Overexpression of CDSP32 (GhTRX134) Cotton Gene Enhances Drought, Salt, and Oxidative Stress Tolerance in Arabidopsis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. qRT-PCR Assays
2.3. Gene Cloning and Sequence Analysis
2.4. GhTRX134 Transgenic Arabidopsis Plants
2.4.1. Construction of the GhTRX134 Plasmids and Transformation of Arabidopsis
2.4.2. Growth of Transgenic Plants under Stress Treatments
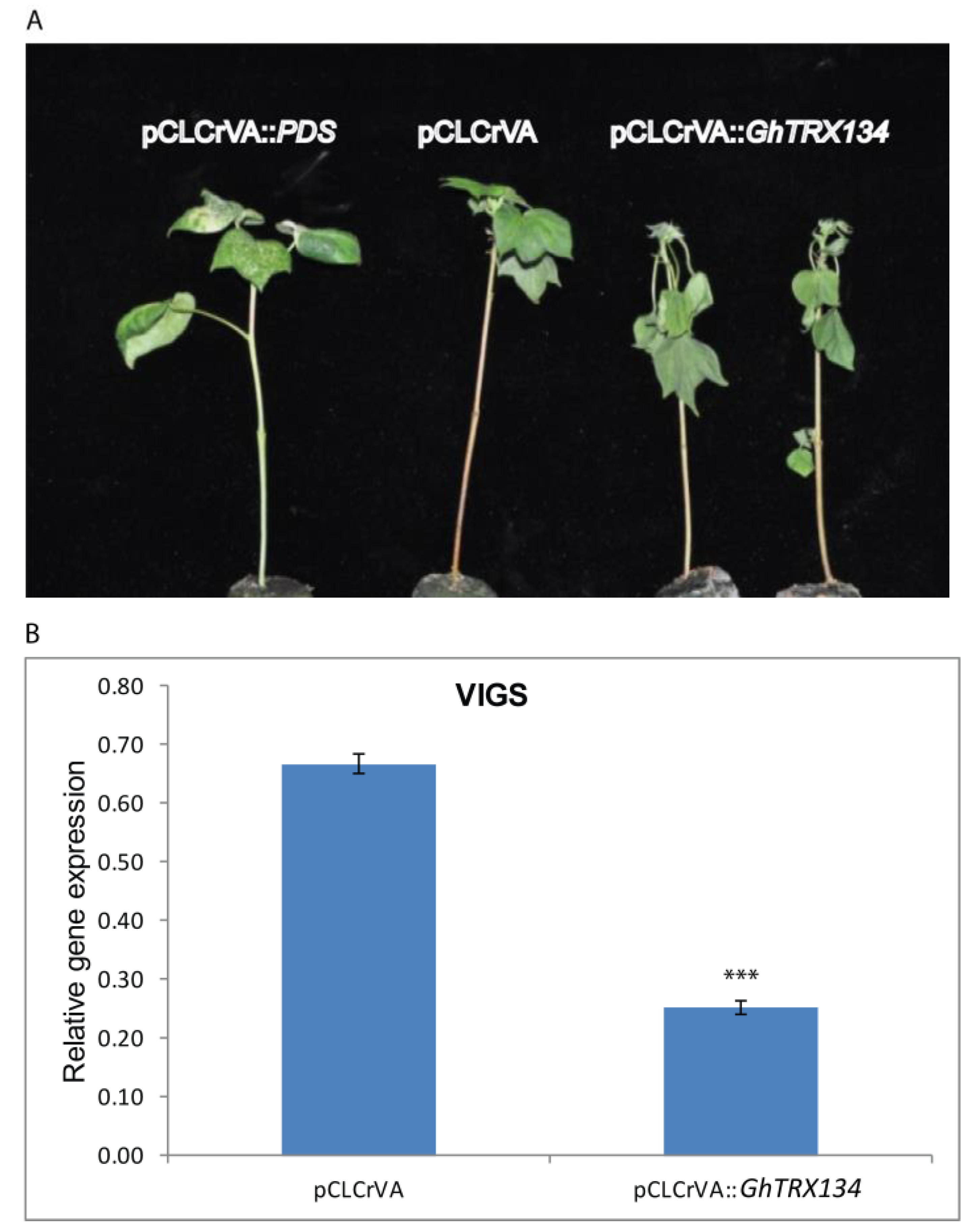
2.5. Plasmid Construction for VIGS Assay and Vector Inoculation
2.6. Analysis of Promoter Cis-Acting Elements
2.7. Trans-Activation Analysis of GhTRX134
2.8. Expression of Stress-Responsive Genes in Transgenic and Wt Plants under Abiotic Stress
2.9. Measurements of Proline Content, Enzyme Activity, and Chlorophyll Content
2.10. Statistical Analysis
3. Results
3.1. Cloning and Characterization of GhTRX134 (CDSP32)
3.2. Expression and Phenotypic Analysis of GhTRX134 Overexpression during Abiotic Stresses
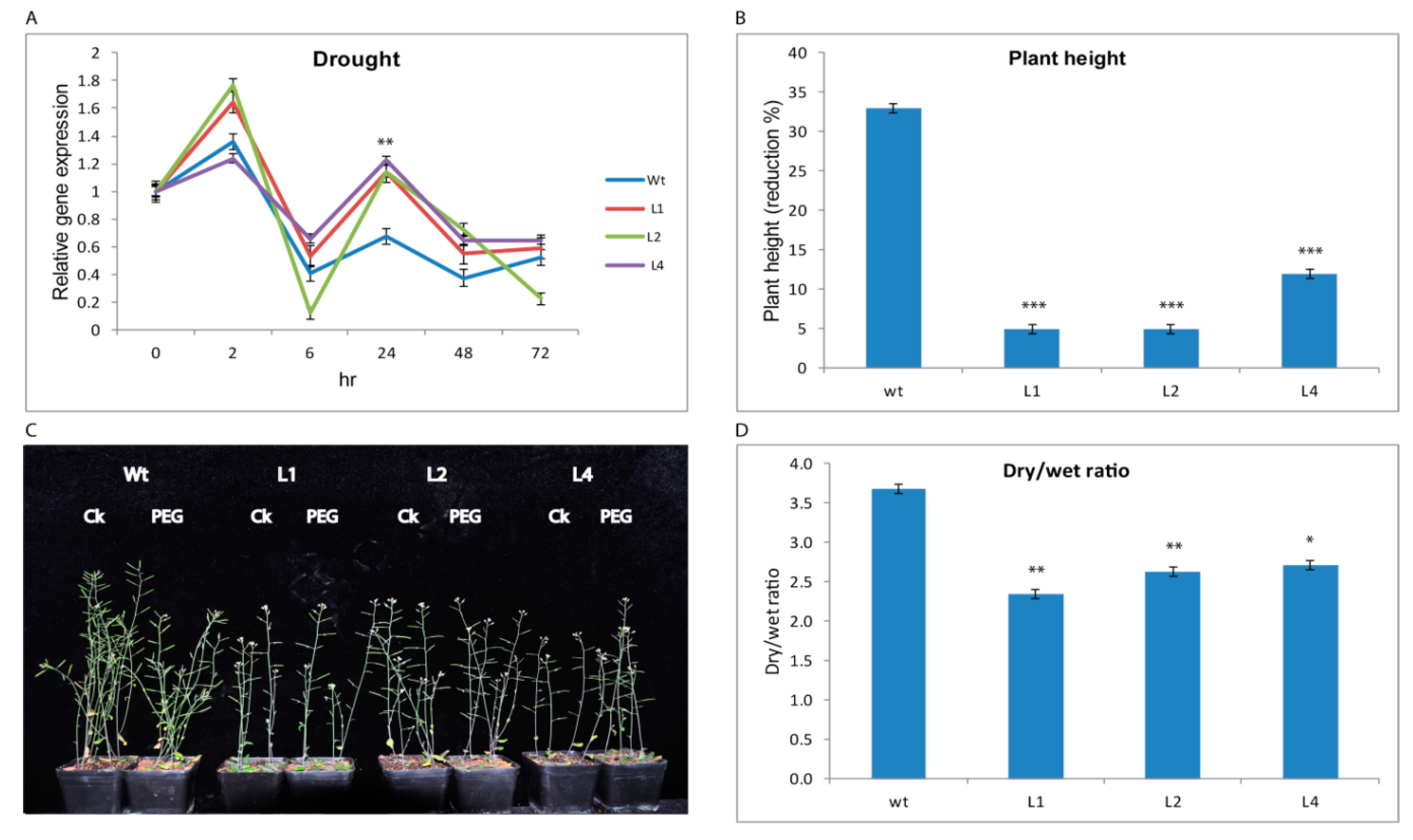
3.2.1. GhTRX134 Overexpression Enhances Drought Tolerance
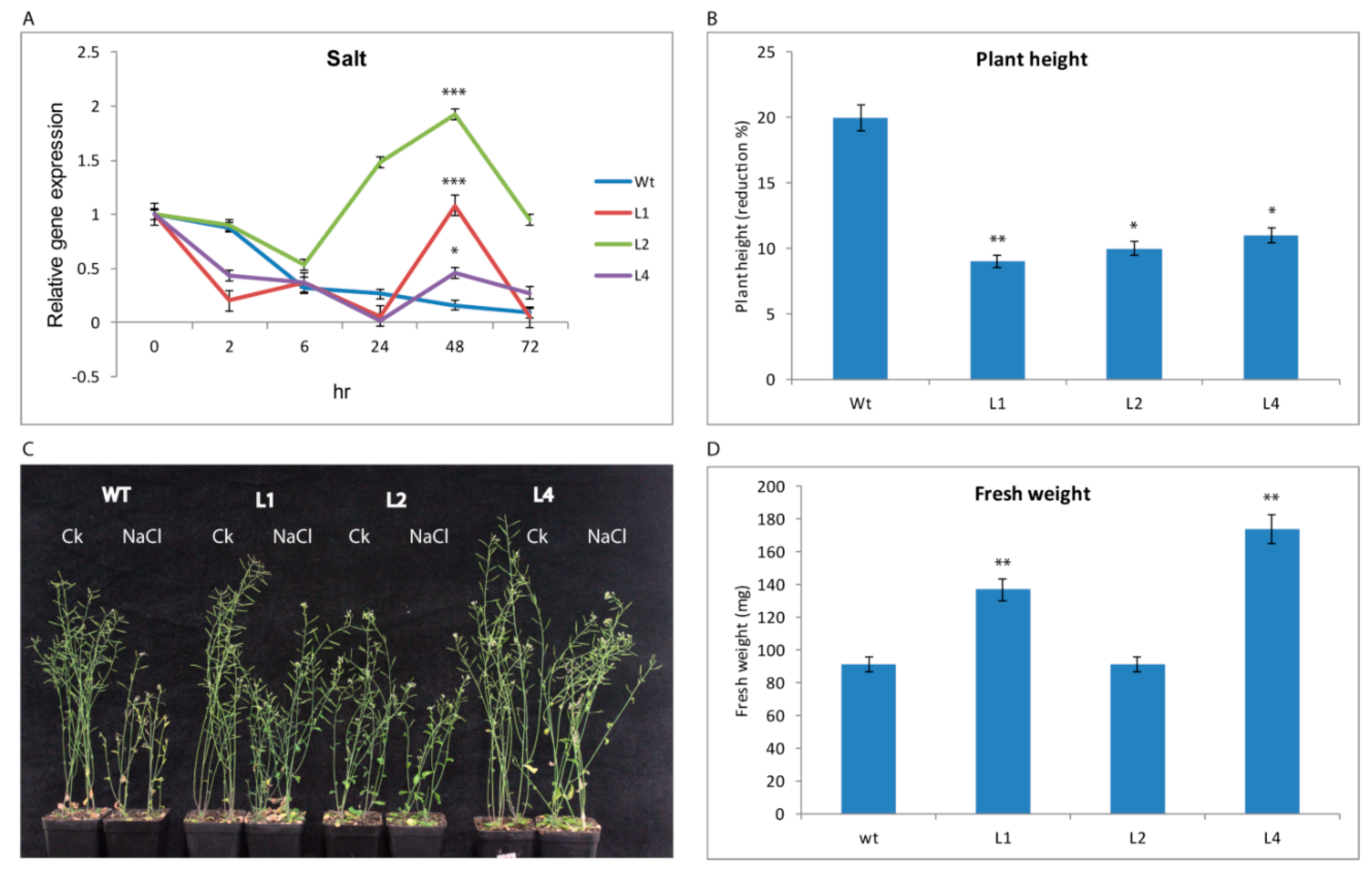
3.2.2. GhTRX134 Overexpression Confers Salt Tolerance
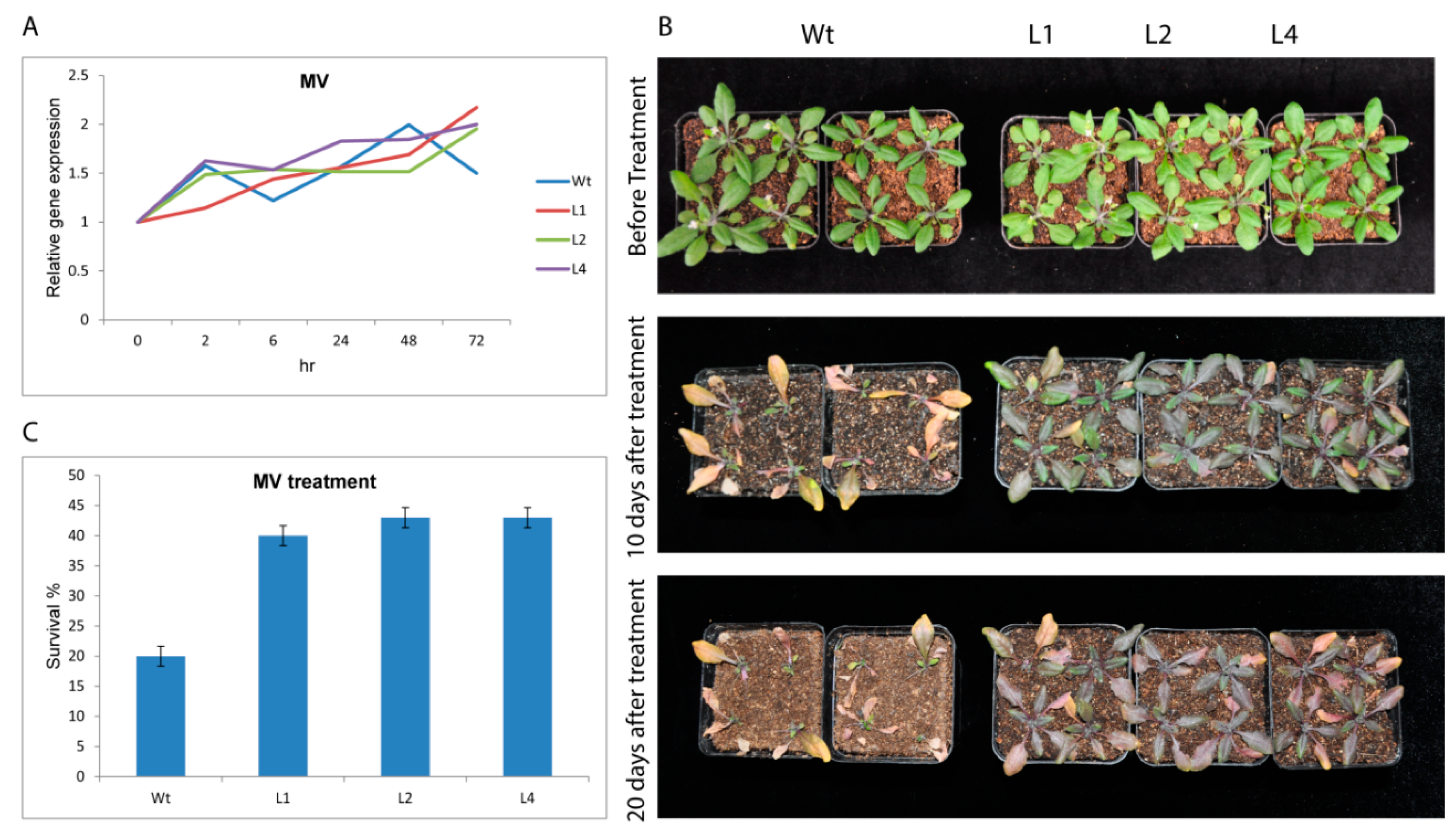
3.2.3. GhTRX134 Overexpression Improves Oxidative Resistance
3.3. GhTRX134 Plays a Positive Role in Drought Stress Tolerence in Cotton Plants
3.4. In Silico Promoter Analysis
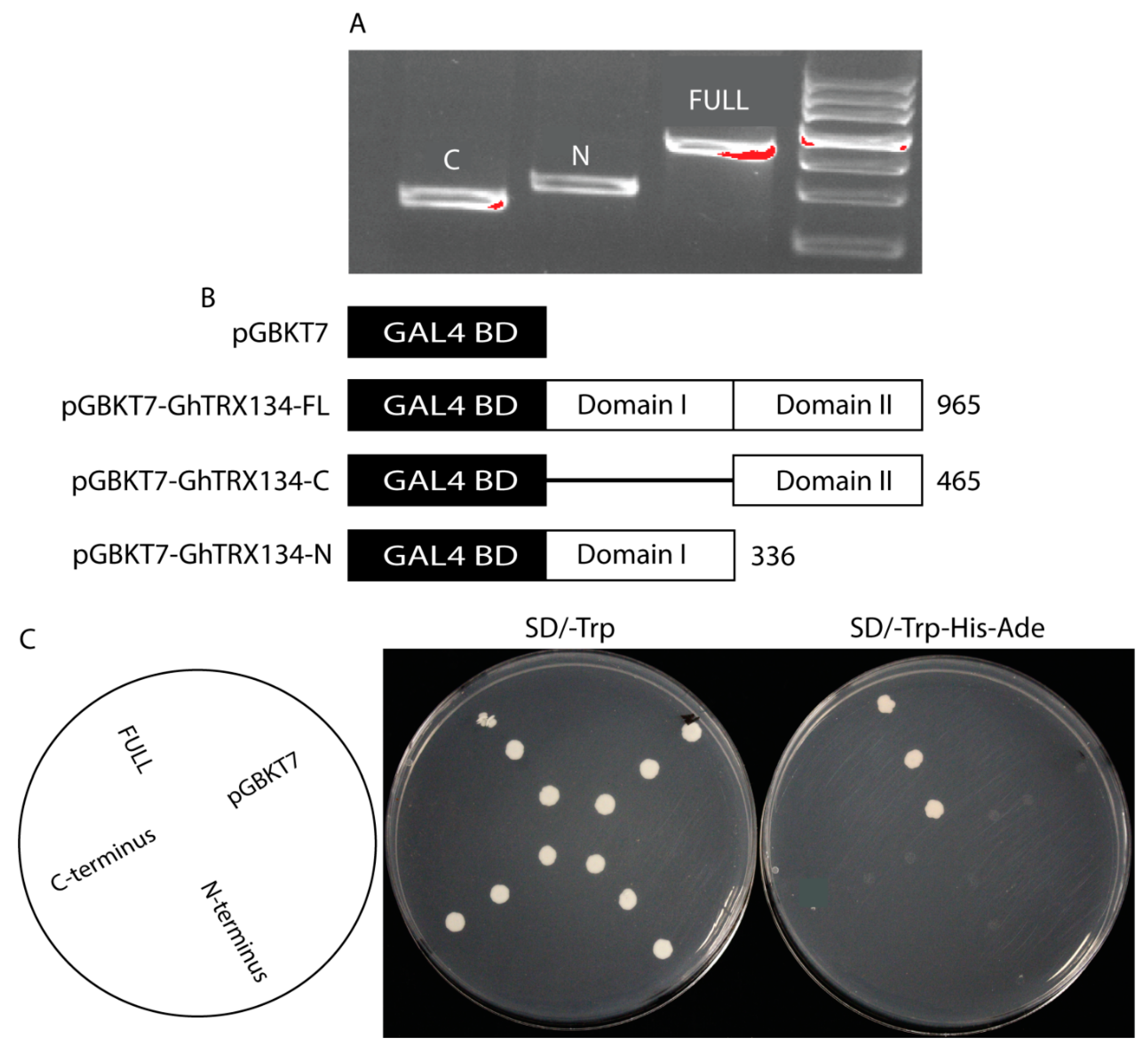
3.5. The GhTRX134 Gene Acts as an Activator
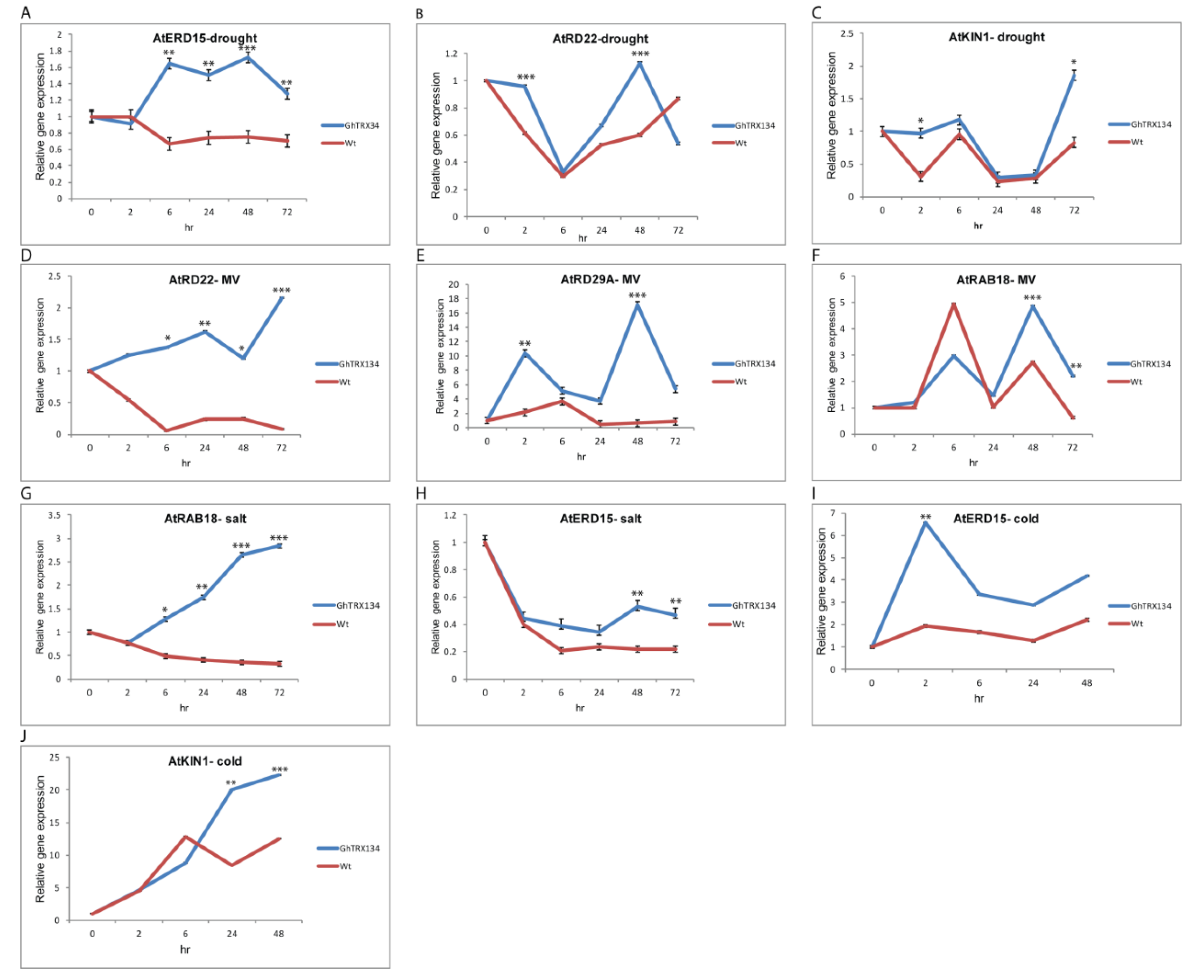
3.6. GhTRX134 Overexpression Regulates the Expression of Stress-Responsive Genes
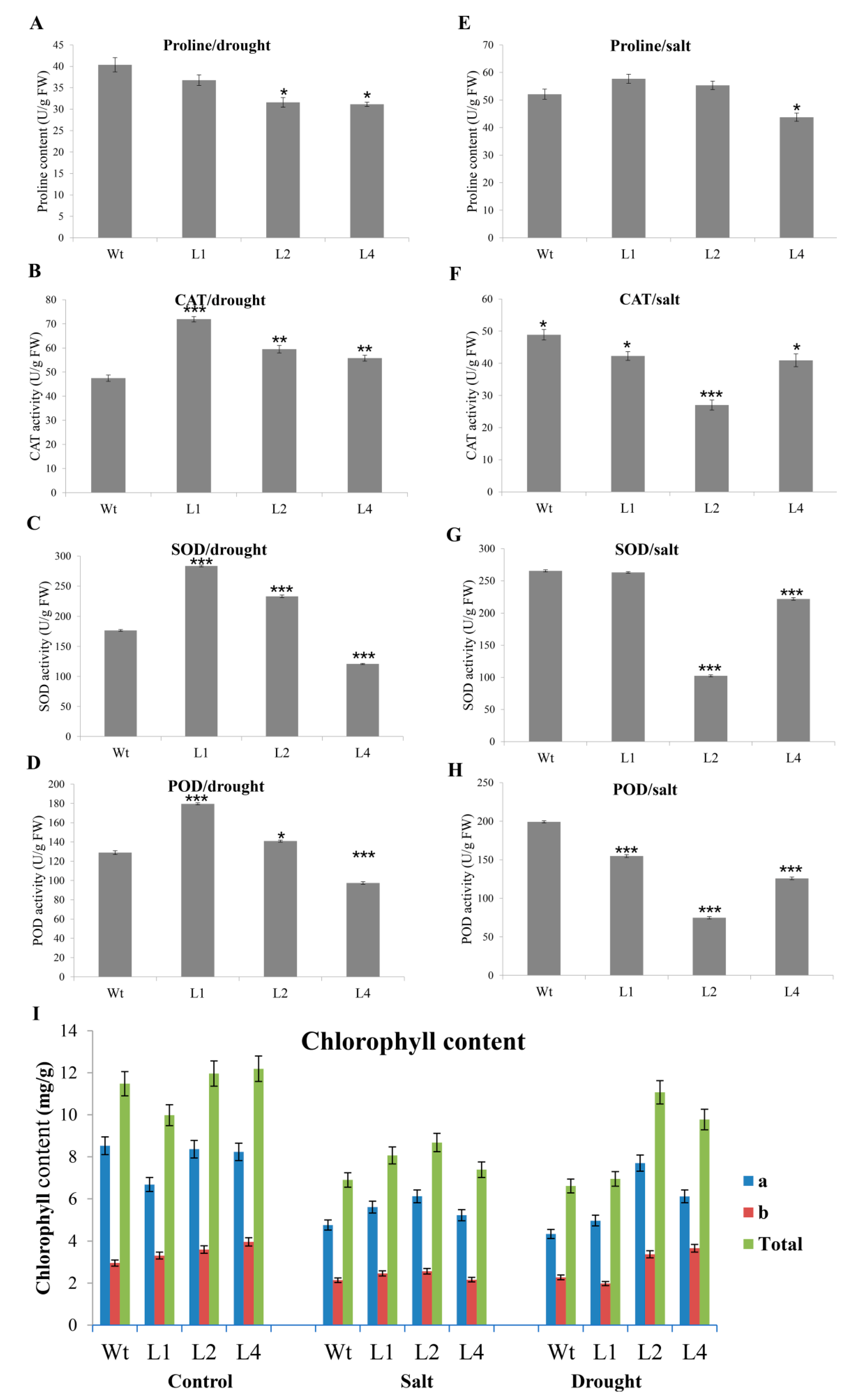
3.7. Measurements of Proline Content, Enzyme Activity, and Chlorophyll Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Wang, X.; Yu, Y.; Sang, J.; Wu, Q.; Zhang, X.; Lin, Z. Intraspecific linkage map construction and QTL mapping of yield and fiber quality of ‘Gossypiumbabrdense’. Aust. J. Crop. Sci. 2013, 7, 1252–1261. Available online: http://search.informit.com.au/documentSummary;dn=625250539555035;res=IELHSS (accessed on 3 March 2020).
- Elasad, M.; Wei, H.; Wang, H.; Su, J.; Ondati, E.; Yu, S. Genome-Wide Analysis and Characterization of the TRX Gene Family in Upland Cotton. Trop. Plant Biol. 2018, 11, 119–130. [Google Scholar] [CrossRef]
- Zhao, G.; Song, Y.; Wang, C.; Butt, H.I.; Wang, Q.; Zhang, C.; Yang, Z.; Liu, Z.; Chen, E.; Zhang, X.; et al. Genome-wide identification and functional analysis of the TIFY gene family in response to drought in cotton. Mol. Genet. Genom. 2016, 291, 2173–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, A.J.; Hell, R. Glutathione homeostasis and redox-regulation by sulfhydryl groups. Photosynth. Res. 2005, 86, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Colville, L.; Kranner, I. Desiccation tolerant plants as model systems to study redox regulation of protein thiols. Plant. Growth Regul. 2010, 62, 241–255. [Google Scholar] [CrossRef]
- Eklund, H.; Gleason, F.K.; Holmgren, A. Structural and functional relations among thioredoxins of different species. Proteins Struct. Funct. Genet. 1991, 11, 13–28. [Google Scholar] [CrossRef]
- Thormählen, I.; Meitzel, T.; Groysman, J.; Öchsner, A.B.; Von Roepenack-Lahaye, E.; Naranjo, B.; Cejudo, F.J.; Geigenberger, P. Thioredoxin f1 and NADPH-dependent thioredoxin reductase C have overlapping functions in regulating photosynthetic metabolism and plant growth in response to varying light conditions. Plant Physiol. 2015, 169, 1766–1786. [Google Scholar] [CrossRef] [Green Version]
- Chibani, K.; Wingsle, G.; Jacquot, J.-P.; Gelhaye, E.; Rouhier, N. Comparative Genomic Study of the Thioredoxin Family in Photosynthetic Organisms with Emphasis on Populus trichocarpa. Mol. Plant 2009, 2, 308–322. [Google Scholar] [CrossRef]
- Broin, M.; Rey, P. Potato Plants Lacking the CDSP32 Plastidic Thioredoxin Exhibit Overoxidation of the BAS1 2-Cysteine Peroxiredoxin and Increased Lipid Peroxidation in Thylakoids under Photooxidative Stress. Plant Physiol. 2003, 132, 1335–1343. [Google Scholar] [CrossRef] [Green Version]
- Meyer, Y.; Siala, W.; Bashandy, T.; Riondet, C.; Vignols, F.; Reichheld, J.P. Glutaredoxins and thioredoxins in plants. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2008, 1783, 589–600. [Google Scholar] [CrossRef]
- Ruiz, J.; Blumwald, E. Salinity-induced glutathione synthesis in Brassica napus. Planta 2002, 214, 965–969. [Google Scholar] [CrossRef]
- Harms, K.; Von Ballmoos, P.; Brunold, C.; Hofgen, R.; Hesse, H. Expression of a bacterial serine acetyltransferase in transgenic potato plants leads to increased levels of cysteine and glutathione. Plant J. 2000, 22, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuruzzaman, M.; Sharoni, A.M.; Satoh, K.; Al-Shammari, T.; Shimizu, T.; Sasaya, T.; Omura, T.; Kikuchi, S. The thioredoxin gene family in rice: Genome-wide identification and expression profiling under different biotic and abiotic treatments. Biochem. Biophys. Res. Commun. 2013, 423, 417–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Cui, J.; Zhang, H.; Wang, G.; Zhao, F.-J.; Shen, Z. Proteomic analysis of copper stress responses in the roots of two rice (Oryza sativa L.) varieties differing in Cu tolerance. Plant Soil 2012, 366, 647–658. [Google Scholar] [CrossRef]
- Chae, H.B.; Moon, J.C.; Shin, M.R.; Chi, Y.H.; Jung, Y.J.; Lee, S.Y.; Nawkar, G.M.; Jung, H.S.; Hyun, J.K.; Kim, W.-Y.; et al. Thioredoxin Reductase Type C (NTRC) Orchestrates Enhanced Thermotolerance to Arabidopsis by Its Redox-Dependent Holdase Chaperone Function. Mol. Plant 2013, 6, 323–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broin, M.; Cuiné, S.; Peltier, G.; Rey, P. Involvement of CDSP 32, a drought-induced thioredoxin, in the response to oxidative stress in potato plants. FEBS Lett. 2000, 467, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Takemoto, T.; Zhang, Q.; Yonei, S. Different Mechanisms of Thioredoxin in its Reduced and Oxidized Forms in Defense Against Hydrogen Peroxide in Escherichia coli. Free Radic. Biol. Med. 1998, 24, 556–562. [Google Scholar] [CrossRef]
- Dos Santos, C.V.; Rey, P. Plant thioredoxins are key actors in the oxidative stress response. Trends Plant Sci. 2006, 11, 329–334. [Google Scholar] [CrossRef]
- Lemaire, S.D.; Guillon, B.; Le Maréchal, P.; Keryer, E.; Miginiac-Maslow, M.; Decottignies, P. New thioredoxin targets in the unicellular photosynthetic eukaryote Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2004, 101, 7475–7480. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Liu, F.; Pang, C.; Fan, S.; Song, M.; Wang, D.; Li, W.; Yu, S. Label-Free Quantitative Proteomics Analysis of Cotton Leaf Response to Nitric Oxide. J. Proteome Res. 2011, 10, 5416–5432. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-H.; Ronald, P.C. A Rapid DNA Minipreparation Method Suitable for AFLP and Other PCR Applications. Plant Mol. Biol. Rep. 1999, 17, 53–57. [Google Scholar] [CrossRef]
- Bhuiyan, M.S.U.; Min, S.R.; Jeong, W.-J.; Sultana, S.; Choi, K.S.; Lim, Y.P.; Song, W.-Y.; Lee, Y.; Liu, J.R. An improved method for Agrobacterium-mediated genetic transformation from cotyledon explants of Brassica juncea. Plant Biotechnol. 2011, 28, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method forAgrobacterium-mediated transformation ofArabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Silencing, A.V.G. Virus-Induced Gene Silencing. Adv. Struct. Saf. Stud. 2013, 975, 1–8. [Google Scholar] [CrossRef]
- Gu, Z.; Huang, C.J.; Li, F.; Zhou, X. A versatile system for functional analysis of genes and microRNAs in cotton. Plant Biotechnol. J. 2014, 12, 638–649. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S. SHORT COMMUNICATION Rapid determination of free proline for water-stress studies. Plant Soil 1973, 207, 2104. [Google Scholar]
- Wang, Y.; Gao, C.; Liang, Y.; Wang, C.; Yang, C.; Liu, G. A novel bZIP gene from Tamarix hispida mediates physiological responses to salt stress in tobacco plants. J. Plant Physiol. 2010, 167, 222–230. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–383. [Google Scholar] [CrossRef]
- Pastori, G.M.; Foyer, C.H. Common Components, Networks, and Pathways of Cross-Tolerance to Stress. The Central Role of “Redox” and Abscisic Acid-Mediated Controls. Plant Physiol. 2002, 129, 460–468. [Google Scholar] [CrossRef] [Green Version]
- Chae, H.Z.; Chung, S.J.; Rhee, S.G. Thioredoxin-dependent peroxide reductase from yeast. J. Biol. Chem. 1994, 269, 27670–27678. [Google Scholar]
- Kasuga, M.; Liu, Q.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nat. Biotechnol. 1999, 17, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, S.J.; Sebolt, A.M.; Salazar, M.P.; Everard, J.D.; Thomashow, M.F. Overexpression of the Arabidopsis CBF3Transcriptional Activator Mimics Multiple Biochemical Changes Associated with Cold Acclimation. Plant Physiol. 2000, 124, 1854–1865. [Google Scholar] [CrossRef] [Green Version]
- Tarrago, L.; Laugier, E.; Zaffagnini, M.; Marchand, C.H.; Le Maréchal, P.; Lemaire, S.D.; Rey, P. Plant Thioredoxin CDSP32 Regenerates 1-Cys Methionine Sulfoxide Reductase B Activity through the Direct Reduction of Sulfenic Acid. J. Biol. Chem. 2010, 285, 14964–14972. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.-C.; Lin, W.-C.; Cheng, W.-H. Salt hypersensitive mutant 9, a nucleolar APUM23 protein, is essential for salt sensitivity in association with the ABA signaling pathway in Arabidopsis. BMC Plant Biol. 2018, 18, 40. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Takano, T.; Liu, S.; Zhang, X. Arabidopsis mitochondrial voltage-dependent anion channel 3 (AtVDAC3) protein interacts with thioredoxin m2. FEBS Lett. 2015, 589, 1207–1213. [Google Scholar] [CrossRef] [Green Version]
- Sunkar, R.; Bartels, D.; Kirch, H.-H. Overexpression of a stress-inducible aldehyde dehydrogenase gene from Arabidopsis thaliana in transgenic plants improves stress tolerance. Plant J. 2003, 35, 452–464. [Google Scholar] [CrossRef]
- Wang, L.; Li, Z.; Wang, C.; Wang, D.; Wang, Y.; Lu, M.-Z. Overexpression of a peroxiredoxin gene from Tamarix hispida, ThPrx1, confers tolerance to oxidative stress in yeast and Arabidopsis. J. Plant Biol. 2017, 60, 548–557. [Google Scholar] [CrossRef]
- Han, H.-J.; Peng, R.-H.; Zhu, B.; Fu, X.-Y.; Zhao, W.; Shi, B.; Yao, Q.-H. Gene expression profiles of arabidopsis under the stress of methyl viologen: A microarray analysis. Mol. Biol. Rep. 2014, 41, 7089–7102. [Google Scholar] [CrossRef]
- Vignols, F.; Bréhélin, C.; Surdin-Kerjan, Y.; Thomas, D.; Meyer, Y. A yeast two-hybrid knockout strain to explore thioredoxin-interacting proteins in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 16729–16734. [Google Scholar] [CrossRef] [Green Version]
- Elhiti, M.; Stasolla, C. Structure and function of homodomain-leucine zipper (HD-Zip) proteins. Plant Signal. Behav. 2009, 4, 86–88. [Google Scholar] [CrossRef] [Green Version]
- Shinde, H.; Dudhate, A.; Tsugama, D.; Gupta, S.K.; Liu, S.; Takano, T. Pearl millet stress-responsive NAC transcription factor PgNAC21 enhances salinity stress tolerance in Arabidopsis. Plant Physiol. Biochem. 2019, 135, 546–553. [Google Scholar] [CrossRef]
- McClung, C.R. Plant Circadian Rhythms. Plant Cell 2006, 18, 792–803. [Google Scholar] [CrossRef] [Green Version]
- Zeller, T.; Klug, G. Thioredoxins in bacteria: Functions in oxidative stress response and regulation of thioredoxin genes. Naturwissenschaften 2006, 93, 259–266. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef]
- Wu, D.; Sun, Y.; Wang, H.; Shi, H.; Su, M.; Shan, H.; Li, T.; Li, Q.-L. The SlNAC8 gene of the halophyte Suaeda liaotungensis enhances drought and salt stress tolerance in transgenic Arabidopsis thaliana. Gene 2018, 662, 10–20. [Google Scholar] [CrossRef]
- Chu, X.; Wang, C.; Chen, X.; Lu, W.; Li, H.; Wang, X.; Hao, L.; Guo, X. The cotton WRKY gene GhWRKY41 positively regulates salt and drought stress tolerance in transgenic Nicotiana benthamiana. PLoS ONE 2015, 10, e0143022. [Google Scholar] [CrossRef]
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotechnol. 2005, 16, 123–132. [Google Scholar] [CrossRef]
- Hmida-Sayari, A.; Gargouri-Bouzid, R.; Bidani, A.; Jaoua, L.; Savouré, A.; Jaoua, S. Overexpression of Δ1-pyrroline-5-carboxylate synthetase increases proline production and confers salt tolerance in transgenic potato plants. Plant Sci. 2005, 169, 746–752. [Google Scholar] [CrossRef]
- Zhang, H.X.; Blumwald, E. Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat. Biotechnol. 2001, 19, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y.; Wu, G.; Njeri, K.V.; Shen, Q.; Zhang, N.; Zhang, R. Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol. Plant. 2016, 158, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-Y.; Niu, H.-B.; Yin, J.; Shao, H.-B.; Niu, J.-S.; Ren, J.-P.; Li, Y.-C.; Wang, X. Transgenic barley with overexpressed PTrx increases aluminum resistance in roots during germination. J. Zhejiang Univ. Sci. B 2010, 11, 862–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey, P.; Pruvot, G.; Becuwe, N.; Eymery, F.; Rumeau, D.; Peltier, G. A novel thioredoxin-like protein located in the chloroplast is induced by water deficit in Solanum tuberosum L. plants. Plant J. 1998, 13, 97–107. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elasad, M.; Ahmad, A.; Wang, H.; Ma, L.; Yu, S.; Wei, H. Overexpression of CDSP32 (GhTRX134) Cotton Gene Enhances Drought, Salt, and Oxidative Stress Tolerance in Arabidopsis. Plants 2020, 9, 1388. https://doi.org/10.3390/plants9101388
Elasad M, Ahmad A, Wang H, Ma L, Yu S, Wei H. Overexpression of CDSP32 (GhTRX134) Cotton Gene Enhances Drought, Salt, and Oxidative Stress Tolerance in Arabidopsis. Plants. 2020; 9(10):1388. https://doi.org/10.3390/plants9101388
Chicago/Turabian StyleElasad, Mohammed, Adeel Ahmad, Hantao Wang, Liang Ma, Shuxun Yu, and Hengling Wei. 2020. "Overexpression of CDSP32 (GhTRX134) Cotton Gene Enhances Drought, Salt, and Oxidative Stress Tolerance in Arabidopsis" Plants 9, no. 10: 1388. https://doi.org/10.3390/plants9101388