Ailanthus altissima Forests Determine a Shift in Herbaceous Layer Richness: A Paired Comparison with Hardwood Native Forests in Sub-Mediterranean Europe


Abstract
:1. Introduction
2. Results
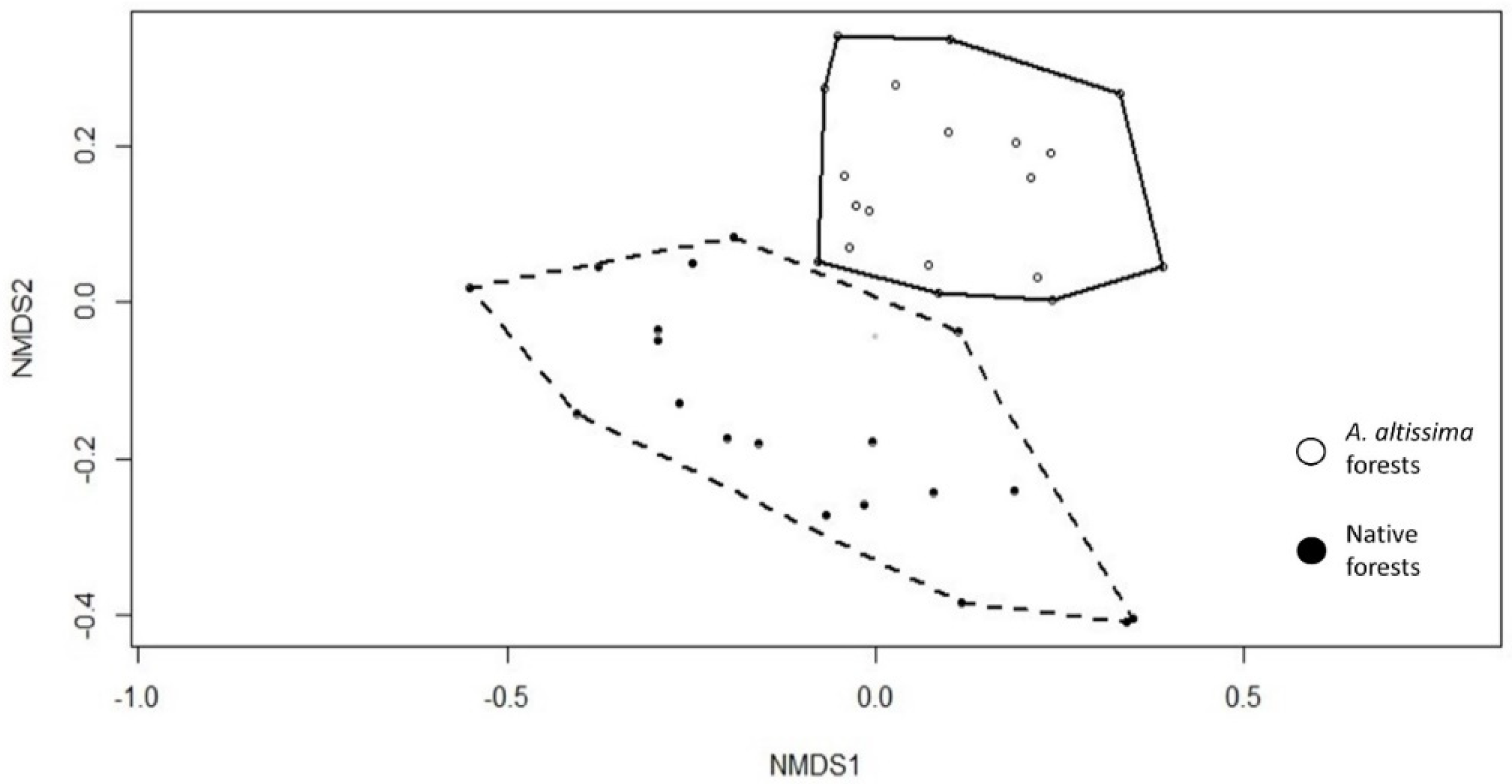
2.1. Community Floristic Diversity
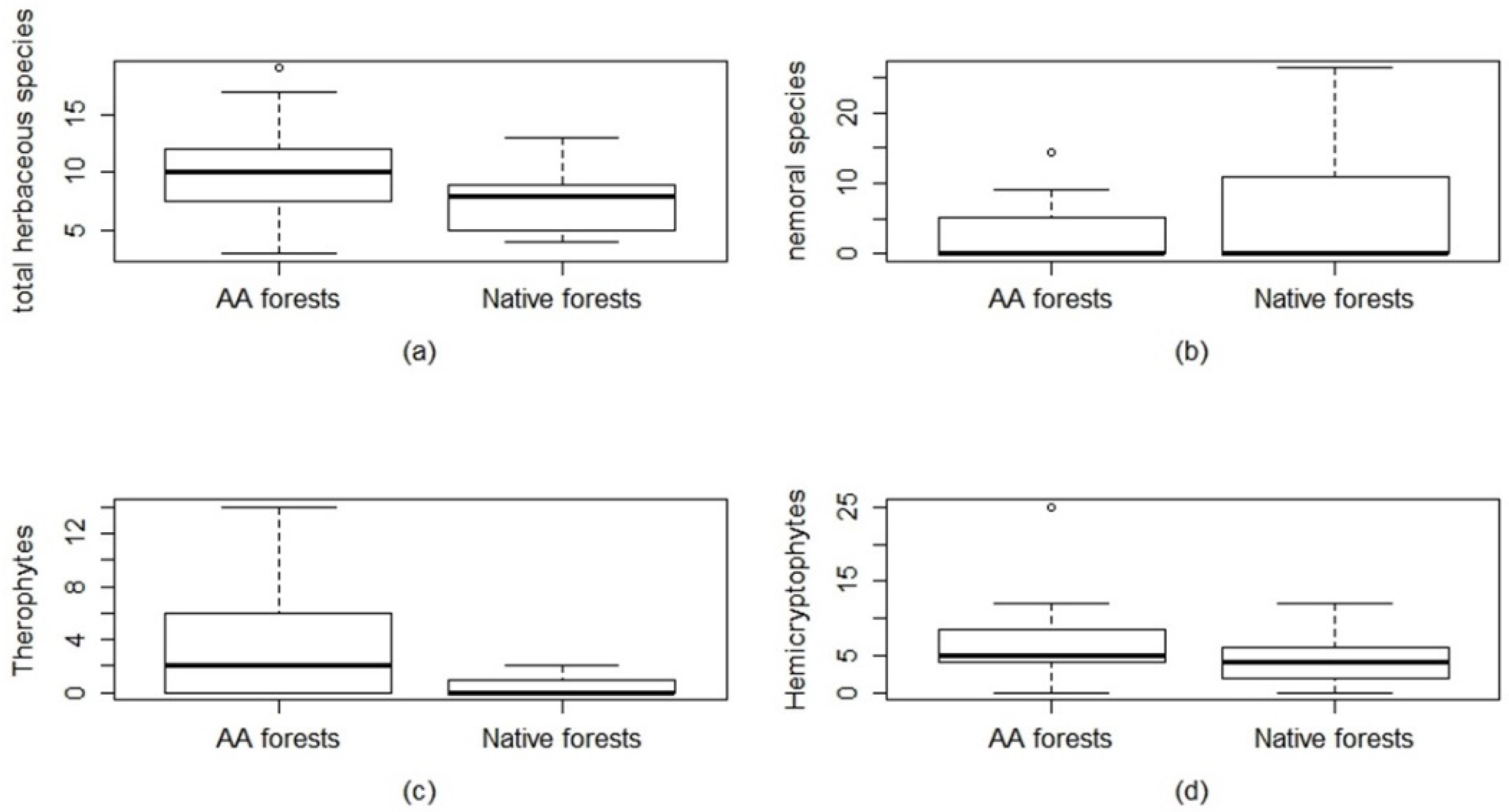
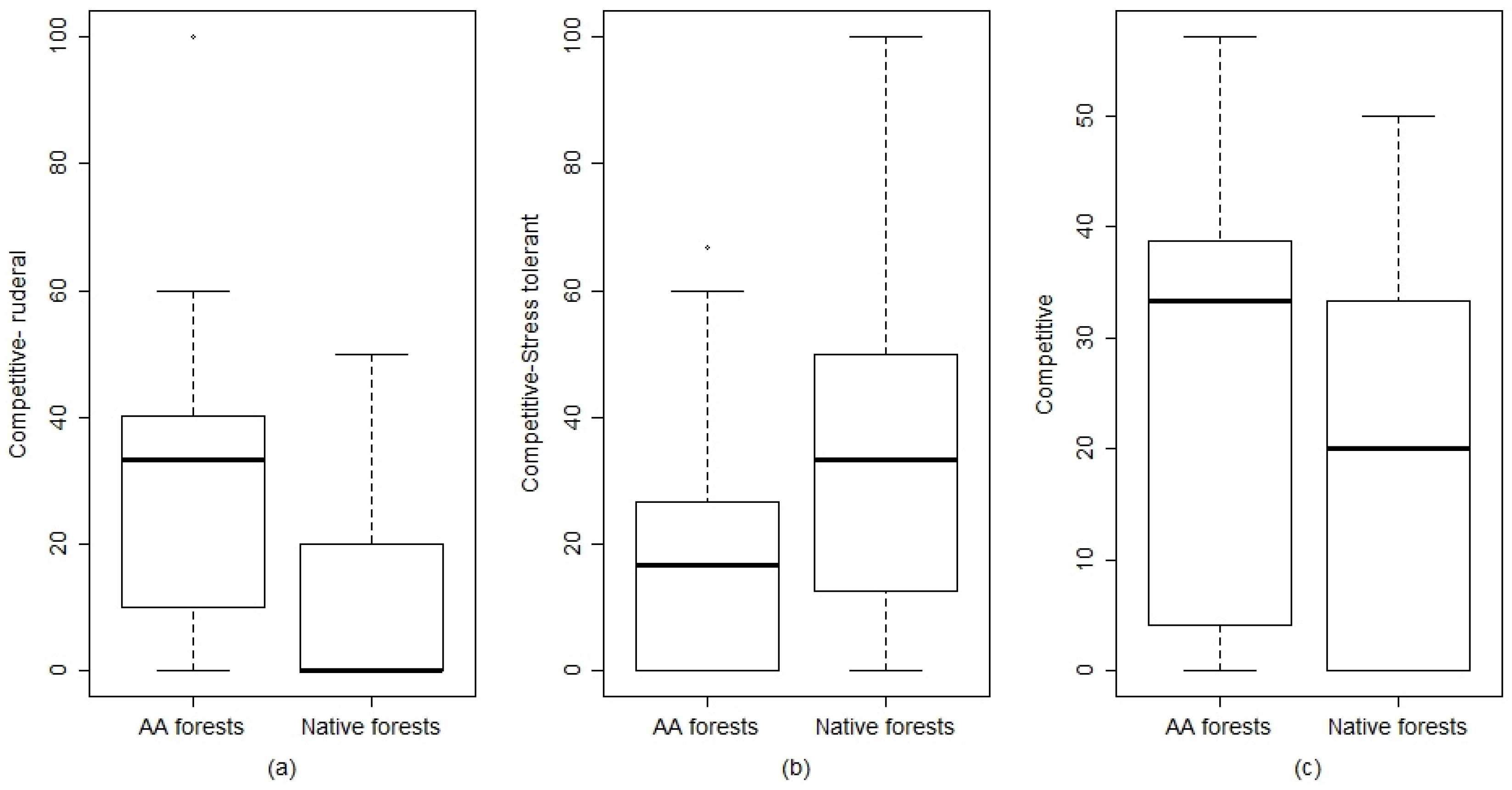
2.2. Herbaceous Layer: Species Diversity, Life Forms, and Grime Strategies
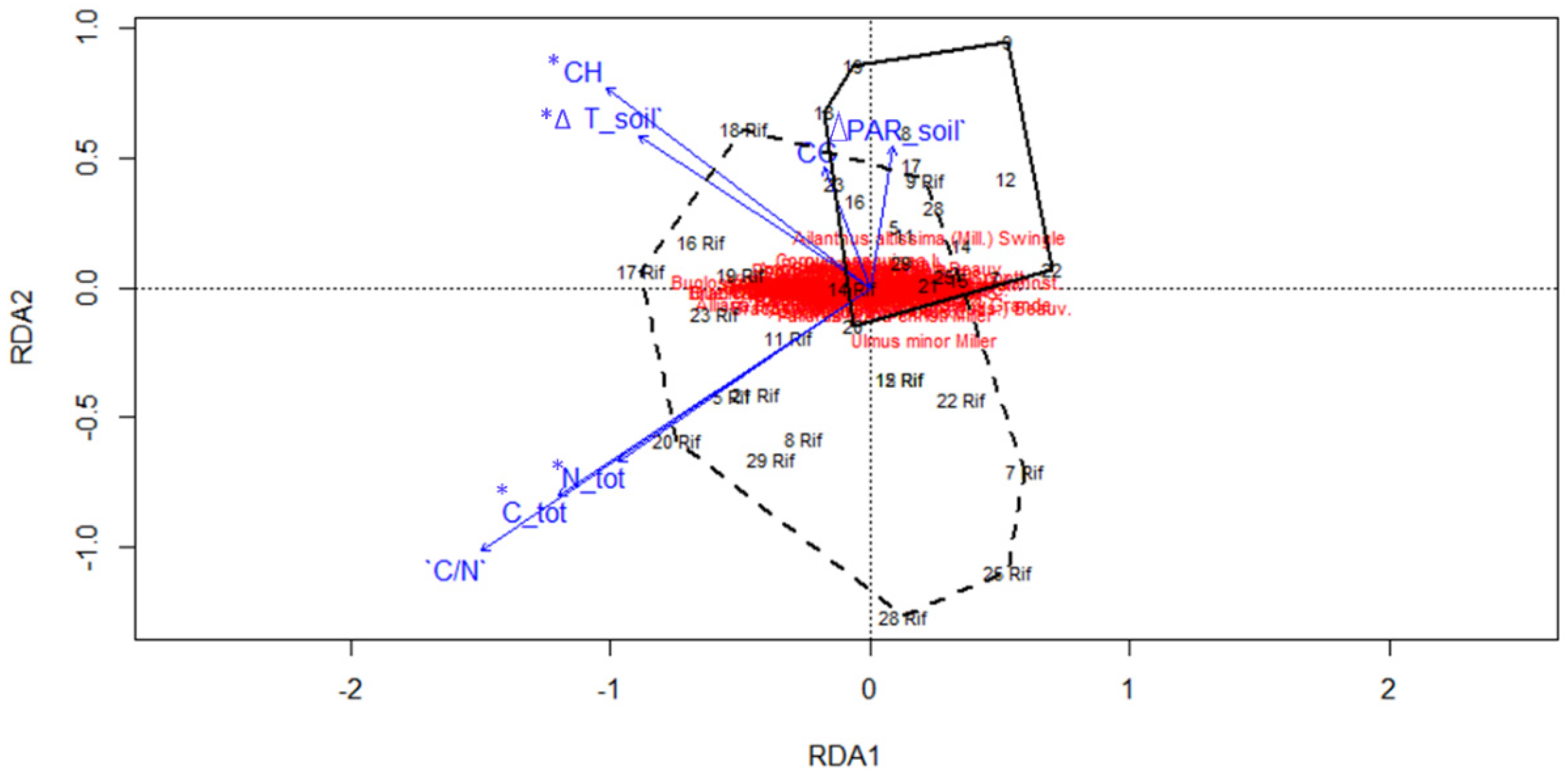
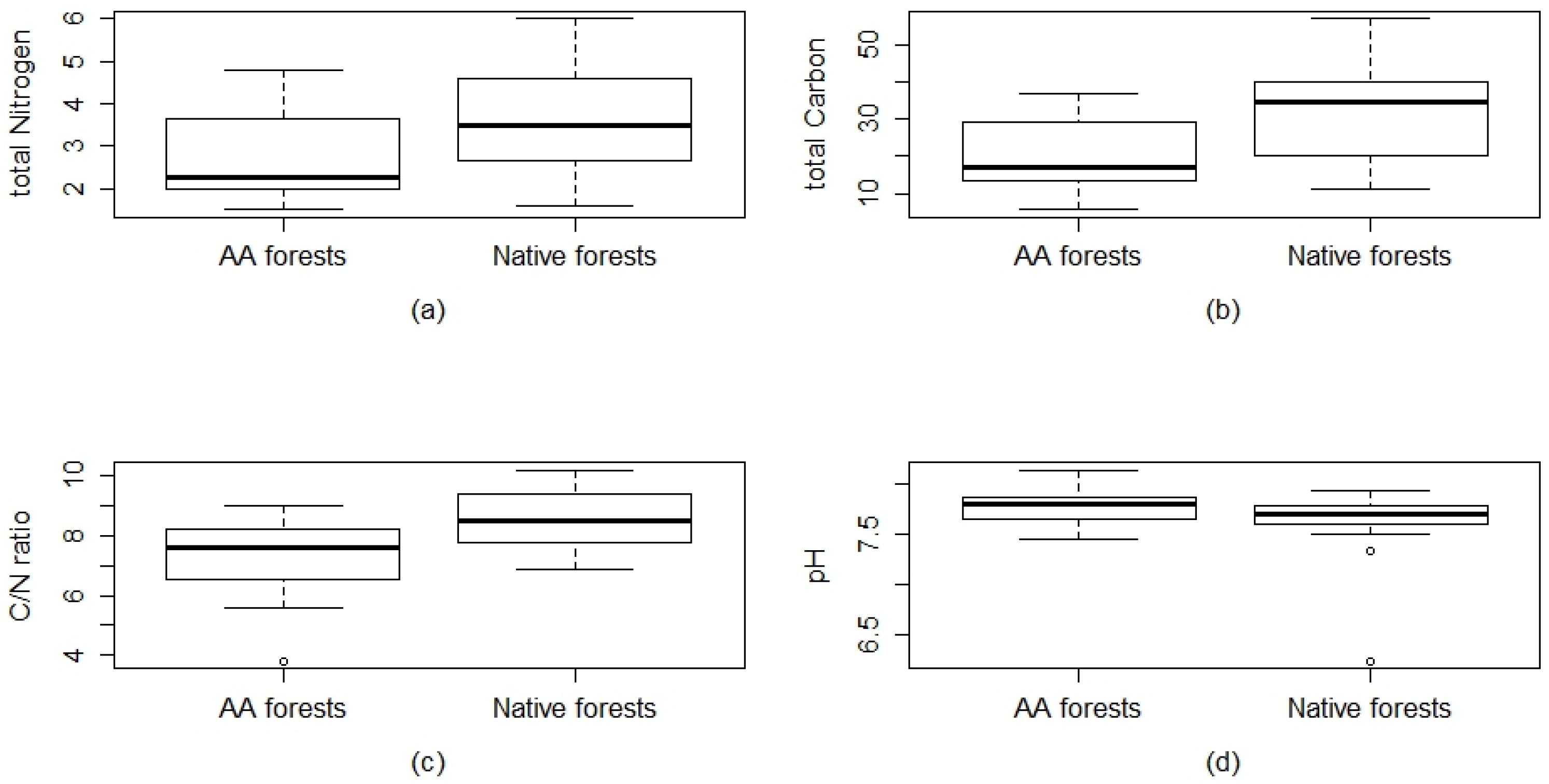
2.3. Ecological Variables
3. Discussion
3.1. Species Diversity, Life Forms, Grime Strategies
3.2. Ecological Variables
4. Materials and Methods
4.1. Study Area
4.2. Sampling Design
4.3. Vegetation Survey
4.4. Sampling of the Ecological Variables
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- van Kleunen, M.; Dawson, W.; Essl, F.; Jan, P.; Winter, M.; Weber, E.; Kreft, H.; Weigelt, P.; Kartesz, J.; Nishino, M.; et al. Global exchange and accumulation of non-native plants. Nature 2015, 525, 100–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabra-Rivas, I.; Saldaña, A.; Castro-Díez, P.; Gallien, L. A multi-scale approach to identify invasion drivers and invaders’ future dynamics. Biol. Invasions 2016, 18, 411–426. [Google Scholar] [CrossRef]
- Wagner, V.; Chytrý, M.; Jiménez-Alfaro, B.; Pergl, J.; Hennekens, S.; Biurrun, I.; Knollová, I.; Berg, C.; Vassilev, K.; Rodwell, J.S.; et al. Alien plant invasions in European woodlands. Divers. Distrib. 2017, 23, 969–981. [Google Scholar] [CrossRef]
- Perrings, C.; Naeem, S.; Ahrestani, F.; Bunker, D.E.; Burkill, P.; Canziani, G.; Elmqvist, T.; Ferrati, R.; Fuhrman, J.; Jaksic, F.; et al. Ecosystem Services for 2020. Science 2010, 330, 323–324. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Regulation (EU) No 1143/2014 of the European Parliament and of the Council of 22 October 2014 on the prevention and management of the introduction and spread of invasive alien species. Off. J. Eur. Union 2014, 57, 35–55. [Google Scholar]
- Lambdon, P.W.; Pyšek, P.; Basnou, C.; Hejda, M.; Arianoutsou, M.; Essl, F.; Jarošík, V.; Pergl, J.; Winter, M.; Anastasiu, P.; et al. Alien flora of Europe: Species diversity, temporal trends, geographical patterns and research needs. Preslia 2008, 80, 101–149. [Google Scholar]
- European Commission. Commission Implementing Regulation (EU) 2019/1262 of 25 July 2019 amending Implementing Regulation (EU) 2016/1141 to update the list of invasive alien species of Union concern. 2019. Available online: https://eur-lex.europa.eu/legal (accessed on 6 June 2020).
- Hu, S.H. Ailanthus. Arnoldia 1979, 39, 29–50. [Google Scholar]
- Gutte, P.; Klotz, S.; Lahr, C.; Trefflich, A. Ailanthus altissima (Mill.) Swingle. A comparative study of plant geography (German). Folia Geobot. Phytotaxon. 1987, 22, 241–262. [Google Scholar] [CrossRef]
- Udvardy, L. Spreading and Coenological circumstances of the tree of heaven (Ailanthus altissima) in Hungary. Acta Bot. Hung. 1998, 41, 229–314. [Google Scholar]
- Howard, T.G.; Gurevitch, J.; Hyatt, L.; Carreiro, M.; Lerdau, M. Forest invasibility in communities in southeastern New York. Biol. Invasions 2004, 6, 393–410. [Google Scholar] [CrossRef]
- Badalamenti, E.; Barone, E.; Salvatore, P.; Sala, G. Ailanthus altissima (Mill.) Swingle (Simaroubaceae) in Sicily and historical notes on its introduction in Italy (Italian). Nat. Sicil. 2012, 36, 117–164. [Google Scholar]
- Galasso, G.; Conti, F.; Peruzzi, L.; Ardenghi, N.M.G.; Banfi, E.; Celesti-Grapow, L.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S.; et al. An updated checklist of the vascular flora alien to Italy. Plant Biosyst. 2018, 152, 556–592. [Google Scholar] [CrossRef]
- Viciani, D.; Vidali, M.; Gigante, D.; Bolpagni, R.; Villani, M.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; Angiolini, C.; et al. A first checklist of the alien-dominated vegetation in Italy. Plant Sociol. 2020, 57, 29–54. [Google Scholar] [CrossRef]
- Lazzaro, L.; Bolpagni, R.; Buffa, G.; Gentili, R.; Lonati, M.; Stinca, A.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; et al. Impact of invasive alien plants on native plant communities and Natura 2000 habitats: State of the art, gap analysis and perspectives in Italy. J. Environ. Manag. 2020, 274, 111140. [Google Scholar] [CrossRef] [PubMed]
- Knapp, L.B.; Canham, C.D. Invasion of an old-growth forest in New York by Ailanthus altissima: Sapling growth and recruitment in canopy gaps. J. Torrey Bot. Soc. 2000, 127, 307–315. [Google Scholar] [CrossRef]
- Motard, E.; Muratet, A.; Clair-Maczulajtys, D.; MacHon, N. Does the invasive species Ailanthus altissima threaten floristic diversity of temperate peri-urban forests? Comptes Rendus Biol. 2011, 334, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Kowarik, I. Clonal growth in Ailanthus altissima. J. Veg. Sci. 1995, 6, 853–856. [Google Scholar] [CrossRef]
- Kowarik, I.; Säumel, I. Biological flora of Central Europe: Ailanthus altissima (Mill.) Swingle. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 207–237. [Google Scholar] [CrossRef]
- von der Lippe, M.; Bullock, J.M.; Kowarik, I.; Knopp, T.; Wichmann, M. Human-Mediated Dispersal of Seeds by the Airflow of Vehicles. PLoS ONE 2013, 8, e52733. [Google Scholar] [CrossRef]
- Fotiadis, G.; Kyriazopoulos, A.P.; Fraggakis, I. The behaviour of Ailanthus altissima weed and its effects on natural ecosystems. J. Environ. Biol. 2011, 32, 801–806. [Google Scholar]
- Höfle, R.; Dullinger, S.; Essl, F. Different factors affect the local distribution, persistence and spread of alien tree species in floodplain forests. Basic Appl. Ecol. 2014, 15, 426–434. [Google Scholar] [CrossRef]
- Costàn-Nava, S.; Soliveres, S.; Torices, R.; Serra, L.; Bonet, A. Direct and indirect effects of invasion by the alien tree Ailanthus altissima on riparian plant communities and ecosystem multifunctionality. Biol. Invasions 2015, 17, 1095–1108. [Google Scholar] [CrossRef]
- Lawrence, J.G.; Colwell, A.; Sexton, O.J. The ecological impact of allelopathy in Ailanthus altissima (Simaroubaceae). Am. J. Bot. 1991, 78, 948–958. [Google Scholar] [CrossRef]
- Bostan, C.; Borlea, F.; Mihoc, C.; Selesan, M. Ailanthus altissima species invasion on biodiversity caused by potential allelopathy. Res. J. Agric. Sci. 2014, 46, 95–103. [Google Scholar]
- Vilà, M.; Tessier, M.; Suehs, C.M.; Brundu, G.; Carta, L.; Galanidis, A.; Lambdon, P.; Manca, M.; Médail, F.; Moragues, E.; et al. Local and regional assessments of the impacts of plant invaders on vegetation structure and soil properties of Mediterranean islands. J. Biogeogr. 2006, 33, 853–861. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Canham, C.D. Neighborhood models of the effects of invasive tree species on ecosystem processes. Ecol. Monogr. 2008, 78, 69–86. [Google Scholar] [CrossRef] [Green Version]
- Castro-Díez, P.; González-Muñoz, N.; Alonso, A.; Gallardo, A.; Poorter, L. Effects of exotic invasive trees on nitrogen cycling: A case study in Central Spain. Biol. Invasions 2009, 11, 1973–1986. [Google Scholar] [CrossRef]
- Medina-Villar, S.; Castro-Díez, P.; Alonso, A.; Cabra-Rivas, I.; Parker, I.M.; Pérez-Corona, E. Do the invasive trees, Ailanthus altissima and Robinia pseudoacacia, alter litterfall dynamics and soil properties of riparian ecosystems in Central Spain? Plant Soil 2015, 396, 311–324. [Google Scholar] [CrossRef]
- Motard, E.; Dusz, S.; Geslin, B.; Akpa-Vinceslas, M.; Hignard, C.; Babiar, O.; Clair-Maczulajtys, D.; Michel-Salzat, A. How invasion by Ailanthus altissima transforms soil and litter communities in a temperate forest ecosystem. Biol. Invasions 2015, 17, 1817–1832. [Google Scholar] [CrossRef]
- Allegrezza, M.; Montecchiari, S.; Ottaviani, C.; Pelliccia, V.; Tesei, G. Syntaxonomy of the Robinia pseudoacacia communities in the central peri-adriatic sector of the Italian peninsula. Plant Biosyst. 2019, 153, 616–623. [Google Scholar] [CrossRef]
- Terwei, A.; Zerbe, S.; Mölder, I.; Annighöfer, P.; Kawaletz, H.; Ammer, C. Response of floodplain understorey species to environmental gradients and tree invasion: A functional trait perspective. Biol. Invasions 2016, 18, 2951–2973. [Google Scholar] [CrossRef]
- Constan Nava, S.; Bonet Jornet, A.; Serra Laliga, L. Efectos de la especie invasora Ailanthus altissima (mill.) swingle sobre la diversidad vegetal en bosques de ribera del lic serra de mariola y carrascal de la font roja. Iberis 2008, 6, 65–75. [Google Scholar]
- Grime, J.P. Plant Strategies, Vegetation Processes and Ecosystem Properties, 2nd ed.; John Wiley & Sons Ltd.: Chichester, UK, 2001. [Google Scholar]
- Montecchiari, S.; Tesei, G.; Allegrezza, M. Effects of Robinia pseudoacacia coverage on diversity and environmental conditions of central-northern Italian Quercus pubescens sub-mediterranean forests (habitat code 91AA*): A threshold assessment. Ann. Bot. Coenol. Plant 2020, 10, 33–54. [Google Scholar] [CrossRef]
- Gentili, R.; Ferrè, C.; Cardarelli, E.; Montagnani, C.; Bogliani, G.; Citterio, S.; Comolli, R. Comparing negative impacts of Prunus serotina, Quercus rubra and Robinia pseudoacacia on native forest ecosystems. Forests 2019, 10, 842. [Google Scholar] [CrossRef] [Green Version]
- Medina-Villar, S.; Rodríguez-Echeverría, S.; Lorenzo, P.; Alonso, A.; Pérez-Corona, E.; Castro-Díez, P. Impacts of the alien trees Ailanthus altissima (Mill.) Swingle and Robinia pseudoacacia L. on soil nutrients and microbial communities. Soil Biol. Biochem. 2016, 96, 65–73. [Google Scholar] [CrossRef]
- Castro-Díez, P.; Fierro-Brunnenmeister, N.; González-Muñoz, N.; Gallardo, A. Effects of exotic and native tree leaf litter on soil properties of two contrasting sites in the Iberian Peninsula. Plant Soil 2012, 350, 179–191. [Google Scholar] [CrossRef]
- Liao, C.; Peng, R.; Luo, Y.; Zhou, X.; Wu, X.; Fang, C.; Chen, J.; Li, B. Altered ecosystem carbon and nitrogen cycles by plant invasion: A meta-analysis. New Phytol. 2008, 177, 706–714. [Google Scholar] [CrossRef]
- Lazzaro, L.; Giuliani, C.; Fabiani, A.; Agnelli, A.E.; Pastorelli, R.; Lagomarsino, A.; Benesperi, R.; Calamassi, R.; Foggi, B. Soil and plant changing after invasion: The case of Acacia dealbata in a Mediterranean ecosystem. Sci. Total Environ. 2014, 497–498, 491–498. [Google Scholar] [CrossRef]
- Cools, N.; Vesterdal, L.; De Vos, B.; Vanguelova, E.; Hansen, K. Tree species is the major factor explaining C:N ratios in European forest soils. For. Ecol. Manag. 2014, 311, 3–16. [Google Scholar] [CrossRef]
- Knüsel, S.; De Boni, A.; Conedera, M.; Schleppi, P.; Thormann, J.J.; Frehner, M.; Wunder, J. Shade tolerance of Ailanthus altissima revisited: Novel insights from southern Switzerland. Biol. Invasions 2017, 19, 455–461. [Google Scholar] [CrossRef]
- Pesaresi, S.; Galdenzi, D.; Biondi, E.; Casavecchia, S. Bioclimate of Italy: Application of the worldwide bioclimatic classification system. J. Maps 2014, 10, 538–553. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Rivas-Sáenz, S.; Penas, A. Worldwide bioclimatic classification system. Glob. Geobot. 2011, 1, 1–634. [Google Scholar]
- Büttner, G.; Kosztra, B.; Soukup, T.; Sousa, A.; Langanke, T. Corine Land Cover 2018; Technical Guidelines; European Environment Agency: Wien, Austria, 2017. [Google Scholar]
- Blasi, C. (Ed.) Vegetation of Italy; Palombi & Partner S.r.l.: Rome, Italy, 2010. (In Italian) [Google Scholar]
- Bazalová, D.; Botková, K.; Hegedüšová, K.; Májeková, J.; Medvecká, J.; Šibíková, M.; Škodová, I.; Zaliberová, M.; Jarolímek, I. Twin plots-appropriate method to assess the impact of alien tree on understory? Hacquetia 2018, 17, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Regione Marche, IPLA Inventory and Forest Map of the Marche Region-The Forest Types of the Marche; IPLA S.p.A.: Torino, Italy, 2001; pp. 1–247. (In Italian)
- Deus, E.; Silva, J.S.; Catry, F.X.; Rocha, M.; Moreira, F. Google Street View as an alternative method to car surveys in large-scale vegetation assessments. Environ. Monit. Assess. 2016, 188, 1–14. [Google Scholar] [CrossRef]
- Sitzia, T.; Campagnaro, T.; Dainese, M.; Cierjacks, A. Plant species diversity in alien black locust stands: A paired comparison with native stands across a north-Mediterranean range expansion. For. Ecol. Manag. 2012, 285, 85–89. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Plant Sociology. Basics of Vegetation Science; Springer: Berlin/Heidelberg, Germany, 1928. (In German) [Google Scholar]
- Braun-Blanquet, J. Plant Sociology. Basics of Vegetation Science, 3rd ed.; Springer: New York, NY, USA, 1964; 865p. (In German) [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Pignatti, S. Flora of Italy; Edagricole: Bologna, Italy, 1982; Volume 1–3. (In Italian) [Google Scholar]
- Kühn, I.; Durka, W.; Klotz, S. BiolFlor—A new plant-trait database as a tool for plant invasion ecology. Divers. Distrib. 2004, 10, 363–365. [Google Scholar] [CrossRef]
- Pladias—Database of the Czech Flora and Vegetation. Available online: www.pladias.cz (accessed on 6 June 2020).
- Hunt, R.; Hodgson, J.G.; Thompson, K.; Bungener, P.; Dunnett, N.P.; Askew, A.P. A new practical tool for deriving a functional signature for herbaceous vegetation. Appl. Veg. Sci. 2004, 7, 163–170. [Google Scholar] [CrossRef]
- Astuti, G.; Ciccarelli, D.; Roma-Marzio, F.; Trinco, A.; Peruzzi, L. Narrow endemic species Bellevalia webbiana shows significant intraspecific variation in tertiary CSR strategy. Plant Biosyst. 2019, 153, 12–18. [Google Scholar] [CrossRef]
- Benhamiche-Hanifi, S.; Moulaï, R. Analysis of phytocenoses of island systems in the regions of Bejaia and Jijel (Algeria) in the presence of the white-backed gull (Larus michahellis) (French). Rev. Ecol. Terre Vie 2012, 67, 375–397. [Google Scholar]
- Bocchieri, E.; Iiriti, G. A contribution to the knowledge of the vascular flora of the continental carbonate formations situated on the Central Eastern margins of the Campidano plain (Southern Sardinia). Flora Mediterr. 2006, 16, 145–168. [Google Scholar]
- Biondi, E.; Blasi, C.; Allegrezza, M.; Anzellotti, I.; Azzella, M.M.; Carli, E.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Facioni, L.; et al. Plant communities of Italy: The Vegetation Prodrome. Plant Biosyst. 2014, 148, 728–814. [Google Scholar] [CrossRef] [Green Version]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Tichý, L. GLAMA-Gap Light Analysis Mobile Application; Department of Botany and Zoology: Brno, Czech Republic, 2014. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B. Vegan: Community Ecology Package. Package version 2.5-1. 2018. Available online: https://cran.r-project.org/web/packages/vegan/ (accessed on 6 June 2020).
- R Development Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. Available online: http://www.r-project.org (accessed on 6 June 2020).
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ailanthus Forests | Native Forests | Ailanthus Vs. Native | |||
|---|---|---|---|---|---|
| Mean | Mean | Mean sq | p-Value | ||
| Species richness | tree layer | 2.42 | 2.95 | 2.63 | 0.18 |
| Species richness | shrub layer | 7.16 | 8.84 | 13.92 | 0.25 |
| Species richness | herb-layer | 8.16 | 7.74 | 50.95 | 0.05 |
| Shannon index | tree layer | 0.68 | 0.89 | 0.41 | 0.18 |
| Shannon index | shrub layer | 1.81 | 2.00 | 0.34 | 0.19 |
| Shannon index | herb-layer | 2.13 | 1.91 | 0.45 | 0.16 |
| Ecological Variables | df | Variance | F | p | |
|---|---|---|---|---|---|
| N_tot | 1 | 0.02694 | 1.69 | 0.012 | * |
| C_tot | 1 | 0.03234 | 2.03 | 0.001 | *** |
| C/N | 1 | 0.01138 | 0.72 | 0.896 | |
| CC | 1 | 0.01431 | 0.90 | 0.596 | |
| CH | 1 | 0.03037 | 1.91 | 0.005 | ** |
| ΔPAR_soil | 1 | 0.01163 | 0.73 | 0.891 | |
| ΔT_soil | 1 | 0.02625 | 1.65 | 0.020 | * |
| Residual | 30 | 0.47722 |
| Topographic and Ecological Variables | Unit | Symbol |
|---|---|---|
| Altitude | m a.s.l. | Alt |
| Slope | ° | Sl |
| Northness | ° | N |
| Canopy Cover | % | CC |
| Canopy Height | m | CH |
| Photosynthetic active radiation at 1.30 m from the soil | (µmol m−2 s−1) | PAR_chest |
| Photosynthetic active radiation at the soil level | (µmol m−2 s−1) | PAR_soil |
| Photosynthetic active radiation in full light condition | (µmol m−2 s−1) | PAR_out |
| Temperature of the topsoil under forest canopy | °C | T_in |
| Temperature of the topsoil outside forest canopy | °C | T_out |
| Temperature of the air | °C | T_air |
| Difference PAR_out-PAR_chest | Δ PAR_chest | |
| Difference PAR_out-PAR_soil | Δ PAR_soil | |
| Difference T_out-T_in | Δ T soil | |
| Total nitrogen | g/kg | N_tot |
| Total extractable carbon | g/kg | C_tot |
| carbon-nitrogen ratio | g/kg | C/N |
| pH in H2O | pH unit | pH |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montecchiari, S.; Tesei, G.; Allegrezza, M. Ailanthus altissima Forests Determine a Shift in Herbaceous Layer Richness: A Paired Comparison with Hardwood Native Forests in Sub-Mediterranean Europe. Plants 2020, 9, 1404. https://doi.org/10.3390/plants9101404
Montecchiari S, Tesei G, Allegrezza M. Ailanthus altissima Forests Determine a Shift in Herbaceous Layer Richness: A Paired Comparison with Hardwood Native Forests in Sub-Mediterranean Europe. Plants. 2020; 9(10):1404. https://doi.org/10.3390/plants9101404
Chicago/Turabian StyleMontecchiari, Silvia, Giulio Tesei, and Marina Allegrezza. 2020. "Ailanthus altissima Forests Determine a Shift in Herbaceous Layer Richness: A Paired Comparison with Hardwood Native Forests in Sub-Mediterranean Europe" Plants 9, no. 10: 1404. https://doi.org/10.3390/plants9101404