, p = 0.015 * | H = 10.51, p = 0.005 * |
| rapeseed | H = 0.00, p = 1.000 | H = 10.18, p = 0.006 * | H = 13.21, p = 0.001 * |
| radish | H = 12.63, p = 0.002 * | H = 13.13, p = 0.001 * | H = 11.69, p = 0.003 * |
| tomato | H = 9.40, p = 0.009 * | H = 10.51, p = 0.005 * | H = 2.36, p = 0.307 |
| aztec tobacco | H = 0.00, p = 1.000 | H = 13.08, p = 0.001 * | H = 13.51, p = 0.001 * |
| cucumber | H = 5.03, p = 0.080 | H = 9.75, p = 0.008 * | H = 10.00, p = 0.007 * |
| French marigold | H = 2.00, p = 0.368 | H = 9.90, p = 0.007 * | H = 13.32, p = 0.001 * |
| Canada thistle | H = 13.43, p = 0.001 * | H = 11.66, p = 0.003 * | H = 13.32, p = 0.001 * |
| wormwood | H = 13.43, p = 0.001 * | H = 0.89, p = 0.640 | H = 9.80, p = 0.008 * |
| dandelion | H = 10.22, p = 0.006 * | H = 3.54, p = 0.170 | H = 2.62, p = 0.270 |
| sowthistle | H = 0.00, p = 1.000 | H = 8.80, p = 0.012 * | H = 0.64, p = 0.725 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubovik, V.; Dalinova, A.; Berestetskiy, A. Effect of Adjuvants on Herbicidal Activity and Selectivity of Three Phytotoxins Produced by the Fungus, Stagonospora cirsii. Plants 2020, 9, 1621. https://doi.org/10.3390/plants9111621
Dubovik V, Dalinova A, Berestetskiy A. Effect of Adjuvants on Herbicidal Activity and Selectivity of Three Phytotoxins Produced by the Fungus, Stagonospora cirsii. Plants. 2020; 9(11):1621. https://doi.org/10.3390/plants9111621
Chicago/Turabian StyleDubovik, Vsevolod, Anna Dalinova, and Alexander Berestetskiy. 2020. "Effect of Adjuvants on Herbicidal Activity and Selectivity of Three Phytotoxins Produced by the Fungus, Stagonospora cirsii" Plants 9, no. 11: 1621. https://doi.org/10.3390/plants9111621