2.1. Production of Interspecific Hybrids
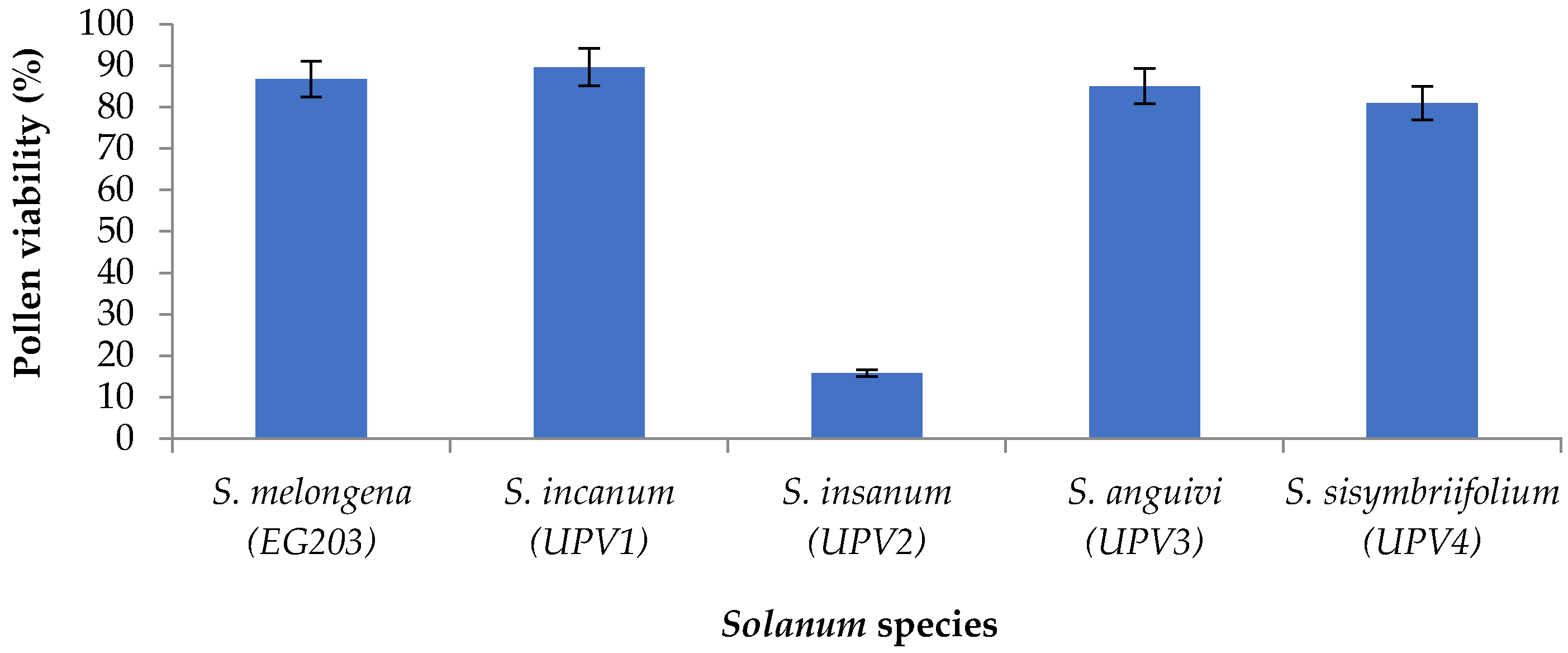
The results of pollen viability in cultivated eggplant and wild accession are presented in
Figure 1. Cultivated eggplant EG203 had high pollen viability (86.76%). Pollen viability in wild accessions was variable depending on the species. The highest pollen fertility was observed in
S. incanum UPV1 (89.65%), followed by
S. anguivi UPV3 (85.03%) and
S. sisymbriifolium UPV4 (80.94%). On the contrary,
S. insanum UPV2 had very low pollen fertility (15.78%).
The results of the interspecific hybridization between the cultivated eggplant EG203 and wild species with respect to mean fruit set (%), number of seeds per fruit, and seed germination (%) are presented in
Table 1. Cultivated eggplant EG203 was successfully crossed with wild accessions when used as the male parent, except in crosses with
S. sisymbriifolium UPV4. However, when
S. melongena EG203 was used as the female parent, crosses were successful only with
S. incanum UPV1. The percentage of fruit set of interspecific hybridizations was variable depending on the direction of the hybridization and the wild species involved. The maximum fruit set (68.8%) was recorded in the cross
S. incanum UPV1 ×
S. melongena EG203 followed by
S. melongena EG203 ×
S. incanum UPV1 (65.2%),
S. anguivi UPV3 ×
S. melongena EG203 (46.2%) and
S. insanum UPV2 ×
S. melongena EG203 (40.0%). When using EG203 as a male parent, the largest amount of seeds per fruit was obtained in hybridizations with
S. insanum UPV2, with an average value of 0.85 g/fruit, which is equivalent to more than 250 seeds/fruit, followed by hybridizations with
S. anguivi UPV3 (0.30 g/fruit), which is equivalent to around 70 seeds/fruit. Seed quantity per fruit in the hybridization with
S. incanum UPV1 was 0.03 g/fruit, which is equivalent to 17 seeds/fruit. When using
S. melongena EG203 as a female parent, a large number of seeds were obtained with the only wild accession (
S. incanum UPV1) in which the crosses were successful.
Considering the three wild species (
S. incanum,
S. insanum, and
S. anguivi) for which we were able to obtain interspecific hybrids with
S. melongena, in general, our results are in agreement with previous studies [
25]. In this respect, Devi et al. [
26], found that
S. incanum was highly crossable with the cultivated eggplant genotypes. In another study, Plazas et al. [
27] also reported that crosses between
S. melongena and
S. incanum, as well as with
S. insanum and
S. anguivi, were successful in both directions, and the highest rate of fruit set was obtained when using
S. melongena as the female parent with
S. insanum, which presented a fruit set above 15%. When using
S. melongena as the male parent, the highest fruit set was obtained with
S. anguivi with values above 25%. In another study, reciprocal crosses were also successful between
S. melongena and
S. incanum [
28]. However, in contrast to our results and those of Plazas et al. [
27], Afful et al. [
29] reported that no fruit set was observed when several
S. melongena cultivars were crossed as the female parent with
S. anguivi. The variation between our results and those of previous studies may be related to the different genetic background of the parental lines [
30], to irregular chromosome associations during the formation of gametes [
31,
32], or to different environmental conditions.
The crosses we performed between
S. melongena and
S. sisymbriifolium were not successful in any of the directions (
Table 1). In another work, Plazas et al. [
27] found that a low fruit set was obtained after crossing
S. melongena and
S. sisymbriifolium; however, these fruits were parthenocarpic. This result confirms that
S. sisymbriifolium is very distant from eggplant [
33], which is considered a tertiary gene pool species. However, hybrids and backcrosses of
S. melongena with another tertiary gene pool species (
S. elaeagnifolium Cav.) have been obtained by García-Fortea et al. [
34] through embryo rescue.
We found that seed germination percentage among the interspecific hybrids was very high for those involving
S. melongena as the male parent, and
S. insanum UPV2 (90%), as well as
S. anguivi UPV3 (88%) as females. However, no germination was observed in the cross between
S. melongena EG2013 with
S. incanum UPV1 in both directions (
Table 1). Furthermore, most of the seeds in these two interspecific hybrids looked as if they were incompletely developed or immature. In a study in which crosses were made between
S. incanum and
S. melongena, Anis et al. [
35], found that unilateral hybrids could be obtained when
S. incanum was used as the female parent. In contrast, Plazas et al. [
27] found that a high seed germination was obtained for the hybrids between
S. melongena and
S. incanum. In addition, Devi et al. [
26] reported that interspecific hybrids of
S. melongena with
S. incanum produced viable and highly vigorous plants with earliness and higher yield. These differences in germination among different studies could be explained by the different genetic background of parental lines despite the same species combination. Moreover, in our case, the abnormal seeds (empty seeds, seeds with a different appearance, low weight) obtained from the interspecific crosses might also have contributed to the lack of germination observed. In this way, in other species, seeds have been reported to contain an under-developed embryo and/or endosperm, which prevent germination [
36].
Given that no viable seeds were obtained in our hybridizations between
S. melongena EG203 and
S. incanum UPV1 in both directions, embryo rescue was employed to obtain hybrids from the crosses between cultivated eggplant EG203 and UPV1 (
Figure 2). In this respect, embryo rescue has been widely used for producing plants from hybridizations in which failure of the endosperm to develop properly causes embryo abortion [
37]. The responses obtained by us of three different tissue culture media (M1–M3) for germinating embryos from the crosses between
S. melongena EG203 and
S. incanum UPV1 at three different stages (globular, heart, and torpedo stage) are presented in
Table 2. The results showed that the torpedo stage was the best embryo stage to germinate interspecific hybrid plantlets, with regeneration efficiency from 78.9% to 88.5% when
S. melongena EG203 is used as the female parent and 20.5% to 32.6% when
S. incanum UPV1 is used as the female parent. In contrast, the globular stage in both directions of interspecific hybridizations did not produce any plantlets under any of the three media used in this study. Regarding culture media, the MS medium supplemented with 0.01 IAA and 0.1 Kin (MS2) was the best medium to develop plantlets from embryos, with regeneration efficiency reaching up to 88.5% when EG203 was used as the female parent and 32.6% when UPV1 was used as the female parent. However, several abnormal plantlets were observed in this same MS2 medium. The MS medium without growth regulators (MS1) produced the lowest number of normal plantlets. Other works have evaluated the use of embryo rescue and different media for obtaining interspecific hybrids of eggplant, obtaining a diversity of results. In this respect, Verba et al. [
38] found that embryos of the combination
S. melongena × S. integrifolium developed into normal plantlets when torpedo embryos were cultured on MS medium with TDZ (0.1 mg/L). In another work, Sharma et al. [
39] achieved partial success in obtaining hybrid plants between
S. melongena ×
S. sisymbriifolium when torpedo embryos were cultured on Nitsch and Nitsch [
40] medium; however, the plants could not survive for a very long time and died. Conversely, in the interspecific crosses between
S. melongena and
S. khasianum Clark [
41], fertile hybrids were obtained when embryos 25 days old were cultured on Nitsch and Nitsch medium. Embryo rescue by culturing immature ovules 15 to 27 days old after pollination on MS medium without growth regulators was also used by Bletsos et al. [
42] to obtain hybrids between
S. torvum Sw. and
S. melongena. These results indicate that the success and efficiency of embryo rescue in interspecific hybrids of eggplant largely depend on the specific hybrid combination and media used.
2.3. Assessment of Disease Resistance
The resistance reaction and category of the susceptible control, four parents, and interspecific hybrids to
R. solanacearum strains Pss97 and Pss2016 at four weeks after inoculation is presented in
Table 4. The susceptible check (EG048) displayed the expected reactions to strains Pss97 and Pss2016, and all EG048 plants wilted and died rapidly at two and three weeks after inoculation by Pss97 and Pss2016, respectively. EG203 parent was resistant to both strains with 11.1% of wilting percentage (W%) and 9.9% of disease index (DI%) for the Pss97 strain, and 33.3% W%, and 20% of DI% for the Pss2016 strain.
Solanum incanum UPV1 displayed a resistant reaction against Pss2016, while
S. anguivi UPV3 was moderately resistant against this same strain. However, both of them were susceptible against strain Pss97.
Solanum insanum UPV2 was susceptible against both strains. However, all interspecific hybrids were susceptible to both strains, with W% and DI%, ranging between 56.4% and 100%. The symptoms appeared one week after inoculation, and most of the plants were completely wilting after three weeks. These results indicate that the resistances in
S. melongena EG203 and
S. incanum UPV1 and the moderate resistance of
S. anguivi UPV3 against strain Pss2016 must be controlled by a recessive gene(s). However, Salgon et al. [
22] found that in an intraspecific cross, the resistance of EG203 was dominant, although strains of
R. solanacearum different to ours were used. This suggests that the mode of inheritance of the resistance of EG203 to bacterial wilt depends, either on the strain, the genetic background, or both. Given our results, we suggest the evaluation of further germplasm accessions in
S. incanum and
S. anguivi to detect new sources of dominant resistance to bacterial wilt. In this respect, Ano et al. [
44] demonstrated that high levels of resistance can be introgressed from one source of resistance of one related species
S. aethiopicum L., whose wild ancestor is
S. anguivi, into the genetic background of
S. melongena. In any case, despite the susceptibility to bacterial wilt of the four interspecific hybrids obtained, given the vigor of interspecific hybrids of
S. melongena with
S. incanum,
S. insanum, and
S. anguivi [
45], they might be of interest to be used as rootstocks in areas free from bacterial wilt.

 ,
, 


{kind=link}
{kind=link}