LHCSR3-Type NPQ Prevents Photoinhibition and Slowed Growth under Fluctuating Light in Chlamydomonas reinhardtii


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
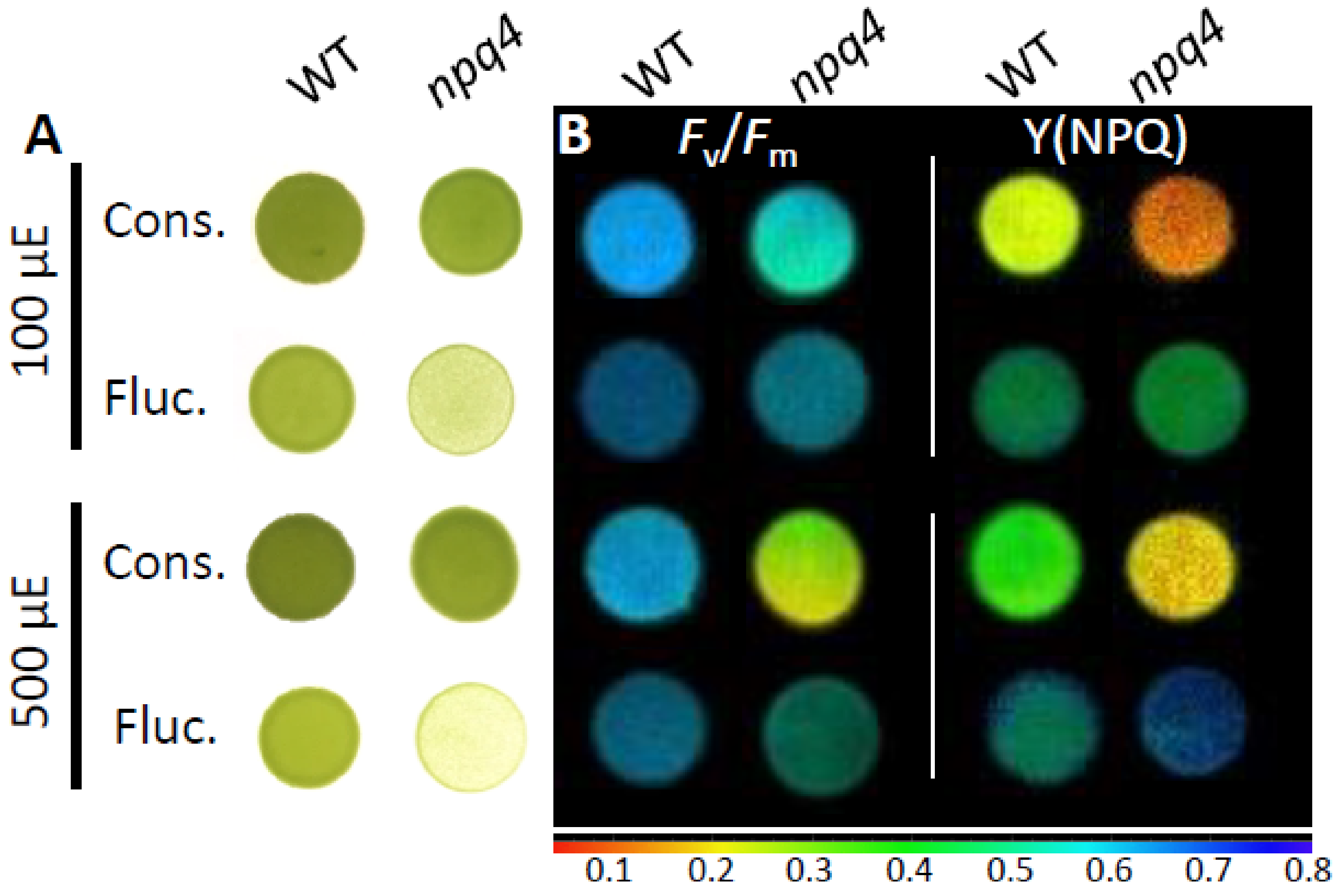
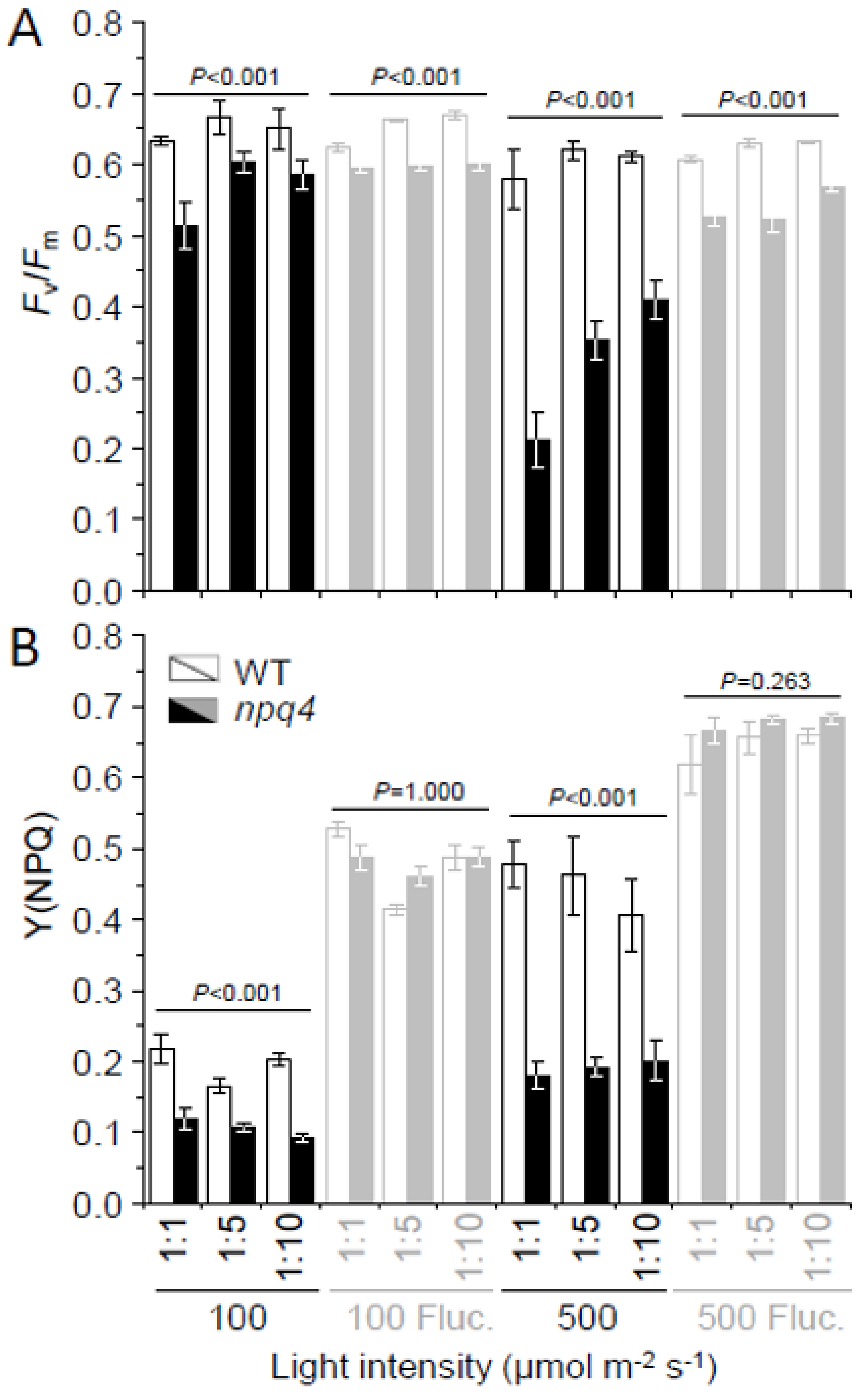
2.1. Light Fluctuations, but Not Constant Light, Slowed npq4 Growth
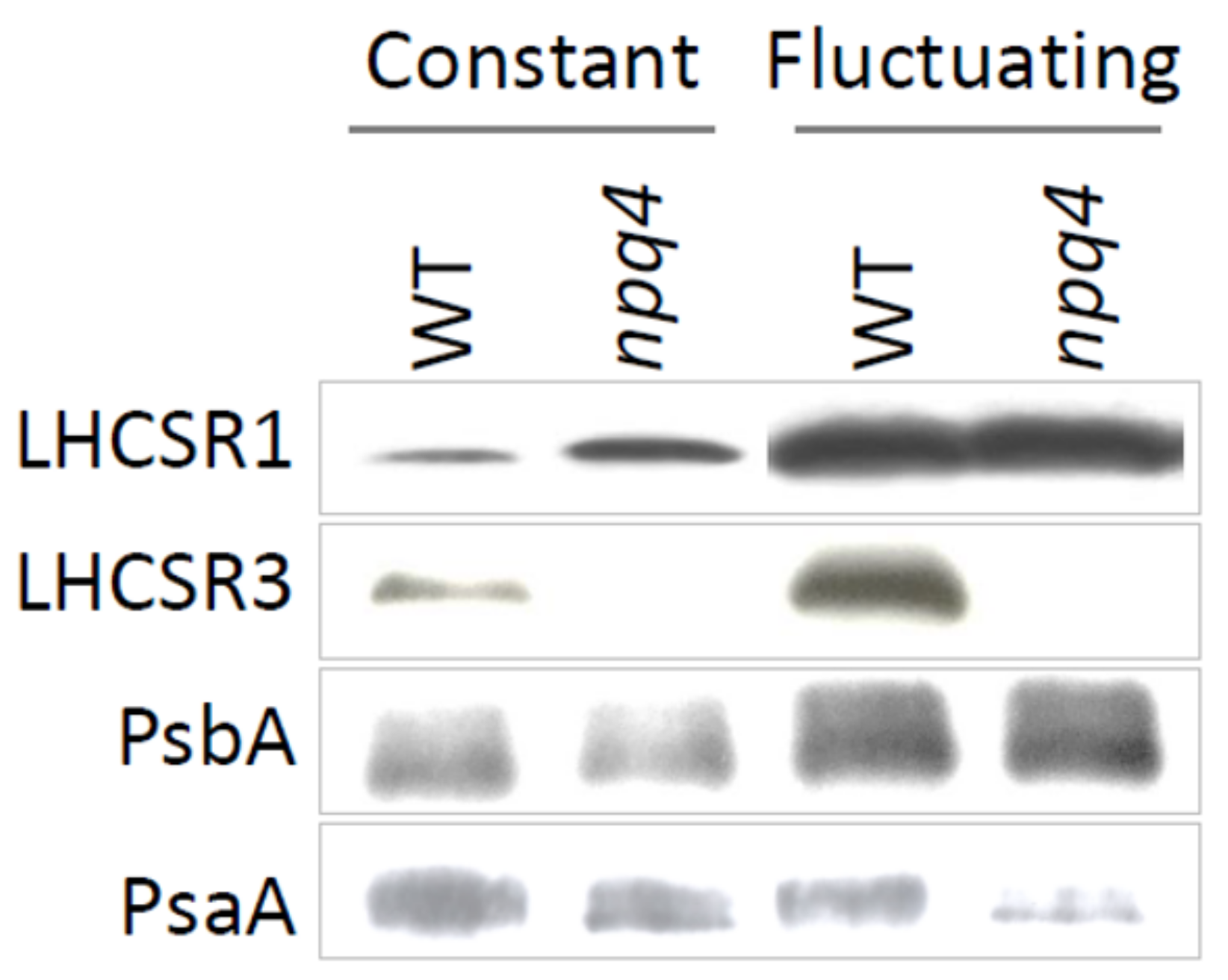
2.2. Light Fluctuations Induced Accumulation of LHCSR1, LHCSR3, and qE Capacity
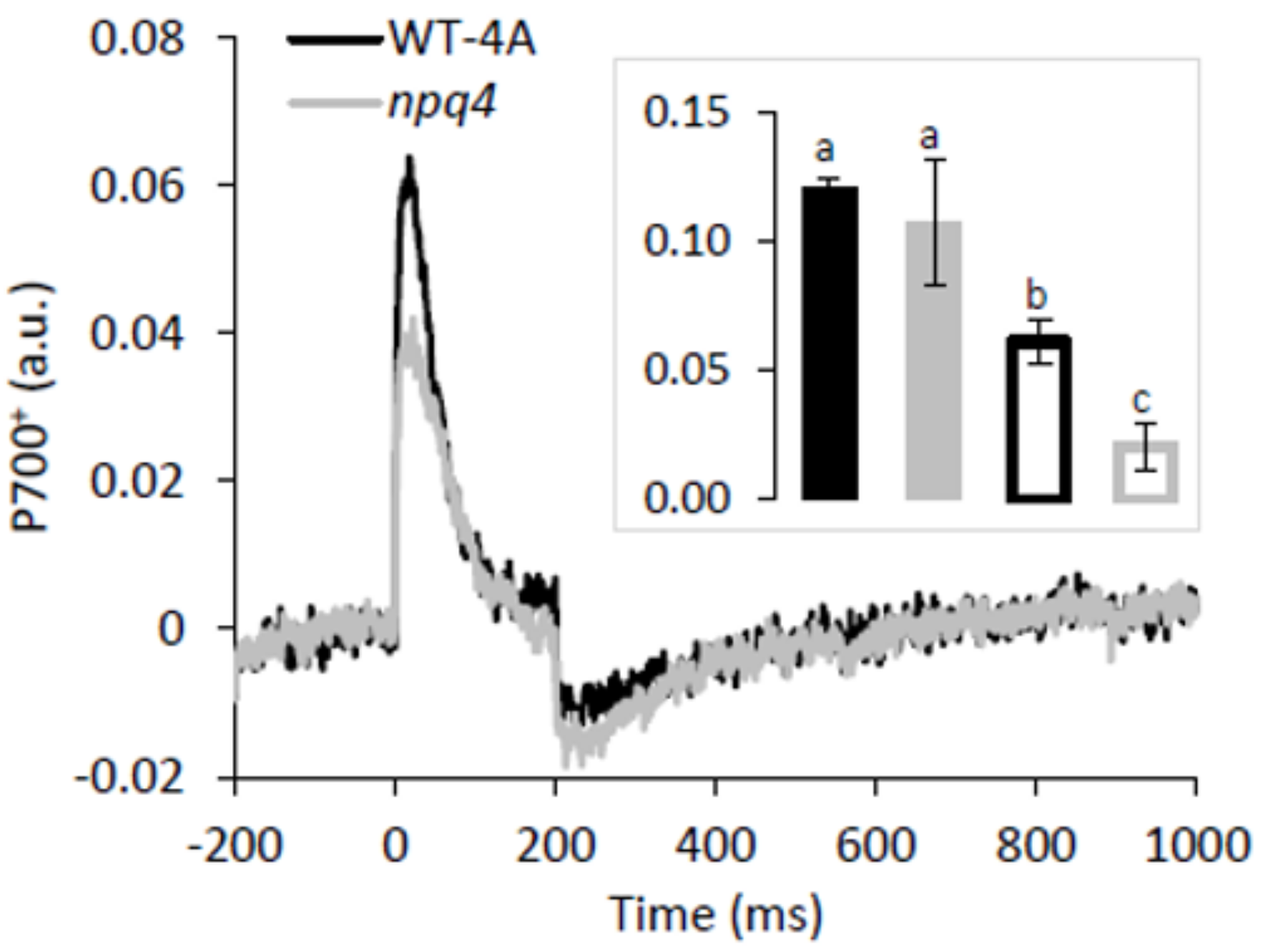
2.3. Enhanced PSII and PSI Photoinhibition Occurred in npq4 Under Constant and Fluctuating Light, Respectively
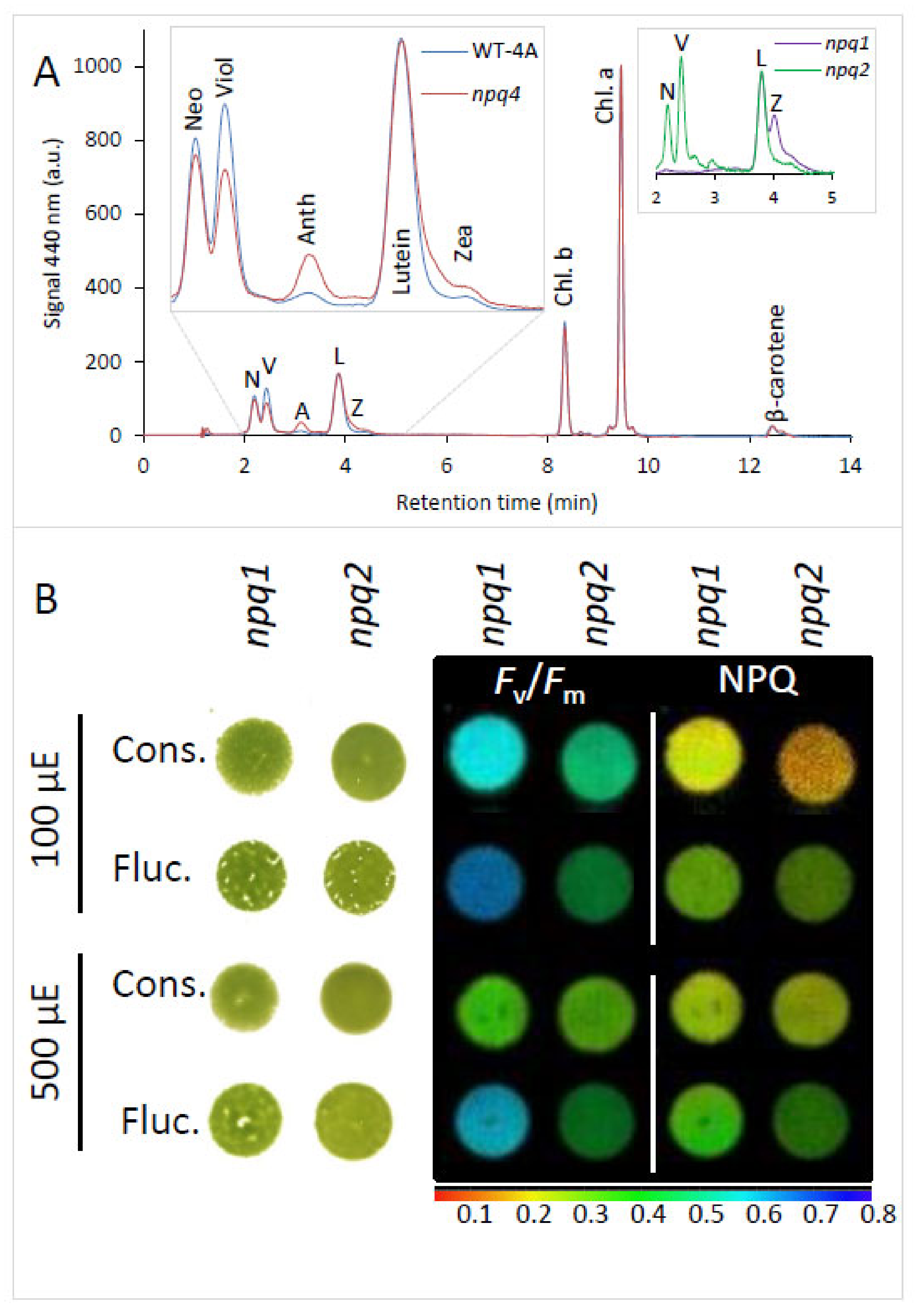
2.4. Xanthophyll Composition Did Not Affect Tolerance to Fluctuating Light
3. Discussion
4. Conclusions
5. Material and Methods
5.1. C. reinhardtii Strains and Growth Conditions
5.2. Chlorophyll Fluorescence
5.3. Western Blotting of Proteins
5.4. HPLC of Photosynthetic Pigments
5.5. Maximum Photo-Oxidizable P700 Level
5.6. Statistics
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Allahverdiyeva, Y.; Suorsa, M.; Tikkanen, M.; Aro, E.M. Photoprotection of photosystems in fluctuating light intensities. J. Exp. Bot. 2015, 66, 2427–2436. [Google Scholar] [CrossRef]
- Alboresi, A.; Storti, M.; Morosinotto, T. Balancing protection and efficiency in the regulation of photosynthetic electron transport across plant evolution. New Phytol. 2019, 221, 105–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, P.; Li, X.-P.; Niyogi, K.K. Update on photosynthesis non-photochemical quenching. A response to excess light energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roach, T.; Krieger-Liszkay, A. Photosynthetic regulatory mechanisms for efficiency and prevention of photo-oxidative stress. Ann. Plant Rev. Online 2019, 273–306. [Google Scholar] [CrossRef]
- Erickson, E.; Wakao, S.; Niyogi, K.K. Light stress and photoprotection in Chlamydomonas reinhardtii. Plant J. 2015, 82, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-P.; Gilmore, A.M.; Caffarri, S.; Bassi, R.; Golan, T.; Kramer, D.; Niyogi, K.K. Regulation of photosynthetic light harvesting involves intrathylakoid lumen pH sensing by the PsbS protein. J. Biol. Chem. 2004, 279, 22866–22874. [Google Scholar] [CrossRef] [Green Version]
- Liguori, N.; Roy, L.M.; Opacic, M.; Durand, G.; Croce, R. Regulation of light harvesting in the green alga Chlamydomonas reinhardtii: The c-terminus of LHCSR is the knob of a dimmer switch. J. Am. Chem. Soc. 2013, 135, 18339–18342. [Google Scholar] [CrossRef]
- Ballottari, M.; Truong, T.B.; De Re, E.; Erickson, E.; Stella, G.R.; Fleming, G.R.; Bassi, R.; Niyogi, K.K. Identification of pH-sensing sites in the light harvesting complex stress-related 3 protein essential for triggering non-photochemical quenching in Chlamydomonas reinhardtii. J. Biol. Chem. 2016, 291, 7334–7346. [Google Scholar] [CrossRef] [Green Version]
- Niyogi, K.K.; Truong, T.B. Evolution of flexible non-photochemical quenching mechanisms that regulate light harvesting in oxygenic photosynthesis. Curr. Opin. Plant Biol. 2013, 16, 307–314. [Google Scholar] [CrossRef]
- Roach, T.; Na, C.S.; Stöggl, W.; Krieger-Liszkay, A. The non-photochemical quenching protein LHCSR3 prevents oxygen-dependent photoinhibition in Chlamydomonas reinhardtii. J. Exp. Bot. 2020, 71, 2650–2660. [Google Scholar] [CrossRef]
- Roach, T.; Baur, T.; Stoggl, W.; Krieger-Liszkay, A. Chlamydomonas reinhardtii responding to high light: A role for 2-propenal (acrolein). Physiol. Plant. 2017, 161, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Polukhina, I.; Fristedt, R.; Dinc, E.; Cardol, P.; Croce, R. Carbon supply and photoacclimation crosstalk in the green alga Chlamydomonas reinhardtii. Plant Physiol. 2016, 172, 1494–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peers, G.; Truong, T.B.; Ostendorf, E.; Busch, A.; Elrad, D.; Grossman, A.R.; Hippler, M.; Niyogi, K.K. An ancient light-harvesting protein is critical for the regulation of algal photosynthesis. Nature 2009, 462, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Girolomoni, L.; Cazzaniga, S.; Pinnola, A.; Perozeni, F.; Ballottari, M.; Bassi, R. LHCSR3 is a nonphotochemical quencher of both photosystems in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2019, 116, 4212–4217. [Google Scholar] [CrossRef] [Green Version]
- Dinc, E.; Tian, L.; Roy, L.M.; Roth, R.; Goodenough, U.; Croce, R. LHCSR1 induces a fast and reversible ph-dependent fluorescence quenching in LHCII in Chlamydomonas reinhardtii cells. Proc. Natl. Acad. Sci. USA 2016, 113, 7673–7678. [Google Scholar] [CrossRef] [Green Version]
- Kosuge, K.; Tokutsu, R.; Kim, E.; Akimoto, S.; Yokono, M.; Ueno, Y.; Minagawa, J. LHCSR1-dependent fluorescence quenching is mediated by excitation energy transfer from LHCII to photosystem I in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2018, 115, 3722–3727. [Google Scholar] [CrossRef] [Green Version]
- Tokutsu, R.; Minagawa, J. Energy-dissipative supercomplex of photosystem II associated with LHCSR3 in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2013, 110, 10016–10021. [Google Scholar] [CrossRef] [Green Version]
- Semchonok, D.A.; Yadav, K.N.S.; Xu, P.Q.; Drop, B.; Croce, R.; Boekema, E.J. Interaction between the photoprotective protein LHCSR3 and c2s2 photosystem II supercomplex in Chlamydomonas reinhardtii. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 379–385. [Google Scholar] [CrossRef]
- Quaas, T.; Berteotti, S.; Ballottari, M.; Flieger, K.; Bassi, R.; Wilhelm, C.; Goss, R. Non-photochemical quenching and xanthophyll cycle activities in six green algal species suggest mechanistic differences in the process of excess energy dissipation. J. Plant Physiol. 2015, 172, 92–103. [Google Scholar] [CrossRef]
- Tian, L.J.; Nawrocki, W.J.; Liu, X.; Polukhina, I.; van Stokkum, I.H.M.; Croce, R. pH dependence, kinetics and light-harvesting regulation of nonphotochemical quenching in Chlamydomonas. Proc. Natl. Acad. Sci. USA 2019, 116, 8320–8325. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Plazaola, J.I.; Esteban, R.; Fernandez-Marin, B.; Kranner, I.; Porcar-Castell, A. Thermal energy dissipation and xanthophyll cycles beyond the arabidopsis model. Photosynth. Res. 2012, 113, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enamala, M.K.; Enamala, S.; Chavali, M.; Donepudi, J.; Yadavalli, R.; Kolapalli, B.; Aradhyula, T.V.; Velpuri, J.; Kuppam, C. Production of biofuels from microalgae—A review on cultivation, harvesting, lipid extraction, and numerous applications of microalgae. Renew. Sustain. Energy Rev. 2018, 94, 49–68. [Google Scholar] [CrossRef]
- Chaux, F.; Burlacot, A.; Mekhalfi, M.; Auroy, P.; Blangy, S.; Richaud, P.; Peltier, G. Flavodiiron proteins promote fast and transient o2 photoreduction in Chlamydomonas. Plant Physiol. 2017, 174, 1825–1836. [Google Scholar] [CrossRef] [Green Version]
- Jokel, M.; Johnson, X.; Peltier, G.; Aro, E.M.; Allahverdiyeva, Y. Hunting the main player enabling Chlamydomonas reinhardtii growth under fluctuating light. Plant J. 2018, 94, 822–835. [Google Scholar] [CrossRef] [Green Version]
- Lepetit, B.; Gelin, G.; Lepetit, M.; Sturm, S.; Vugrinec, S.; Rogato, A.; Kroth, P.G.; Falciatore, A.; Lavaud, J. The diatom phaeodactylum tricornutum adjusts nonphotochemical fluorescence quenching capacity in response to dynamic light via fine-tuned lhcx and xanthophyll cycle pigment synthesis. New Phytol. 2017, 214, 205–218. [Google Scholar] [CrossRef]
- Niyogi, K.K.; Bjorkman, O.; Grossman, A.R. Chlamydomonas xanthophyll cycle mutants identified by video imaging of chlorophyll fluorescence quenching. Plant Cell 1997, 9, 1369–1380. [Google Scholar] [CrossRef] [Green Version]
- Roach, T.; Krieger-Liszkay, A. Regulation of photosynthetic electron transport and photoinhibition. Curr. Protein Pept. Sci. 2014, 15, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Roach, T.; Krieger-Liszkay, A. The role of the PsbS protein in the protection of photosystems I and II against high light in Arabidopsis thaliana. BBA Bioenerg. 2012, 1817, 2158–2165. [Google Scholar] [CrossRef] [Green Version]
- Cantrell, M.; Peers, G. A mutant of Chlamydomonas without LHCSR maintains high rates of photosynthesis, but has reduced cell division rates in sinusoidal light conditions. PLoS ONE 2017, 12, e0179395. [Google Scholar] [CrossRef] [Green Version]
- Masojídek, J.; Kopecká, J.; Koblížek, M.; Torzillo, G. The xanthophyll cycle in green algae (chlorophyta): Its role in the photosynthetic apparatus. Plant Biol. 2004, 6, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.A.; Kanazawa, A.; Schottler, M.A.; Kohzuma, K.; Froehlich, J.E.; Rutherford, A.W.; Satoh-Cruz, M.; Minhas, D.; Tietz, S.; Dhingra, A.; et al. Limitations to photosynthesis by proton motive force-induced photosystem II photodamage. eLife 2016, 5, e16921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suorsa, M.; Jarvi, S.; Grieco, M.; Nurmi, M.; Pietrzykowska, M.; Rantala, M.; Kangasjarvi, S.; Paakkarinen, V.; Tikkanen, M.; Jansson, S.; et al. Proton gradient regulation5 is essential for proper acclimation of arabidopsis photosystem I to naturally and artificially fluctuating light conditions. Plant Cell 2012, 24, 2934–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergner, S.V.; Scholz, M.; Trompelt, K.; Barth, J.; Gabelein, P.; Steinbeck, J.; Xue, H.; Clowez, S.; Fucile, G.; Goldschmidt-Clermont, M.; et al. State transition7-dependent phosphorylation is modulated by changing environmental conditions, and its absence triggers remodeling of photosynthetic protein complexes. Plant Physiol. 2015, 168, 615–634. [Google Scholar] [CrossRef] [Green Version]
- Chaux, F.; Johnson, X.; Auroy, P.; Beyly-Adriano, A.; Te, I.; Cuine, S.; Peltier, G. Pgrl1 and LHCSR3 compensate for each other in controlling photosynthesis and avoiding photosystem I photoinhibition during high light acclimation of Chlamydomonas cells. Mol. Plant 2017, 10, 216–218. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Aro, E.-M.; Millar, A.H. Mechanisms of photodamage and protein turnover in photoinhibition. Trends Plant Sci. 2018, 23, 667–676. [Google Scholar] [CrossRef]
- Depege, N.; Bellafiore, S.; Rochaix, J.D. Role of chloroplast protein kinase stt7 in LHCII phosphorylation and state transition in Chlamydomonas. Science 2003, 299, 1572–1575. [Google Scholar] [CrossRef] [Green Version]
- Roach, T.; Na, C.S. LHCSR3 affects de-coupling and re-coupling of LHCII to PSII during state transitions in Chlamydomonas reinhardtii. Sci. Rep. 2017, 7, 43145. [Google Scholar] [CrossRef] [Green Version]
- Bennoun, P. The present model for chlororespiration. Photosynth. Res. 2002, 73, 273–277. [Google Scholar] [CrossRef]
- Allorent, G.; Tokutsu, R.; Roach, T.; Peers, G.; Cardol, P.; Girard-Bascou, J.; Seigneurin-Berny, D.; Petroutsos, D.; Kuntz, M.; Breyton, C.; et al. A dual strategy to cope with high light in Chlamydomonas reinhardtii. Plant Cell 2013, 25, 545–557. [Google Scholar] [CrossRef] [Green Version]
- Goldschmidt-Clermont, M.; Bassi, R. Sharing light between two photosystems: Mechanism of state transitions. Curr. Opin. Plant Biol. 2015, 25, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquot, J.-P. Dark deactivation of chloroplast enzymes finally comes to light. Proc. Natl. Acad. Sci. USA 2018, 115, 9334–9335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonoike, K. Photoinhibition of photosystem I. Physiol. Plant. 2011, 142, 56–64. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roach, T. LHCSR3-Type NPQ Prevents Photoinhibition and Slowed Growth under Fluctuating Light in Chlamydomonas reinhardtii. Plants 2020, 9, 1604. https://doi.org/10.3390/plants9111604
Roach T. LHCSR3-Type NPQ Prevents Photoinhibition and Slowed Growth under Fluctuating Light in Chlamydomonas reinhardtii. Plants. 2020; 9(11):1604. https://doi.org/10.3390/plants9111604
Chicago/Turabian StyleRoach, Thomas. 2020. "LHCSR3-Type NPQ Prevents Photoinhibition and Slowed Growth under Fluctuating Light in Chlamydomonas reinhardtii" Plants 9, no. 11: 1604. https://doi.org/10.3390/plants9111604