Disruption of Very-Long-Chain-Fatty Acid Synthesis Has an Impact on the Dynamics of Cellulose Synthase in Arabidopsis thaliana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
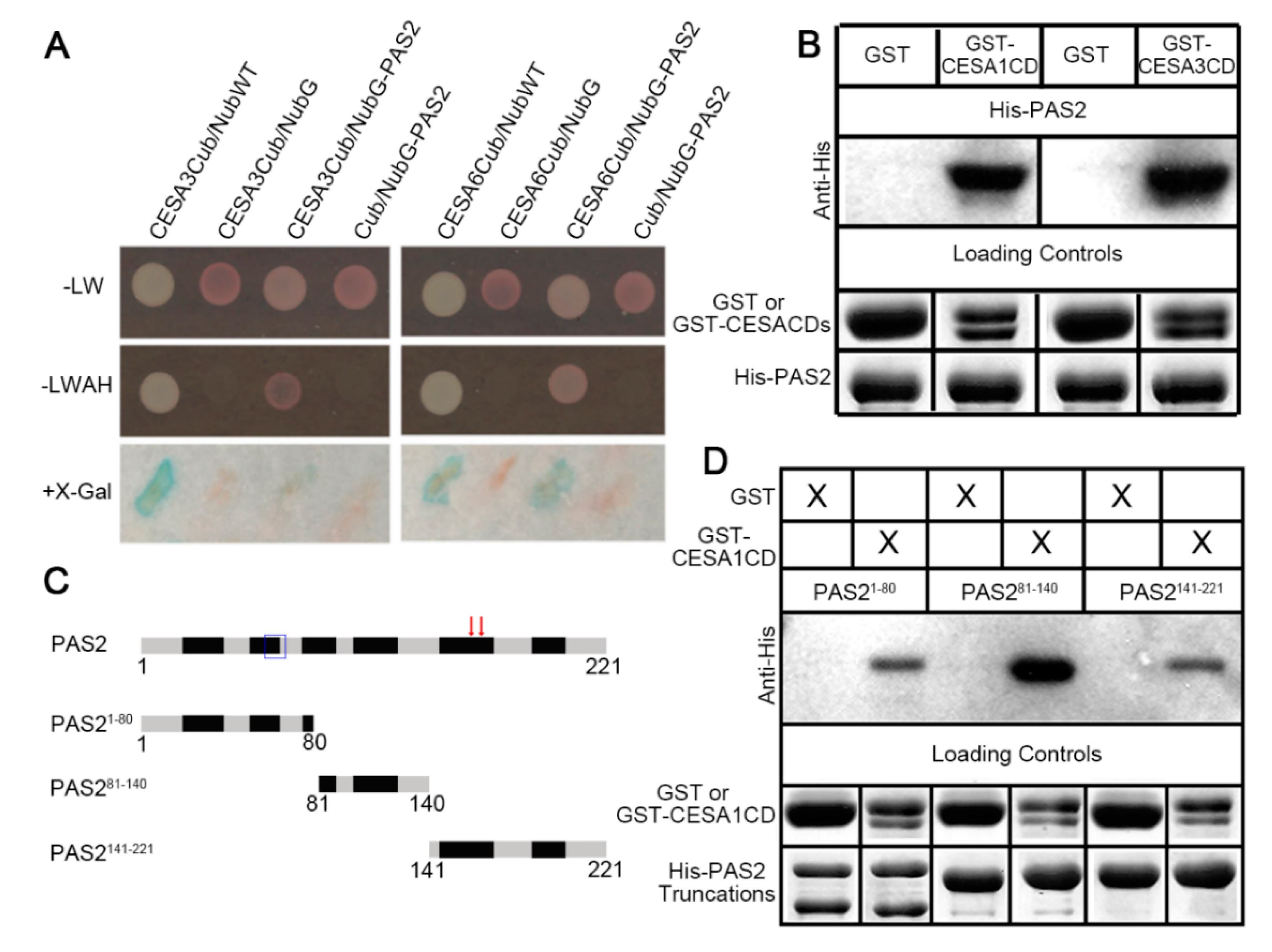
2.1. PAS2 Directly Interacts with CESA Proteins
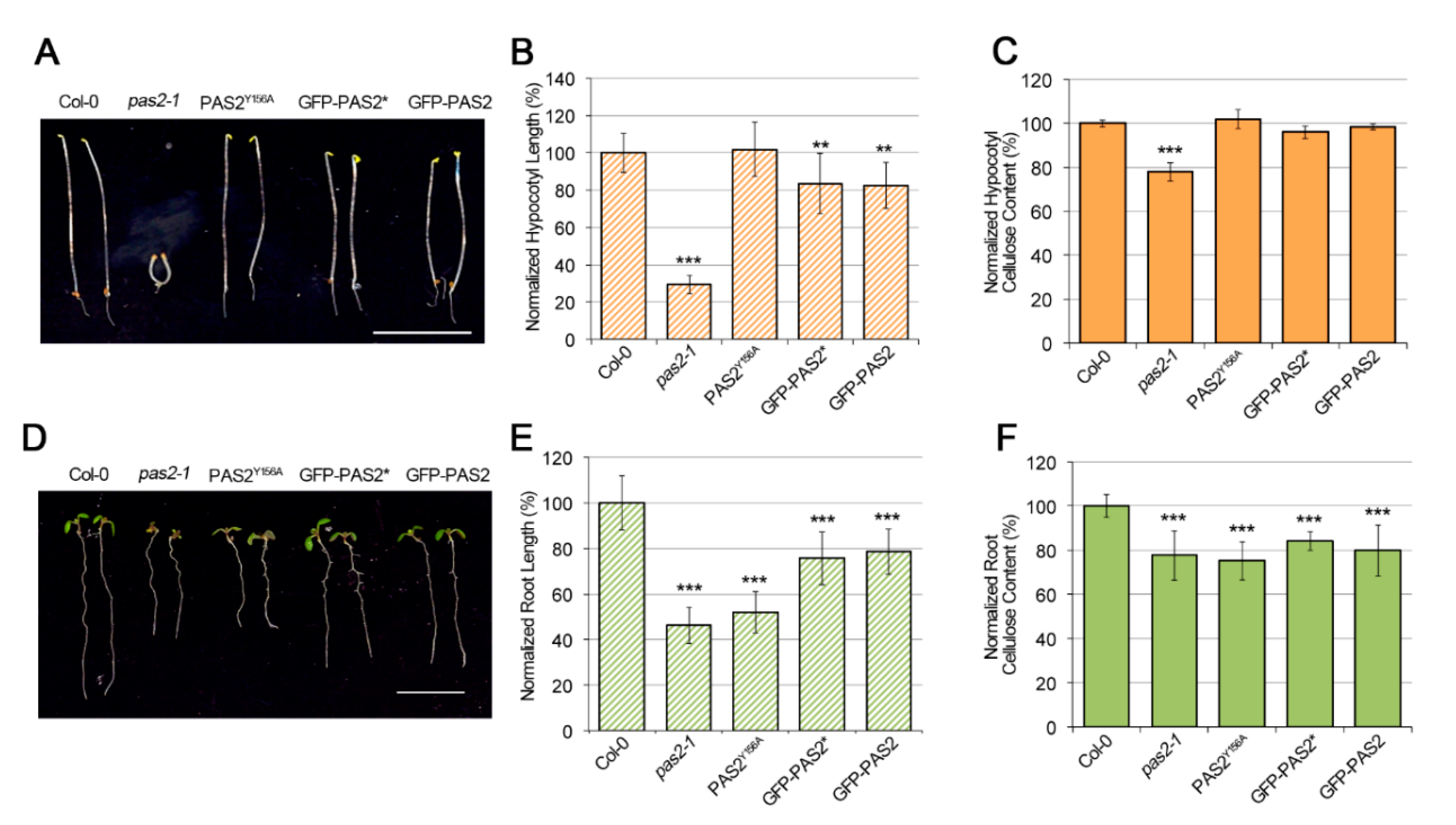
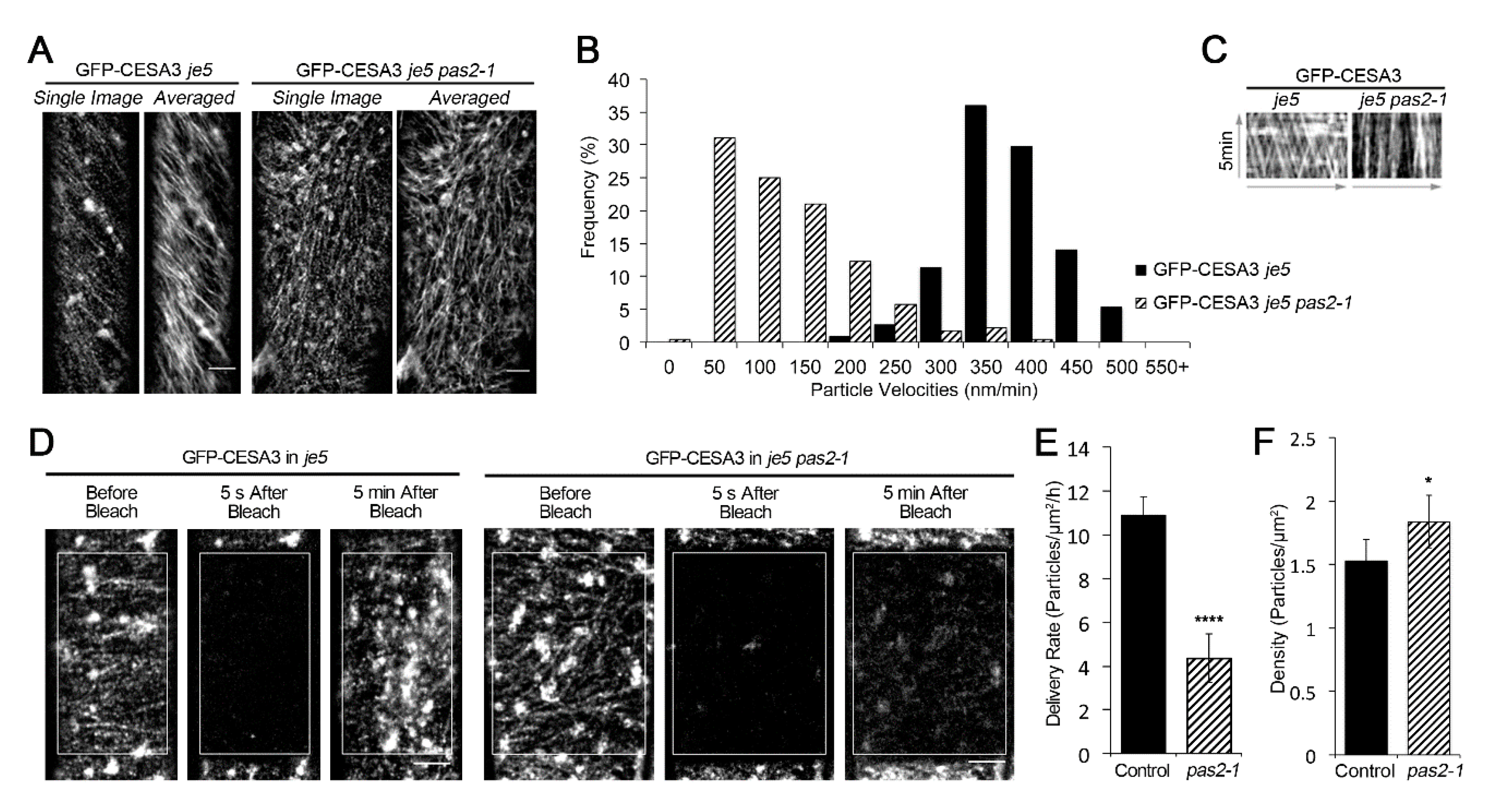
2.2. Mutants of PAS2 Have Reduced Cellulose Contents and Reduced Motility of GFP-CESA3
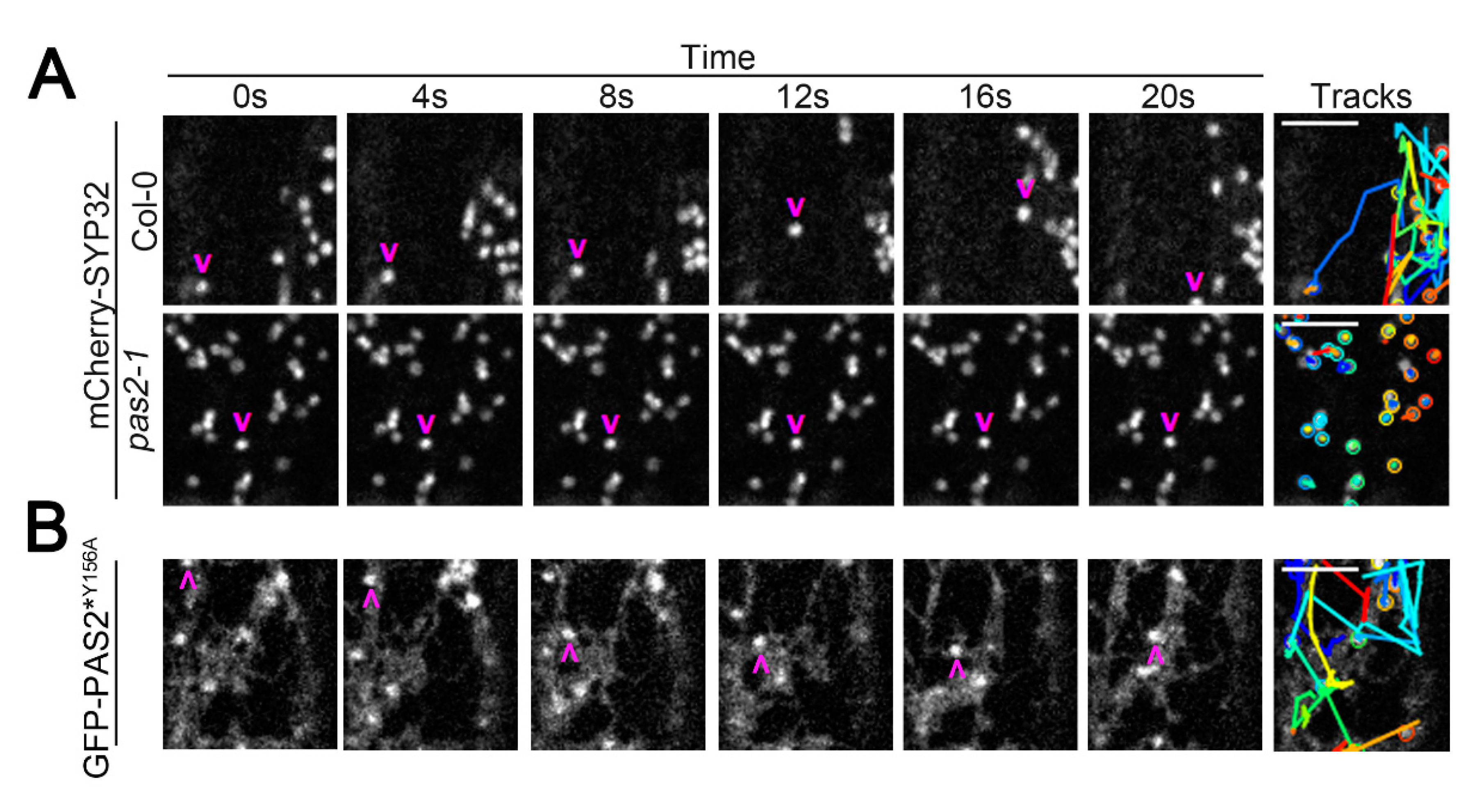
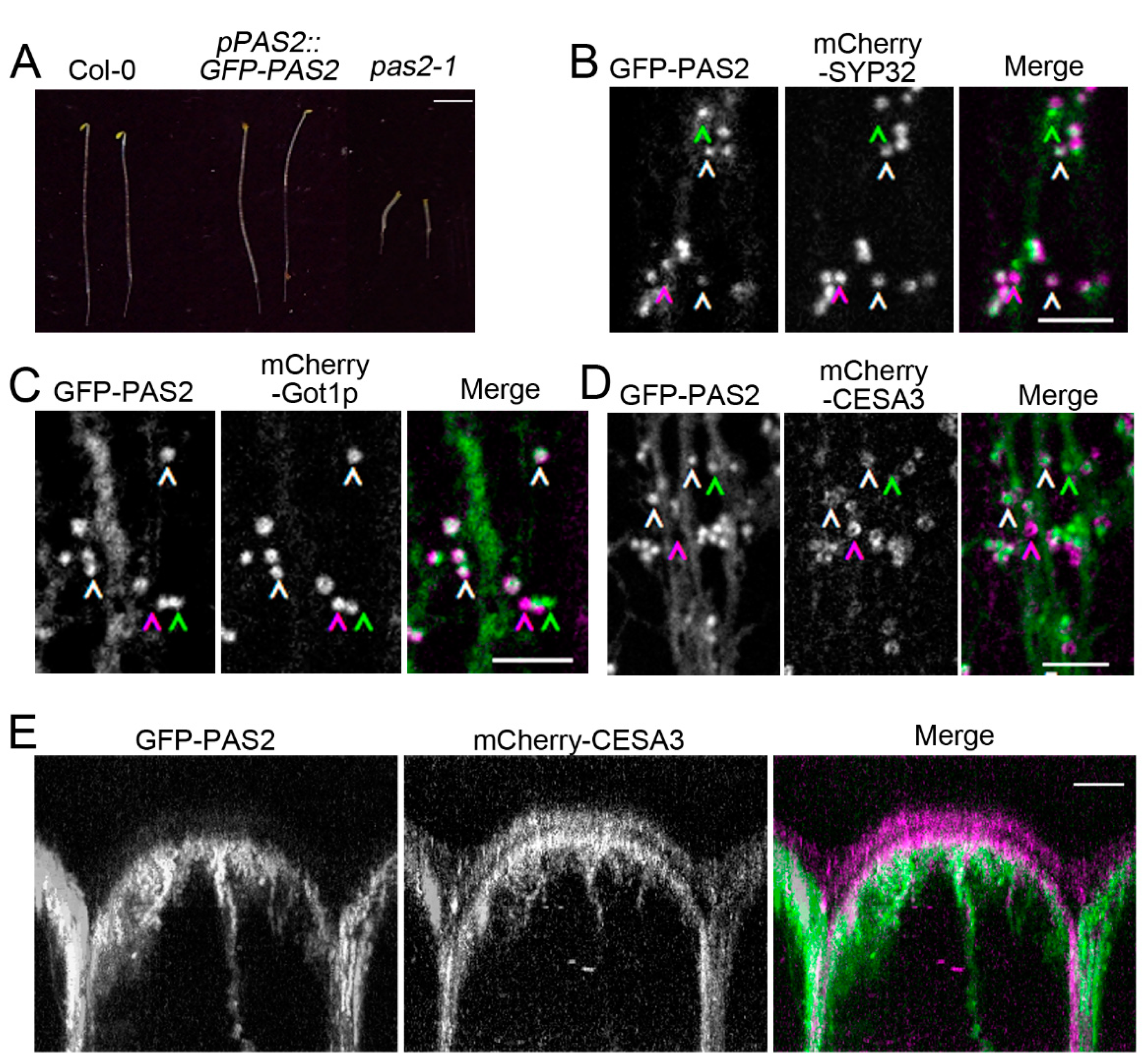
2.3. PAS2 Is Required for Proper Golgi Dynamics
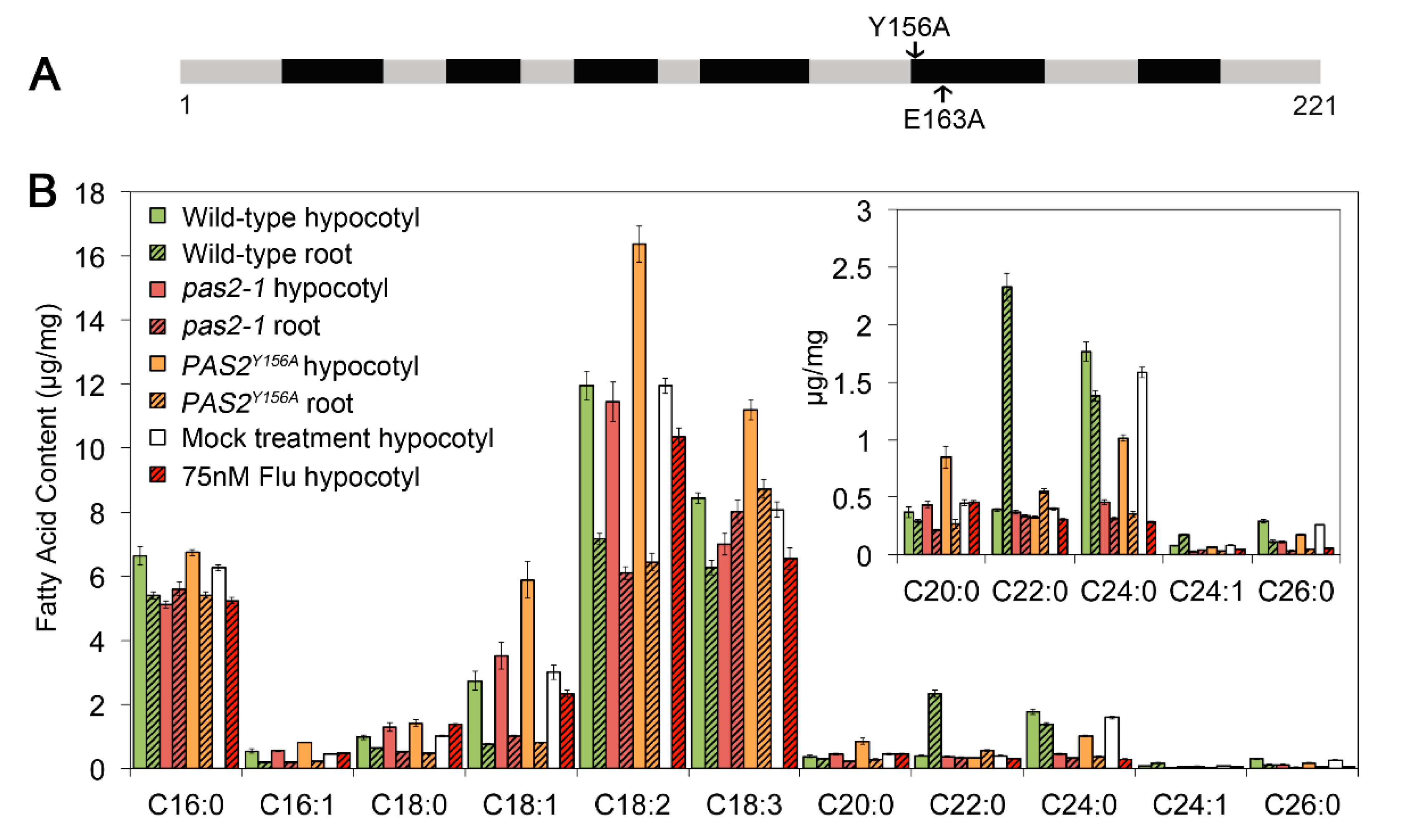
2.4. Disruption of PAS2′s Dehydratase Activity Had Different Effects in Etiolated Hypocotyls and Light-Grown Roots
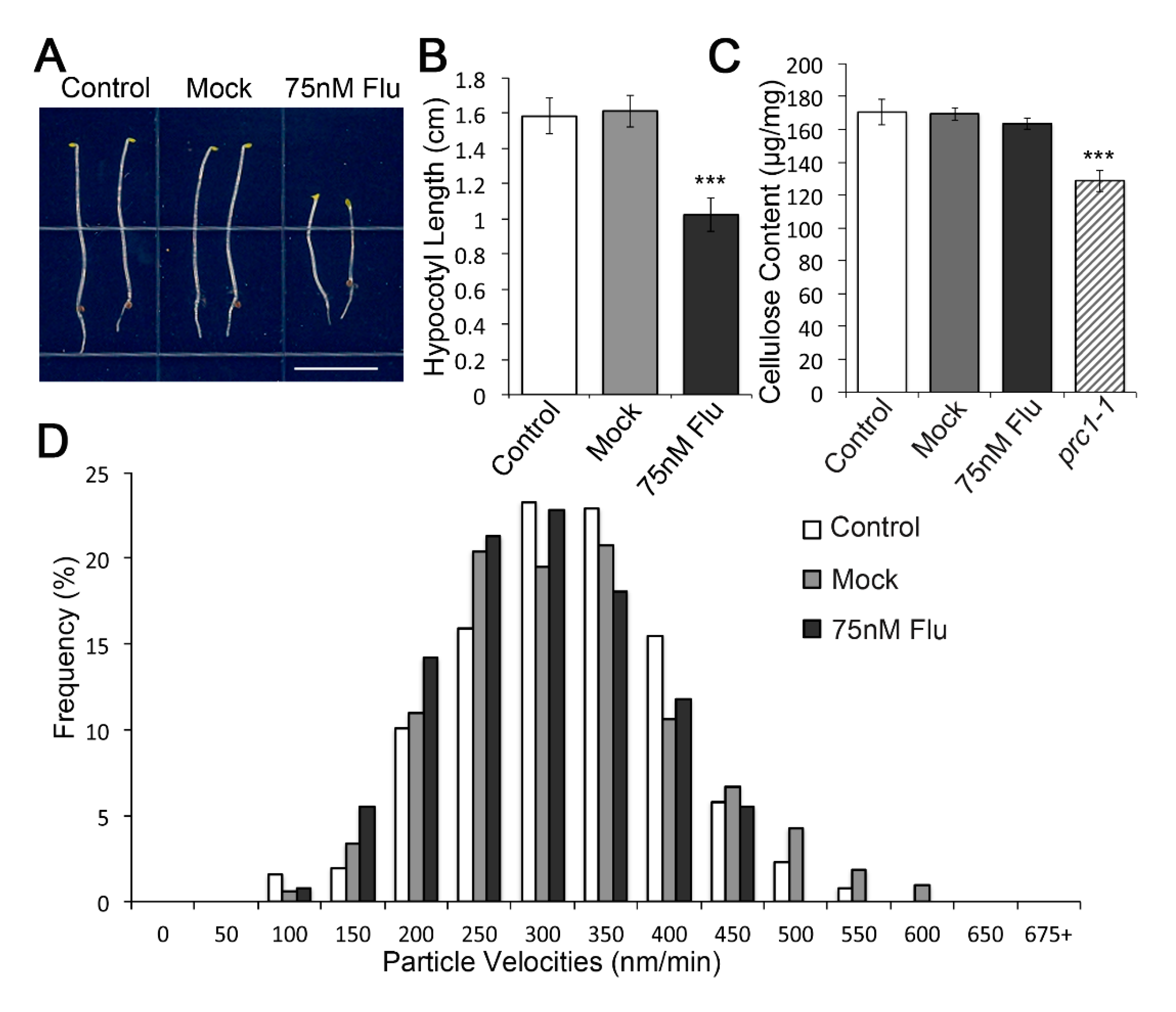
2.5. VLCFA Reduction by Flufenacet in Wild Type Hypocotyls to a Level Comparable to That of pas2-1 Mutants Did Not Cause Cellulose Synthesis Defect
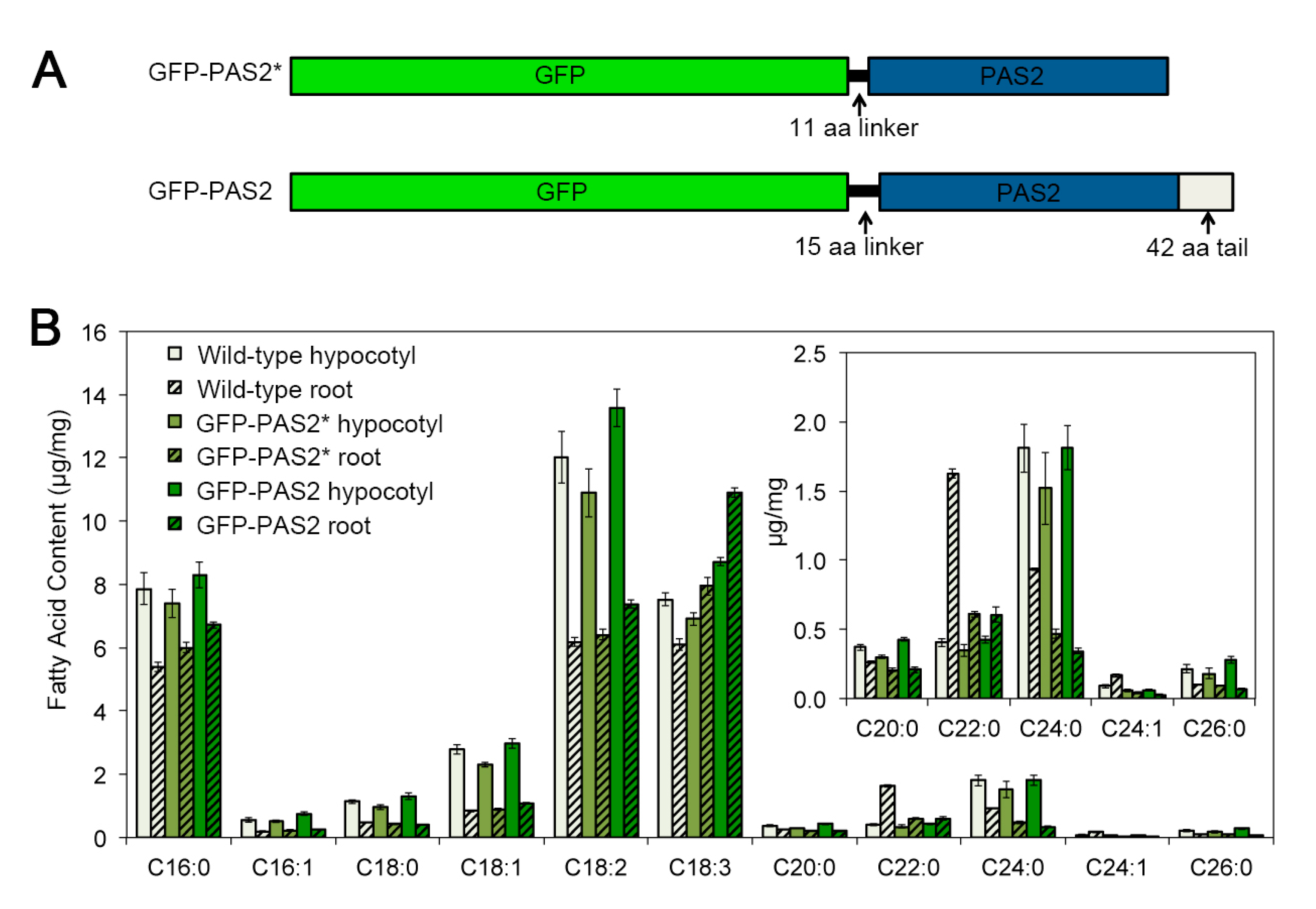
2.6. Investigation of PAS2′s Subcellular Localization in Arabidopsis Hypocotyls
3. Discussion
4. Methods
4.1. Plant Material and Growth Conditions
4.2. Transgenic Lines
4.3. Split-Ubiquitin Yeast Two-Hybrid Assay
4.4. Cellulose Content Measurement
4.5. Live Cell Imaging
4.6. Drug Treatments
4.7. Lipid Analysis
4.8. Protein Purification
4.9. In Vitro Pull-Down Assay
4.10. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kimura, S.; Laosinchai, W.; Itoh, T.; Cui, X.; Linder, C.R.; Brown, R.M. Immunogold Labeling of Rosette Terminal Cellulose-Synthesizing Complexes in the Vascular Plant Vigna angularis. Plant Cell 1999, 11, 2075–2085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brett, C.T. Cellulose microfibrils in plants: Biosynthesis, deposition, and integration into the cell wall. Int. Rev. Cytol. 2000, 199, 161–199. [Google Scholar] [CrossRef] [PubMed]
- Desprez, T.; Juraniec, M.; Crowell, E.F.; Jouy, H.; Pochylova, Z.; Parcy, F.; Hofte, H.; Gonneau, M.; Vernhettes, S. Organization of cellulose synthase complexes involved in primary cell wall synthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 15572–15577. [Google Scholar] [CrossRef] [Green Version]
- Persson, S.; Paredez, A.; Carroll, A.; Palsdottir, H.; Doblin, M.; Poindexter, P.; Khitrov, N.; Auer, M.; Somerville, C.R. Genetic evidence for three unique components in primary cell-wall cellulose synthase complexes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 15566–15571. [Google Scholar] [CrossRef] [Green Version]
- Taylor, N.G.; Laurie, S.; Turner, S.R. Multiple Cellulose Synthase Catalytic Subunits Are Required for Cellulose Synthesis in Arabidopsis. Plant Cell 2000, 12, 2529–2539. [Google Scholar] [CrossRef] [Green Version]
- Taylor, N.G.; Howells, R.M.; Huttly, A.K.; Vickers, K.; Turner, S.R. Interactions among three distinct CesA proteins essential for cellulose synthesis. Proc. Natl. Acad. Sci. USA 2003, 100, 1450–1455. [Google Scholar] [CrossRef] [Green Version]
- Atanassov, I.I.; Pittman, J.K.; Turner, S.R. Elucidating the Mechanisms of Assembly and Subunit Interaction of the Cellulose Synthase Complex ofArabidopsisSecondary Cell Walls. J. Biol. Chem. 2008, 284, 3833–3841. [Google Scholar] [CrossRef] [Green Version]
- Gonneau, M.; Desprez, T.; Guillot, A.; Vernhettes, S.; Höfte, H. Catalytic Subunit Stoichiometry within the Cellulose Synthase Complex. Plant Physiol. 2014, 166, 1709–1712. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.L.; Hammudi, M.B.; Tien, M.; Liu, Y.; You, S.; Taylor-Teeples, M.; Li, W.L.; Schuetz, M.; Brady, S.M.; Douglas, C.J. The Arabidopsis Cellulose Synthase Complex: A Proposed Hexamer of CESA Trimers in an Equimolar Stoichiometry. Plant Cell 2014, 26, 4834–4842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredez, A.R.; Somerville, C.R.; Ehrhardt, D.W. Visualization of Cellulose Synthase Demonstrates Functional Association with Microtubules. Science 2006, 312, 1491–1495. [Google Scholar] [CrossRef] [Green Version]
- Crowell, E.F.; Bischoff, V.; Desprez, T.; Rolland, A.; Stierhof, Y.-D.; Schumacher, K.; Gonneau, M.; Höfte, H.; Vernhettes, S. Pausing of Golgi Bodies on Microtubules Regulates Secretion of Cellulose Synthase Complexes in Arabidopsis. Plant Cell 2009, 21, 1141–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, R.; Lindeboom, J.J.; Paredez, A.R.; Emons, A.M.C.; Ehrhardt, D.W. Arabidopsis cortical microtubules position cellulose synthase delivery to the plasma membrane and interact with cellulose synthase trafficking compartments. Nat. Cell Biol. 2009, 11, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Reiter, W.-D. Biosynthesis and properties of the plant cell wall. Curr. Opin. Plant Biol. 2002, 5, 536–542. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef]
- Baskin, T.I.; Gu, Y. Making parallel lines meet. Cell Adhes. Migr. 2012, 6, 404–408. [Google Scholar] [CrossRef] [Green Version]
- Crowell, E.; Gonneau, M.; Stierhof, Y.-D.; Höfte, H.; Vernhettes, S. Regulated trafficking of cellulose synthases. Curr. Opin. Plant Biol. 2010, 13, 700–705. [Google Scholar] [CrossRef]
- Lei, L.; Zhang, T.; Strasser, R.; Lee, C.M.; Gonneau, M.; Mach, L.; Vernhettes, S.; Kim, S.H.; Cosgrove, D.J.; Li, S.; et al. The jiaoyao1 Mutant Is an Allele of korrigan1 That Abolishes Endoglucanase Activity and Affects the Organization of Both Cellulose Microfibrils and Microtubules in Arabidopsis. Plant Cell 2014, 26, 2601–2616. [Google Scholar] [CrossRef] [Green Version]
- Vain, T.; Crowell, E.F.; Timpano, H.; Biot, E.; Desprez, T.; Mansoori, N.; Trindade, L.M.; Pagant, S.; Robert, S.; Höfte, H.; et al. The Cellulase KORRIGAN Is Part of the Cellulose Synthase Complex. Plant Physiol. 2014, 165, 1521–1532. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Lei, L.; Yingling, Y.G.; Gu, Y. Microtubules and cellulose biosynthesis: The emergence of new players. Curr. Opin. Plant Biol. 2015, 28, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Kaplinsky, N.; Bringmann, M.; Cobb, A.; Carroll, A.; Sampathkumar, A.; Baskin, T.I.; Persson, S.; Somerville, C.R. Identification of a cellulose synthase-associated protein required for cellulose biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 12866–12871. [Google Scholar] [CrossRef] [Green Version]
- Bringmann, M.; Li, E.; Sampathkumar, A.; Kocabek, T.; Hauser, M.-T.; Persson, S. POM-POM2/CELLULOSE SYNTHASE INTERACTING1 Is Essential for the Functional Association of Cellulose Synthase and Microtubules in Arabidopsis. Plant Cell 2012, 24, 163–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Lei, L.; Somerville, C.R.; Gu, Y. Cellulose synthase interactive protein 1 (CSI1) links microtubules and cellulose synthase complexes. Proc. Natl. Acad. Sci. USA 2012, 109, 185–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endler, A.; Kesten, C.; Schneider, R.; Zhang, Y.; Ivakov, A.; Froehlich, A.; Funke, N.; Persson, S. A Mechanism for Sustained Cellulose Synthesis during Salt Stress. Cell 2015, 162, 1353–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, H.; Wei, D.; Gu, Y.; Li, S. A historical perspective on the regulation of cellulose biosynthesis. Carbohydr. Polym. 2021, 252, 117022. [Google Scholar] [CrossRef]
- Haigler, C.H.; Brown, R.M. Transport of rosettes from the golgi apparatus to the plasma membrane in isolated mesophyll cells ofZinnia elegans during differentiation to tracheary elements in suspension culture. Protoplasma 1986, 134, 111–120. [Google Scholar] [CrossRef]
- Miart, F.; Desprez, T.; Biot, E.; Morin, H.; Belcram, K.; Höfte, H.; Gonneau, M.; Vernhettes, S. Spatio-temporal analysis of cellulose synthesis during cell plate formation in Arabidopsis. Plant J. 2013, 77, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Bashline, L.; Li, S.; Anderson, C.T.; Lei, L.; Gu, Y. The Endocytosis of Cellulose Synthase in Arabidopsis Is Dependent on μ2, a Clathrin-Mediated Endocytosis Adaptin. Plant Physiol. 2013, 163, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Bashline, L.; Li, S.; Zhu, X.; Gu, Y. The TWD40-2 protein and the AP2 complex cooperate in the clathrin-mediated endocytosis of cellulose synthase to regulate cellulose biosynthesis. Proc. Natl. Acad. Sci. USA 2015, 112, 12870–12875. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Singh, A.; Bashline, L.; Li, S.; Yingling, Y.G.; Gu, Y. CELLULOSE SYNTHASE INTERACTIVE1 Is Required for Fast Recycling of Cellulose Synthase Complexes to the Plasma Membrane in Arabidopsis. Plant Cell 2015, 27, 2926–2940. [Google Scholar] [CrossRef] [Green Version]
- Faure, J.D.; Vittorioso, P.; Santoni, V.; Fraisier, V.; Prinsen, E.; Barlier, I.; Van Onckelen, H.; Caboche, M.; Bellini, C. The PASTICCINO genes of Arabidopsis thaliana are involved in the control of cell division and differentiation. Development 1998, 125, 909–918. [Google Scholar]
- Wishart, M.J.; Denu, J.M.; Williams, J.A.; Dixon, J.E. A Single Mutation Converts a Novel Phosphotyrosine Binding Domain into a Dual-specificity Phosphatase. J. Biol. Chem. 1995, 270, 26782–26785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vivo, I.; Cui, X.; Domen, J.; Cleary, M.L. Growth stimulation of primary B cell precursors by the anti-phosphatase Sbf1. Proc. Natl. Acad. Sci. USA 1998, 95, 9471–9476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellec, Y.; Harrar, Y.; Butaeye, C.; Darnet, S.; Bellini, C.; Faure, J.-D. Pasticcino2 is a protein tyrosine phosphatase-like involved in cell proliferation and differentiation in Arabidopsis. Plant J. 2002, 32, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Haberer, G.; Erschadi, S.; Ruiz, R.A.T. The Arabidopsis gene PEPINO/PASTICCINO2 is required for proliferation control of meristematic and non-meristematic cells and encodes a putative anti-phosphatase. Dev. Genes Evol. 2002, 212, 542–550. [Google Scholar] [CrossRef]
- Da Costa, M.; Bach, L.; Landrieu, I.; Bellec, Y.; Catrice, O.; Brown, S.; De Veylder, L.; Lippens, G.; Inzé, D.; Faure, J.-D. Arabidopsis PASTICCINO2 Is an Antiphosphatase Involved in Regulation of Cyclin-Dependent Kinase A. Plant Cell 2006, 18, 1426–1437. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Castillo, L.P.; Mnaimneh, S.; Hughes, T.R.; Brown, G.W. A Survey of Essential Gene Function in the Yeast Cell Division Cycle. Mol. Biol. Cell 2006, 17, 4736–4747. [Google Scholar] [CrossRef]
- Kihara, A.; Sakuraba, H.; Ikeda, M.; Denpoh, A.; Igarashi, Y. Membrane Topology and Essential Amino Acid Residues of Phs1, a 3-Hydroxyacyl-CoA Dehydratase Involved in Very Long-chain Fatty Acid Elongation. J. Biol. Chem. 2008, 283, 11199–11209. [Google Scholar] [CrossRef] [Green Version]
- Bach, L.; Michaelson, L.V.; Haslam, R.; Bellec, Y.; Gissot, L.; Marion, J.; Da Costa, M.; Boutin, J.P.; Miquel, M.; Tellier, F.; et al. The very-long-chain hydroxy fatty acyl-CoA dehydratase PASTICCINO2 is essential and limiting for plant development. Proc. Natl. Acad. Sci. USA 2008, 105, 14727–14731. [Google Scholar] [CrossRef] [Green Version]
- Harrar, Y.; Bellec, Y.; Bellini, C.; Faure, J.-D. Hormonal Control of Cell Proliferation Requires PASTICCINO Genes. Plant Physiol. 2003, 132, 1217–1227. [Google Scholar] [CrossRef] [Green Version]
- Fagard, M.; Desnos, T.; Desprez, T.; Goubet, F.; Refregier, G.; Mouille, G.; McCann, M.; Rayon, C.; Vernhettes, S.; Höfte, H. PROCUSTE1 Encodes a Cellulose Synthase Required for Normal Cell Elongation Specifically in Roots and Dark-Grown Hypocotyls of Arabidopsis. Plant Cell 2000, 12, 2409–2423. [Google Scholar] [CrossRef] [Green Version]
- Updegraff, D.M. Semimicro determination of cellulose inbiological materials. Anal. Biochem. 1969, 32, 420–424. [Google Scholar] [CrossRef]
- Denic, V.; Weissman, J.S. A Molecular Caliper Mechanism for Determining Very Long-Chain Fatty Acid Length. Cell 2007, 130, 663–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bach, L.; Faure, J.-D. Role of very-long-chain fatty acids in plant development, when chain length does matter. Comptes Rendus Biol. 2010, 333, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Trenkamp, S.; Martin, W.; Tietjen, K. Specific and differential inhibition of very-long-chain fatty acid elongases from Arabidopsis thaliana by different herbicides. Proc. Natl. Acad. Sci. USA 2004, 101, 11903–11908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bach, L.; Gissot, L.; Marion, J.; Tellier, F.; Moreau, P.; Satiat-Jeunemaitre, B.; Palauqui, J.-C.; Napier, J.A.; Faure, J.-D.; Hua, F.; et al. Very-long-chain fatty acids are required for cell plate formation during cytokinesis in Arabidopsis thaliana. J. Cell Sci. 2011, 124, 3223–3234. [Google Scholar] [CrossRef] [Green Version]
- Geldner, N.; Dénervaud-Tendon, V.; Hyman, D.L.; Mayer, U.; Stierhof, Y.-D.; Chory, J. Rapid, combinatorial analysis of membrane compartments in intact plants with a multicolor marker set. Plant J. 2009, 59, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Bashline, L.; Li, S.; Gu, Y. The trafficking of the cellulose synthase complex in higher plants. Ann. Bot. 2014, 114, 1059–1067. [Google Scholar] [CrossRef]
- Morineau, C.; Gissot, L.; Bellec, Y.; Hematy, K.; Tellier, F.; Renne, C.; Haslam, R.P.; Beaudoin, F.; Napier, J.A.; Faure, J.-D. Dual Fatty Acid Elongase Complex Interactions in Arabidopsis. PLoS ONE 2016, 11, e0160631. [Google Scholar] [CrossRef] [Green Version]
- Nobusawa, T.; Okushima, Y.; Nagata, N.; Kojima, M.; Sakakibara, H.; Umeda, M. Synthesis of Very-Long-Chain Fatty Acids in the Epidermis Controls Plant Organ Growth by Restricting Cell Proliferation. PLoS Biol. 2013, 11, e1001531. [Google Scholar] [CrossRef] [Green Version]
- Markham, J.E.; Molino, D.; Gissot, L.; Bellec, Y.; Hématy, K.; Marion, J.; Belcram, K.; Palauqui, J.-C.; Satiat-Jeunemaître, B.; Faure, J.-D. Sphingolipids Containing Very-Long-Chain Fatty Acids Define a Secretory Pathway for Specific Polar Plasma Membrane Protein Targeting in Arabidopsis. Plant Cell 2011, 23, 2362–2378. [Google Scholar] [CrossRef] [Green Version]
- Molino, D.; Van Der Giessen, E.; Gissot, L.; Hematy, K.; Marion, J.; Barthélémy, J.; Bellec, Y.; Vernhettes, S.; Satiat-Jeunemaitre, B.; Galli, T.; et al. Inhibition of very long acyl chain sphingolipid synthesis modifies membrane dynamics during plant cytokinesis. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2014, 1841, 1422–1430. [Google Scholar] [CrossRef] [PubMed]
- Granrut, A.D.B.D.; Cacas, J.-L. How Very-Long-Chain Fatty Acids Could Signal Stressful Conditions in Plants? Front. Plant Sci. 2016, 7, 1490. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Ishikawa, T.; Rennie, E.A.; Murawska, G.M.; Lao, J.; Yan, J.; Tsai, A.Y.-L.; Baidoo, E.E.; Xu, J.; Keasling, J.D.; et al. Loss of Inositol Phosphorylceramide Sphingolipid Mannosylation Induces Plant Immune Responses and Reduces Cellulose Content in Arabidopsis. Plant Cell 2016, 28, 2991–3004. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Song, B.; Shen, W.; Ding, S.-Y. A mutation in the catalytic domain of cellulose synthase 6 halts its transport to the Golgi apparatus. J. Exp. Bot. 2019, 70, 6071–6083. [Google Scholar] [CrossRef]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY™ vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method forAgrobacterium-mediated transformation ofArabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Li, S.; Du, J.; Bashline, L.; Gu, Y. CELLULOSE SYNTHASE INTERACTIVE3 Regulates Cellulose Biosynthesis in Both a Microtubule-Dependent and Microtubule-Independent Manner in Arabidopsis. Plant Cell 2013, 25, 4912–4923. [Google Scholar] [CrossRef] [Green Version]
- Bashline, L.; Gu, Y. Using the Split-Ubiquitin Yeast Two-Hybrid System to Test Protein–Protein Interactions of Transmembrane Proteins. Adv. Struct. Saf. Stud. 2014, 1242, 143–158. [Google Scholar] [CrossRef]
- Li, Y.; Beisson, F.; Pollard, M.; Ohlrogge, J.B.; Li-Beisson, Y.; Beisson, F. Oil content of Arabidopsis seeds: The influence of seed anatomy, light and plant-to-plant variation. Phytochemistry 2006, 67, 904–915. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Tellier, F.; Gu, Y.; Li, S. Disruption of Very-Long-Chain-Fatty Acid Synthesis Has an Impact on the Dynamics of Cellulose Synthase in Arabidopsis thaliana. Plants 2020, 9, 1599. https://doi.org/10.3390/plants9111599
Zhu X, Tellier F, Gu Y, Li S. Disruption of Very-Long-Chain-Fatty Acid Synthesis Has an Impact on the Dynamics of Cellulose Synthase in Arabidopsis thaliana. Plants. 2020; 9(11):1599. https://doi.org/10.3390/plants9111599
Chicago/Turabian StyleZhu, Xiaoyu, Frédérique Tellier, Ying Gu, and Shundai Li. 2020. "Disruption of Very-Long-Chain-Fatty Acid Synthesis Has an Impact on the Dynamics of Cellulose Synthase in Arabidopsis thaliana" Plants 9, no. 11: 1599. https://doi.org/10.3390/plants9111599