High-Throughput Sequencing Application in the Diagnosis and Discovery of Plant-Infecting Viruses in Africa, A Decade Later



Abstract
:1. Introduction
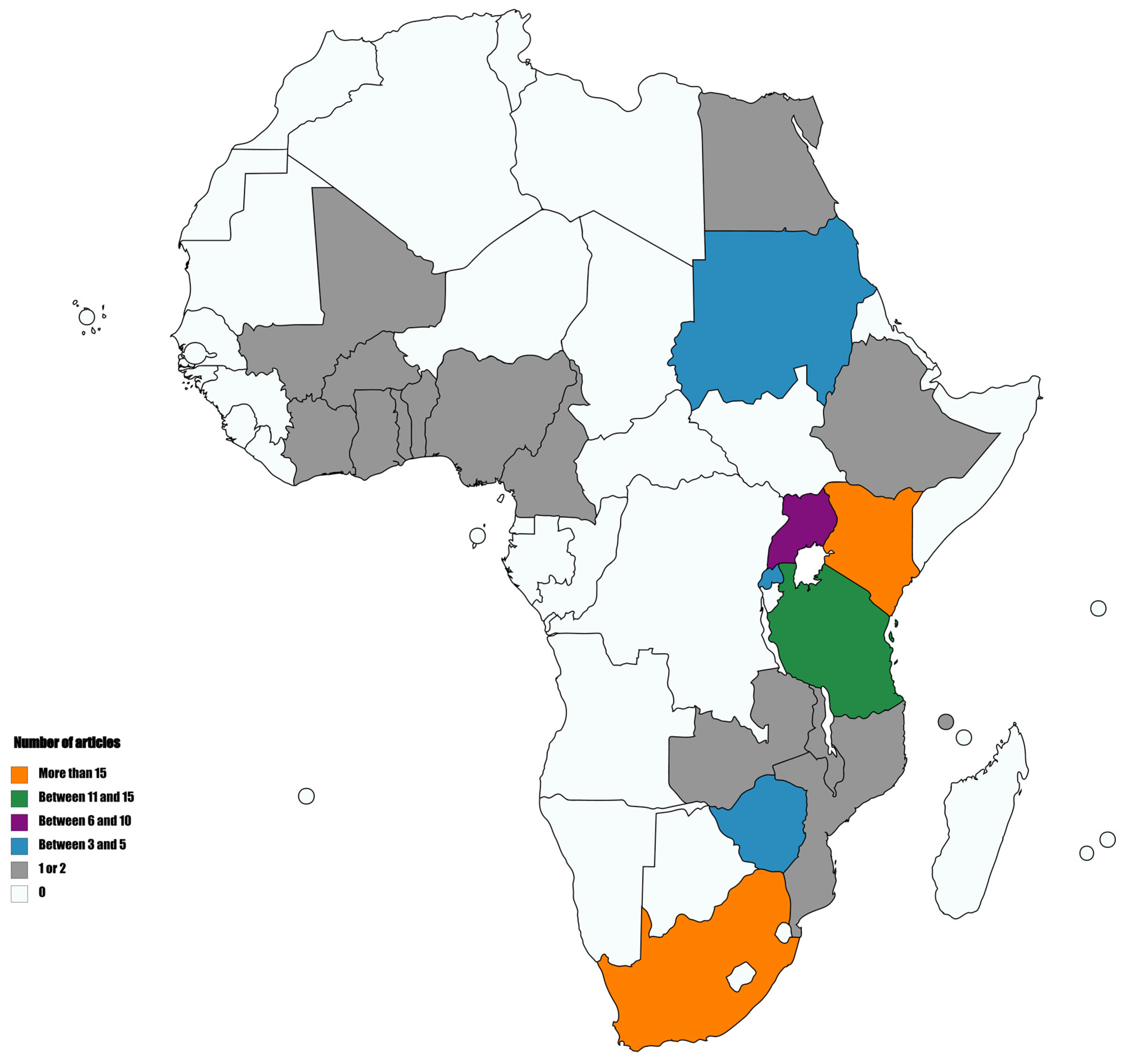
2. Integration of HTS in Plant Virology in Africa
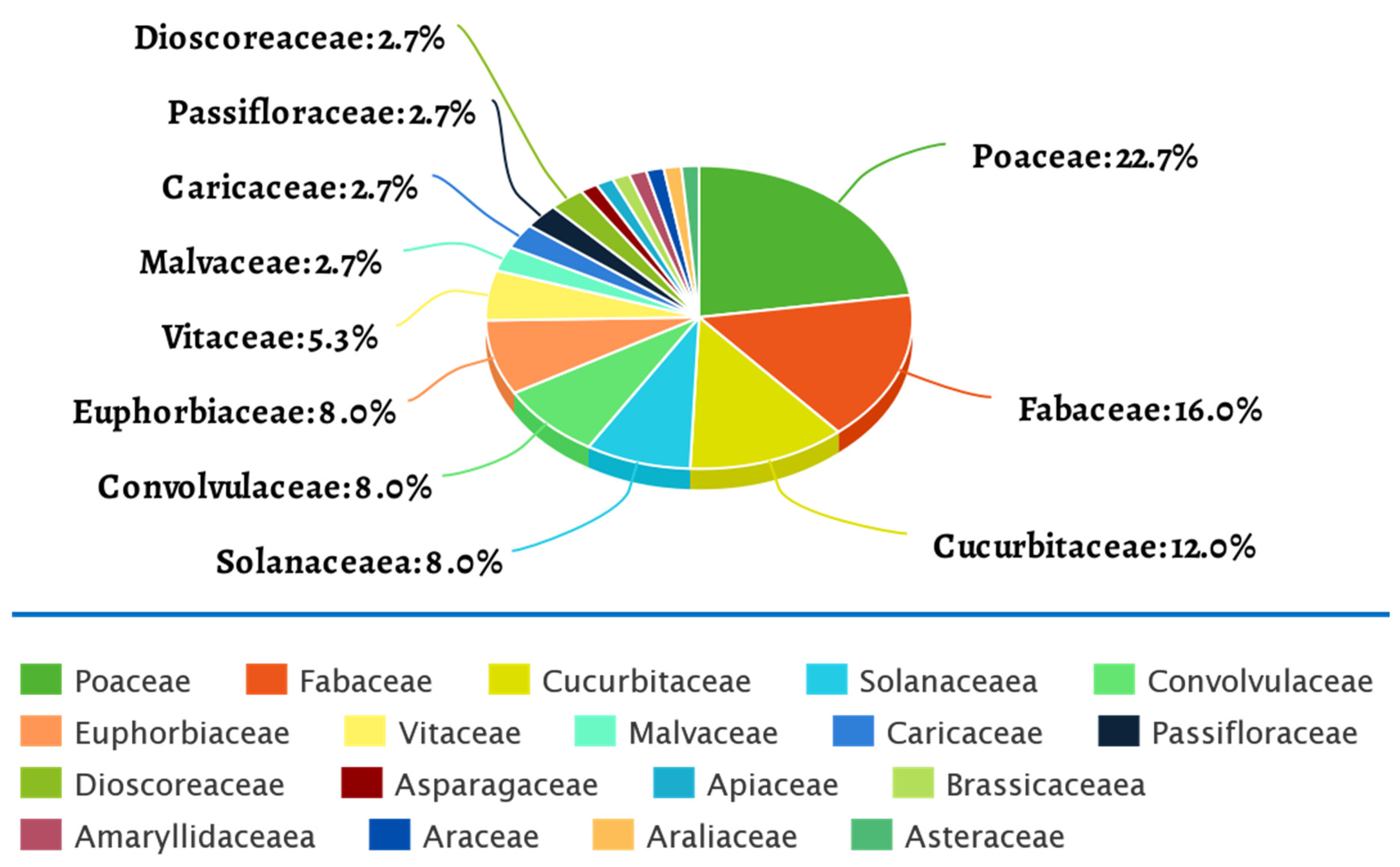
3. Host Plants Subjected to HTS
3.1. Maize
3.2. Common Bean and Cowpea
3.3. Sweet Potato
3.4. Cassava
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Van Der Want, J.P.H.; Dijkstra, J. A history of plant virology. Arch. Virol. 2006, 151, 1467–1498. [Google Scholar] [CrossRef] [PubMed]
- Frizzi, A.; Zhang, Y.; Kao, J.; Hagen, C.; Huang, S. Small RNA Profiles from Virus-Infected Fresh Market Vegetables. J. Agric. Food Chem. 2014, 62, 12067–12074. [Google Scholar] [CrossRef] [PubMed]
- Tolin, S.A.; Fayad, A. Virus diseases of tropical vegetable crops and their management. In Integrated Pest Management of Tropical Vegetable Crops; Muniappan, R., Heinrichs, E.A., Eds.; Springer: Amsterdam, The Netherlands, 2016; pp. 41–76. ISBN 9789402409246. [Google Scholar]
- Makkouk, K.M.; Kumari, S.G.; van Leur, J.A.G.; Jones, R.A.C. Control of plant virus diseases in cool-season grain legume crops. In Advances in Virus Research; Loebenstein, G., Katis, N., Eds.; Academic Press Inc.: Cambridge, MA, USA, 2014; Volume 90, pp. 207–253. [Google Scholar] [CrossRef]
- Hema, M.; Sreenivasulu, P.; Patil, B.L.; Kumar, P.L.; Reddy, D.V.R. Tropical Food Legumes. Virus Diseases of Economic Importance and Their Control. In Advances in Virus Research; Loebenstein, G., Katis, N., Eds.; Academic Press Inc.: Cambridge, MA, USA, 2014; Volume 90, pp. 431–505. [Google Scholar] [CrossRef]
- Rakotondrafara, A.M.; Byamukama, E.; Plumb, R.T. Virus Diseases of Cereals. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 1–12. [Google Scholar] [CrossRef]
- Suzuki, N.; Sasaya, T.; Choi, I.-R. Editorial: Viruses threatening stable production of cereal crops. Front. Microbiol. 2015, 6, 470. [Google Scholar] [CrossRef] [Green Version]
- Umer, M.; Liu, J.; You, H.; Xu, C.; Dong, K.; Luo, N.; Kong, L.; Li, X.; Hong, N.; Wang, G.; et al. Genomic, Morphological and Biological Traits of the Viruses Infecting Major Fruit Trees. Viruses 2019, 11, 515. [Google Scholar] [CrossRef] [Green Version]
- Mitrofanova, I.; Zakubanskiy, A.V.; Mitrofanova, O.V. Viruses infecting main ornamental plants: An overview. Ornam. Hortic. 2018, 24, 95–102. [Google Scholar] [CrossRef]
- Malmstrom, C.M.; Alexander, H.M. Effects of crop viruses on wild plants. Curr. Opin. Virol. 2016, 19, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Elbeaino, T.; Digiaro, M.; Mielke-Ehret, N.; Muehlbach, H.P.; Martelli, G.P. ICTV Virus Taxonomy Profile: Fimoviridae. J. Gen. Virol. 2018, 99, 1478–1479. [Google Scholar] [CrossRef]
- Walker, P.J.; Blasdell, K.R.; Calisher, C.H.; Dietzgen, R.G.; Kondo, H.; Kurath, G.; Longdon, B.; Stone, D.M.; Tesh, R.B.; Tordo, N.; et al. ICTV Virus Taxonomy Profile: Rhabdoviridae. J. Gen. Virol. 2018, 99, 447–448. [Google Scholar] [CrossRef]
- ICTV International Committee on Taxonomy of Viruses (ICTV). Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 23 April 2020).
- Nicaise, V. Crop immunity against viruses: Outcomes and future challenges. Front. Plant Sci. 2014, 5, 660. [Google Scholar] [CrossRef]
- Gergerich, R.C.; Dolja, V.V. Introduction to Plant Viruses, the Invisible Foe. Plant Health Instr. 2006. [Google Scholar] [CrossRef]
- Islam, W.; Noman, A.; Naveed, H.; Alamri, S.A.; Hashem, M.; Huang, Z.; Chen, H.Y.H. Plant-insect vector-virus interactions under environmental change. Sci. Total Environ. 2020, 701, 135044. [Google Scholar] [CrossRef]
- Wylie, S.J.; Adams, M.; Chalam, C.; Kreuze, J.; López-Moya, J.-J.; Ohshima, K.; Praveen, S.; Rabenstein, F.; Stenger, D.; Wang, A.; et al. ICTV Virus Taxonomy Profile: Potyviridae. J. Gen. Virol. 2017, 98, 352–354. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, K.I.; Eskelin, K.; Lõhmus, A.; Mäkinen, K. Molecular and cellular mechanisms underlying potyvirus infection. J. Gen. Virol. 2014, 95, 1415–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, K.; Jhabarmal, J.; Tarafdar, A. Detection of plant viruses for their management: Recent trends trends plant pathogen interaction. In Plant Pathogen Interaction: Recent Trends; Mitra, R., Barman, A., Eds.; Sharma Pubs & Distributors: New Delhi, India, 2016; pp. 71–88. ISBN 9789382310051. [Google Scholar]
- Jeong, J.-J.; Ju, H.-J.; Noh, J. A Review of Detection Methods for the Plant Viruses. Res. Plant Dis. 2014, 20, 173–181. [Google Scholar] [CrossRef]
- Yadav, N.; Khurana, S.M.P. Plant virus detection and diagnosis: Progress and challenges. In Frontier Discoveries and Innovations in Interdisciplinary Microbiology; Shukla, P., Ed.; Springer: New Delhi, India, 2016; pp. 97–132. [Google Scholar] [CrossRef]
- López, M.M.; Llop, P.; Olmos, A.; Marco-Noales, E.; Cambra, M.; Bertolini, E. Are molecular tools solving the challenges posed by detection of plant pathogenic bacteria and viruses? Curr. Issues Mol. Biol. 2009, 11, 13–46. [Google Scholar] [CrossRef] [Green Version]
- Santala, J.; Valkonen, J.P.T. Sensitivity of Small RNA-Based Detection of Plant Viruses. Front. Microbiol. 2018, 9, 939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacroix, C.; Renner, K.; Cole, E.; Seabloom, E.W.; Borer, E.T.; Malmstrom, C.M. Methodological Guidelines for Accurate Detection of Viruses in Wild Plant Species. Appl. Environ. Microbiol. 2016, 82, 1966–1975. [Google Scholar] [CrossRef] [Green Version]
- Kulski, J.K. Next-Generation Sequencing—An Overview of the History, Tools, and “Omic” Applications. In Next Generation Sequencing—Advances, Applications and Challenges; Kulski, J.K., Ed.; InTechOpen: London, UK, 2016; pp. 3–60. [Google Scholar]
- Krishna, B.M.; Khan, M.A.; Khan, S.T. Next-Generation Sequencing (NGS) Platforms: An Exciting Era of Genome Sequence Analysis. In Microbial Genomics in Sustainable Agroecosystems; Tripathi, V., Kumar, P., Tripathi, P., Kishore, A., Kamle, M., Eds.; Springer Nature: Singapore, 2019; Volume 2, pp. 89–109. [Google Scholar]
- McCombie, W.R.; McPherson, J.D.; Mardis, E.R. Next-Generation Sequencing Technologies. Cold Spring Harb. Perspect. Med. 2019, 9, a036798. [Google Scholar] [CrossRef]
- Kchouk, M.; Gibrat, J.-F.; Elloumi, M. Generations of Sequencing Technologies: From First to Next Generation. Biol. Med. 2017, 9, 395. [Google Scholar] [CrossRef] [Green Version]
- Heather, J.M.; Chain, B. The sequence of sequencers: The history of sequencing DNA. Genomics 2016, 107, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.E.; Myers, R.M. Advancements in Next-Generation Sequencing. Annu. Rev. Genom. Hum. Genet. 2016, 17, 95–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barba, M.; Czosnek, H.; Hadidi, A. Historical Perspective, Development and Applications of Next-Generation Sequencing in Plant Virology. Viruses 2014, 6, 106–136. [Google Scholar] [CrossRef]
- Hadidi, A.; Flores, R.; Candresse, T.; Barba, M. Next-Generation Sequencing and Genome Editing in Plant Virology. Front. Microbiol. 2016, 7, 1325. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Baizan-Edge, A.; Macfarlane, S.; Torrance, L. Viral Diagnostics in Plants Using Next Generation Sequencing: Computational Analysis in Practice. Front. Plant Sci. 2017, 8, 1770. [Google Scholar] [CrossRef]
- Massart, S.; Chiumenti, M.; De Jonghe, K.; Glover, R.; Haegeman, A.; Koloniuk, I.; Komínek, P.; Kreuze, J.; Kutnjak, D.; Lotos, L.; et al. Virus Detection by High-Throughput Sequencing of Small RNAs: Large-Scale Performance Testing of Sequence Analysis Strategies. Phytopathology 2019, 109, 488–497. [Google Scholar] [CrossRef] [Green Version]
- Blawid, R.; Silva, J.M.F.; Nagata, T. Discovering and sequencing new plant viral genomes by next-generation sequencing: Description of a practical pipeline. Ann. Appl. Biol. 2017, 170, 301–314. [Google Scholar] [CrossRef]
- Pecman, A.; Kutnjak, D.; Gutiérrez-Aguirre, I.; Adams, I.; Fox, A.; Boonham, N.; Ravnikar, M. Next Generation Sequencing for Detection and Discovery of Plant Viruses and Viroids: Comparison of Two Approaches. Front. Microbiol. 2017, 8, 1998. [Google Scholar] [CrossRef] [Green Version]
- Chalupowicz, L.; Dombrovsky, A.; Gaba, V.; Luria, N.; Reuven, M.; Beerman, A.; Lachman, O.; Dror, O.; Nissan, G.; Manulis-Sasson, S. Diagnosis of plant diseases using the Nanopore sequencing platform. Plant Pathol. 2019, 68, 229–238. [Google Scholar] [CrossRef]
- Villamor, D.E.; Ho, T.; Al Rwahnih, M.; Martin, R.R.; Tzanetakis, I. High Throughput Sequencing for Plant Virus Detection and Discovery. Phytopathology 2019, 109, 716–725. [Google Scholar] [CrossRef]
- Helmy, M.; Awad, M.; Mosa, K.A. Limited resources of genome sequencing in developing countries: Challenges and solutions. Appl. Transl. Genom. 2016, 9, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Mulder, N.; Christoffels, A.; De Oliveira, T.; Gamieldien, J.; Hazelhurst, S.; Joubert, F.; Kumuthini, J.; Pillay, C.S.; Snoep, J.L.; Bishop, Ö.T.; et al. The Development of Computational Biology in South Africa: Successes Achieved and Lessons Learnt. PLoS Comput. Biol. 2016, 12, e1004395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, Ö.T.; Adebiyi, E.F.; Alzohairy, A.M.; Everett, D.; Ghedira, K.; Ghouila, A.; Kumuthini, J.; Mulder, N.; Panji, S.; Patterton, H.-G. Bioinformatics Education—Perspectives and Challenges out of Africa. Brief Bioinform. 2014, 16, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mlotshwa, B.C.; Mwesigwa, S.; Mboowa, G.; Williams, L.; Retshabile, G.; Kekitiinwa, A.; Wayengera, M.; Kyobe, S.; Mardon, G.; Hanchard, N.A.; et al. The collaborative African genomics network training program: A trainee perspective on training the next generation of African scientists. Genet. Med. 2017, 19, 826–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karikari, T. Bioinformatics in Africa: The Rise of Ghana? PLoS Comput. Biol. 2015, 11, e1004308. [Google Scholar] [CrossRef] [Green Version]
- Mulenga, R.M.; Miano, D.W.; Kaimoyo, E.; Akello, J.; Nzuve, F.M.; Al Rwahnih, M.; Chikoti, P.C.; Chiona, M.; Simulundu, E.; Alabi, O.; et al. First Report of Southern Bean Mosaic Virus Infecting Common Bean in Zambia. Plant Dis. 2020, 104, 1880. [Google Scholar] [CrossRef]
- Bester, R.; Maree, H.J. First report of plum bark necrosis stem pitting-associated virus in Japanese plums in South Africa. Plant Dis. 2020, 4. [Google Scholar] [CrossRef]
- Mostert, I.; Visser, M.; Gazendam, I.; Cloete, M.; Burger, J.; Maree, H.J. Complete genome sequence of a novel polerovirus in Ornithogalum thyrsoides from South Africa. Arch. Virol. 2020, 165, 483–486. [Google Scholar] [CrossRef]
- Bester, R.; Burger, J.; Maree, H.J. Genomic characterisation of a newly identified badnavirus infecting ivy (Hedera helix). Arch. Virol. 2020, 1–4. [Google Scholar] [CrossRef]
- Karavina, C.; Ibaba, J.D.; Gubba, A. Full genome sequence of a chrysanthemum-infecting tomato spotted wilt virus isolate from Zimbabwe obtained by next-generation sequencing. Acta Virol. 2020, 64, 88–92. [Google Scholar] [CrossRef]
- Karavina, C.; Ibaba, J.D.; Gubba, A. High-throughput sequencing of virus-infected Cucurbita pepo samples revealed the presence of Zucchini shoestring virus in Zimbabwe. BMC Res. Notes 2020, 13, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avedi, E.K.; Kilalo, C.D.; Olubayo, F.M.; Macharia, I.; Adediji, A.O.; Ateka, E.M.; Machuka, E.M.; Mutuku, J.M. Complete Genome Sequence of a New Chickpea Chlorotic Dwarf Virus Strain Isolated from Tomato in Kenya, Obtained from Illumina Sequencing. Microbiol. Resour. Announc. 2020, 9, e01344-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumo, N.N.; Mamati, G.E.; Ateka, E.M.; Rimberia, F.K.; Asudi, G.O.; Boykin, L.M.; Machuka, E.M.; Njuguna, J.N.; Pelle, R.; Stomeo, F. Metagenomic Analysis of Plant Viruses Associated with Papaya Ringspot Disease in Carica papaya L. in Kenya. Front. Microbiol. 2020, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Read, D.A.; Muoma, J.; Thompson, G.D. Metaviromic analysis reveals coinfection of papaya in western Kenya with a unique strain of Moroccan watermelon mosaic virus and a novel member of the family Alphaflexiviridae. Arch. Virol. 2020, 165, 1231–1234. [Google Scholar] [CrossRef]
- Wokorach, G.; Otim, G.; Njuguna, J.; Edema, H.; NjungE, V.; Machuka, E.M.; Yao, N.; Stomeo, F.; Echodu, R. Genomic analysis of Sweet potato feathery mottle virus from East Africa. Physiol. Mol. Plant Pathol. 2020, 110, 101473. [Google Scholar] [CrossRef]
- Asiimwe, T.; Stewart, L.R.; Willie, K.; Massawe, D.P.; Kamatenesi, J.; Redinbaugh, M.G. Maize lethal necrosis viruses and other maize viruses in Rwanda. Plant Pathol. 2020, 69, 585–597. [Google Scholar] [CrossRef]
- Kiruwa, F.H.; Mutiga, S.K.; Njuguna, J.; Machuka, E.; Senay, S.; Feyissa, T.; Ndakidemi, P.A.; Stomeo, F. Status and Epidemiology of Maize Lethal Necrotic Disease in Northern Tanzania. Pathogens 2019, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Read, D.A.; Featherston, J.; Rees, D.J.G.; Thompson, G.D.; Roberts, R.; Flett, B.C.; Mashingaidze, K.; Pietersen, G.; Kiula, B.; Kullaya, A.; et al. Characterization and detection of maize-associated pteridovirus (MaPV), infecting maize (Zea mays) in the Arusha region of Tanzania. Eur. J. Plant Pathol. 2019, 154, 1165–1170. [Google Scholar] [CrossRef]
- Read, D.A.; Featherston, J.; Rees, D.J.G.; Thompson, G.D.; Roberts, R.; Flett, B.C.; Mashingaidze, K.; Pietersen, G.; Kiula, B.; Kullaya, A.; et al. Diversity and distribution of Maize-associated totivirus strains from Tanzania. Virus Genes 2019, 55, 429–432. [Google Scholar] [CrossRef]
- Read, D.A.; Featherston, J.; Rees, D.J.G.; Thompson, G.D.; Roberts, R.; Flett, B.C.; Mashingaidze, K.; Pietersen, G.; Kiula, B.; Kullaya, A.; et al. Molecular characterization of Morogoro maize-associated virus, a nucleorhabdovirus detected in maize (Zea mays) in Tanzania. Arch. Virol. 2019, 164, 1711–1715. [Google Scholar] [CrossRef]
- Read, D.A.; Featherstone, J.; Rees, D.J.G.; Thompson, G.; Roberts, R.; Flett, B.C.; Mashingaidze, K.; Berger, D.K.; Welgemoed, T.; Pietersen, G.; et al. First report of maize yellow mosaic virus (MaYMV) on maize (Zea mays) in Tanzania. J. Plant Pathol. 2019, 101, 203. [Google Scholar] [CrossRef]
- Moodley, V.; Gubba, A.; Mafongoya, P.L. Prevalence, epidemiology and molecular studies of Tomato chlorosis virus (ToCV) in South Africa. PLoS ONE 2019, 14, e0220298. [Google Scholar] [CrossRef] [Green Version]
- Mulabisana, M.; Cloete, M.; Laurie, S.; Mphela, W.; Maserumule, M.; Nhlapo, T.; Cochrane, N.; Oelofse, D.; Rey, M. Yield evaluation of multiple and co-infections of begomoviruses and potyviruses on sweet potato varieties under field conditions and confirmation of multiple infection by NGS. Crop Prot. 2019, 119, 102–112. [Google Scholar] [CrossRef]
- Richet, C.; Kraberger, S.; Filloux, D.; Bernardo, P.; Harkins, G.W.; Martin, D.P.; Roumagnac, P.; Varsani, A. Novel circular DNA viruses associated with Apiaceae and Poaceae from South Africa and New Zealand. Arch. Virol. 2019, 164, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Karavina, C.; Jacques, D.I.; Augustine, G. Characterization of three full Iris yellow spot virus genes of a garlic-infecting isolate from Zimbabwe using next-generation sequencing technology. Afr. J. Biotechnol. 2019, 18, 928–934. [Google Scholar] [CrossRef] [Green Version]
- Amisse, J.; Ndunguru, J.; Tairo, F.; Ateka, E.; Boykin, L.M.; Kehoe, M.A.; Cossa, N.; Rey, C.; Sseruwagi, P. Analyses of seven new whole genome sequences of cassava brown streak viruses in Mozambique reveals two distinct clades: Evidence for new species. Plant Pathol. 2019, 68, 1007–1018. [Google Scholar] [CrossRef] [Green Version]
- Scussel, S.; Candresse, T.; Marais, A.; Claverie, S.; Hoareau, M.; Azali, H.A.; Verdin, E.; Tepfer, M.; Filloux, D.; Fernandez, E.; et al. High-throughput sequencing of complete genomes of ipomoviruses associated with an epidemic of cassava brown streak disease in the Comoros Archipelago. Arch. Virol. 2019, 164, 2193–2196. [Google Scholar] [CrossRef]
- Desbiez, C.; Caciagli, P.; Wipf-Scheibel, C.; Millot, P.; Ruiz, L.; Marian, D.; Dafalla, G.; Lecoq, H. Evidence for long-term prevalence of cucumber vein yellowing virus in Sudan and genetic variation of the virus in Sudan and the Mediterranean Basin. Plant Pathol. 2019, 68, 1268–1275. [Google Scholar] [CrossRef] [Green Version]
- Kidanemariam, D.B.; Sukal, A.C.; Abraham, A.D.; Njuguna, J.N.; Stomeo, F.; Dale, J.; Harding, R.; James, A.P. Molecular characterisation of a putative new polerovirus infecting pumpkin (Cucurbita pepo) in Kenya. Arch. Virol. 2019, 164, 1717–1721. [Google Scholar] [CrossRef]
- Mukoye, B.; Ndonga, M.F.O.; Were, H.K. Incidence of Groundnut Rosette Disease (GRD) and Genetic Diversity of Groundnut Rosette Assistor Virus (GRAV) in Western Kenya. Int. J. Genet. Genom. 2019, 7, 98. [Google Scholar] [CrossRef]
- Collins, M.B.; Karakacha, W.H.; Benard, M.; Milicent, N. First Full Length Genome Sequence of Bean Common Mosaic Necrosis Virus (BCMNV) Isolated from Common Bean in Western Kenya. Int. J. Genet. Genom. 2019, 7, 132. [Google Scholar] [CrossRef]
- Wainaina, J.M.; Ateka, E.; Makori, T.; Kehoe, M.A.; Boykin, L.M. A metagenomic study of DNA viruses from samples of local varieties of common bean in Kenya. PeerJ 2019, 7, e6465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wainaina, J.M.; Kubatko, L.; Harvey, J.; Ateka, E.; Makori, T.; Karanja, D.; Boykin, L.M.; Kehoe, M.A. Evolutionary insights of Bean common mosaic necrosis virus and Cowpea aphid-borne mosaic virus. PeerJ 2019, 7, e6297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munguti, F.; Maina, S.; Nyaboga, E.N.; Kilalo, D.; Kimani, E.; Macharia, M.; Holton, T. Transcriptome Sequencing Reveals a Complete Genome Sequence of Cowpea Aphid-Borne Mosaic Virus from Passion Fruit in Kenya. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidanemariam, D.B.; Sukal, A.C.; Abraham, A.D.; Njuguna, J.N.; Mware, B.O.; Stomeo, F.; Dale, J.L.; James, A.P.; Harding, R. Characterisation of a subgroup IB isolate of Cucumber mosaic virus from Xanthosoma sp. in sub-Saharan Africa. Australas. Plant Pathol. 2019, 48, 457–460. [Google Scholar] [CrossRef]
- Mbeyagala, E.K.; Maina, S.; Macharia, M.W.; Mukasa, S.B.; Holton, T. Illumina Sequencing Reveals the First Near-Complete Genome Sequence of Ugandan Passiflora Virus. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Bömer, M.; Rathnayake, A.I.; Visendi, P.; Sewe, S.O.; Sicat, J.P.A.; Silva, G.; Kumar, P.L.; Seal, S. Tissue culture and next-generation sequencing: A combined approach for detecting yam (Dioscorea spp.) viruses. Physiol. Mol. Plant Pathol. 2019, 105, 54–66. [Google Scholar] [CrossRef]
- Silva, G.; Bömer, M.; Rathnayake, A.I.; Sewe, S.O.; Visendi, P.; Oyekanmi, J.; Quain, M.D.; Akomeah, B.; Kumar, P.L.; Seal, S. Molecular Characterization of a New Virus Species Identified in Yam (Dioscorea spp.) by High-Throughput Sequencing. Plants 2019, 8, 167. [Google Scholar] [CrossRef] [Green Version]
- Karavina, C.; Ibaba, J.D.; Gubba, A. Genome Sequence of a Tomato-Infecting Tomato Mosaic Virus Isolate from Zimbabwe. Genome Announc. 2018, 6, e01457-17. [Google Scholar] [CrossRef] [Green Version]
- Mulabisana, M.; Cloete, M.; Mabasa, K.; Laurie, S.; Oelofse, D.; Esterhuizen, L.; Rey, M. Surveys in the Gauteng, Limpopo and Mpumalanga provinces of South Africa reveal novel isolates of sweet potato viruses. S. Afr. J. Bot. 2018, 114, 280–294. [Google Scholar] [CrossRef]
- Nhlapo, T.; Rees, D.; Odeny, D.; Mulabisana, J.; Rey, M. Viral metagenomics reveals sweet potato virus diversity in the Eastern and Western Cape provinces of South Africa. S. Afr. J. Bot. 2018, 117, 256–267. [Google Scholar] [CrossRef]
- Nhlapo, T.F.; Mulabisana, J.M.; Odeny, D.A.; Rey, M.E.C.; Rees, D.J.G. First Report of Sweet potato badnavirus A and Sweet potato badnavirus B in South Africa. Plant Dis. 2018, 102, 1865. [Google Scholar] [CrossRef] [PubMed]
- Wainaina, J.M.; Ateka, E.; Makori, T.; Kehoe, M.A.; Boykin, L.M. Phylogenomic relationship and evolutionary insights of sweet potato viruses from the western highlands of Kenya. PeerJ 2018, 6, e5254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wainaina, J.M.; Harvey, J.; Ateka, E.; Makori, T.; Karanja, D.; Kehoe, M.A.; Boykin, L.M. Genomic characterisation and evolutionary relationships of groundnut rosette virus from the western highlands of Kenya. Trop. Plant Pathol. 2018, 43, 583–585. [Google Scholar] [CrossRef]
- Mutuku, J.M.; Wamonje, F.O.; Mukeshimana, G.; Njuguna, J.; Wamalwa, M.; Choi, S.-K.; Tungadi, T.; Djikeng, A.; Kelly, K.; Entfellner, J.-B.D.; et al. Metagenomic Analysis of Plant Virus Occurrence in Common Bean (Phaseolus vulgaris) in Central Kenya. Front. Microbiol. 2018, 9, 2939. [Google Scholar] [CrossRef]
- Mbeyagala, E.K.; Tukamuhabwa, P.; Bisikwa, J.; Holton, T.; Mukasa, S.B. Next-Generation Sequencing Reveals the First Complete Genome Sequence of Cowpea aphid-borne mosaic virus from Uganda. Genome Announc. 2018, 6, e01491-17. [Google Scholar] [CrossRef] [Green Version]
- Mwaipopo, B.; Njau, P.J.R.; Mark, D.; Mbanzibwa, D.; Nchimbi-Msolla, S. Comprehensive Surveys of Bean common mosaic virus and Bean common mosaic necrosis virus and Molecular Evidence for Occurrence of Other Phaseolus vulgaris Viruses in Tanzania. Plant Dis. 2018, 102, 2361–2370. [Google Scholar] [CrossRef] [Green Version]
- Massawe, D.P.; Stewart, L.R.; Kamatenesi, J.; Asiimwe, T.; Redinbaugh, M.G. Complete sequence and diversity of a maize-associated Polerovirus in East Africa. Virus Genes 2018, 54, 432–437. [Google Scholar] [CrossRef]
- Wamaitha, M.J.; Nigam, D.; Maina, S.; Stomeo, F.; Wangai, A.; Njuguna, J.; Holton, T.A.; Wanjala, B.W.; Wamalwa, M.; Lucas, T.; et al. Metagenomic analysis of viruses associated with maize lethal necrosis in Kenya. Virol. J. 2018, 15, 90. [Google Scholar] [CrossRef]
- Muller, E.; Ravel, S.; Agret, C.; Abrokwah, F.; Dzahini-Obiatey, H.; Galyuon, I.; Kouakou, K.; Jeyaseelan, E.; Allainguillaume, J.; Wetten, A. Next generation sequencing elucidates cacao badnavirus diversity and reveals the existence of more than ten viral species. Virus Res. 2018, 244, 235–251. [Google Scholar] [CrossRef] [Green Version]
- Afouda, L.; Kone, D.; Zinsou, V.; Dossou, L.; Kenyon, L.; Winter, S.; Knierim, D. Virus surveys of Capsicum spp. in the Republic of Benin reveal the prevalence of pepper vein yellows virus and the identification of a previously uncharacterised polerovirus species. Arch. Virol. 2017, 162, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Mbewe, W.; Winter, S.; Mukasa, S.; Tairo, F.; Sseruwagi, P.; Ndunguru, J.; Duffy, S. Deep Sequencing Reveals a Divergent Ugandan cassava brown streak virus Isolate from Malawi. Genome Announc. 2017, 5, e00818-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordenstedt, N.; Marcenaro, D.; Chilagane, D.; Mwaipopo, B.; Rajamäki, M.-L.; Nchimbi-Msolla, S.; Njau, P.J.R.; Mbanzibwa, D.; Valkonen, J.P.T. Pathogenic seedborne viruses are rare but Phaseolus vulgaris endornaviruses are common in bean varieties grown in Nicaragua and Tanzania. PLoS ONE 2017, 12, e0178242. [Google Scholar] [CrossRef]
- Desbiez, C.; Wipf-Scheibel, C.; Millot, P.; Verdin, E.; Dafalla, G.; Lecoq, H. New species in the papaya ringspot virus cluster: Insights into the evolution of the PRSV lineage. Virus Res. 2017, 241, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibaba, J.D.; Laing, M.D.; Gubba, A. Pepo aphid-borne yellows virus: A new species in the genus Polerovirus. Virus Genes 2017, 53, 134–136. [Google Scholar] [CrossRef] [PubMed]
- Ximba, S.; Ibaba, J.D.; Gubba, A. Potato virus Y strains infecting potatoes in the Msinga district in the province of KwaZulu-Natal, South Africa. Crop Prot. 2017, 96, 188–194. [Google Scholar] [CrossRef]
- New, S.-A.; Van Heerden, S.W.; Pietersen, G.; Esterhuizen, L.L. First report of a Turnip Yellows virus in association with the Brassica stunting disorder in South Africa. Plant Dis. 2016, 100, 2341. [Google Scholar] [CrossRef]
- Ibaba, J.D.; Laing, M.D.; Gubba, A. Zucchini shoestring virus: A distinct potyvirus in the papaya ringspot virus cluster. Arch. Virol. 2016, 161, 2321–2323. [Google Scholar] [CrossRef]
- Ibaba, J.D.; Laing, M.D.; Gubba, A. Genome sequence analysis of two South African isolates of Moroccan watermelon mosaic virus infecting cucurbits. Virus Genes 2016, 52, 896–899. [Google Scholar] [CrossRef]
- Lecoq, H.; Verdin, E.; Tepfer, M.; Wipf-Scheibel, C.; Millot, P.; Dafalla, G.; Desbiez, C. Characterization and occurrence of squash chlorotic leaf spot virus, a tentative new torradovirus infecting cucurbits in Sudan. Arch. Virol. 2016, 161, 1651–1655. [Google Scholar] [CrossRef]
- Desbiez, C.; Verdin, E.; Tepfer, M.; Wipf-Scheibel, C.; Millot, P.; Dafalla, G.; Lecoq, H. Characterization of a new cucurbit-infecting ipomovirus from Sudan. Arch. Virol. 2016, 161, 2913–2915. [Google Scholar] [CrossRef] [PubMed]
- Leke, W.; Mignouna, D.; Brown, J.; Fondong, V. First report of Soybean chlorotic blotch virus and West African Asystasia virus 1 infecting cassava and a wild cassava relative in Cameroon and Togo. New Dis. Rep. 2016, 33, 24. [Google Scholar] [CrossRef] [Green Version]
- Leke, W.N.; Khatabi, B.; Fondong, V.N.; Brown, J.K. Complete genome sequence of a new bipartite begomovirus infecting fluted pumpkin (Telfairia occidentalis) plants in Cameroon. Arch. Virol. 2016, 161, 2347–2350. [Google Scholar] [CrossRef] [PubMed]
- Leke, W.N.; Khatabi, B.; Mignouna, D.B.; Brown, J.K.; Fondong, V.N. Complete genome sequence of a new bipartite begomovirus infecting cotton in the Republic of Benin in West Africa. Arch. Virol. 2016, 161, 2329–2333. [Google Scholar] [CrossRef] [PubMed]
- Palanga, E.; Filloux, D.; Martin, D.P.; Fernández, E.M.; Gargani, D.; Ferdinand, R.; Zabré, J.; Bouda, Z.; Neya, J.B.; Sawadogo, M.; et al. Metagenomic-Based Screening and Molecular Characterization of Cowpea-Infecting Viruses in Burkina Faso. PLoS ONE 2016, 11, e0165188. [Google Scholar] [CrossRef] [Green Version]
- Maree, H.J.; Pirie, M.D.; Oosthuizen, K.; Bester, R.; Jasper, D.; Rees, G.; Burger, J.T. Phylogenomic Analysis Reveals Deep Divergence and Recombination in an Economically Important Grapevine Virus. PLoS ONE 2015, 10, e0126819. [Google Scholar] [CrossRef]
- Molenaar, N.; Burger, J.T.; Maree, H.J. Detection of a divergent variant of grapevine virus F by next-generation sequencing. Arch. Virol. 2015, 160, 2125–2127. [Google Scholar] [CrossRef]
- Ndunguru, J.; Sseruwagi, P.; Tairo, F.; Stomeo, F.; Maina, S.; Djinkeng, A.; Kehoe, M.; Boykin, L.M. Analyses of Twelve New Whole Genome Sequences of Cassava Brown Streak Viruses and Ugandan Cassava Brown Streak Viruses from East Africa: Diversity, Supercomputing and Evidence for Further Speciation. PLoS ONE 2015, 10, e0139321. [Google Scholar] [CrossRef]
- Mahuku, G.; Wangai, A.; Sadessa, K.; Teklewold, A.; Wegary, D.; Ayalneh, D.; Adams, I.; Smith, J.; Bottomley, E.; Bryce, S.; et al. First Report of Maize chlorotic mottle virus and Maize Lethal Necrosis on Maize in Ethiopia. Plant Dis. 2015, 99, 1870. [Google Scholar] [CrossRef]
- Adams, I.P.; Harju, V.; Hodges, T.; Hany, U.; Skelton, A.; Rai, S.; Deka, M.; Smith, J.; Fox, A.; Uzayisenga, B.; et al. First report of maize lethal necrosis disease in Rwanda. New Dis. Rep. 2014, 29, 22. [Google Scholar] [CrossRef] [Green Version]
- Mbanzibwa, D.; Tugume, A.; Chiunga, E.; Mark, D.; Tairo, F. Small RNA deep sequencing-based detection and further evidence of DNA viruses infecting sweetpotato plants in Tanzania. Ann. Appl. Biol. 2014, 165, 329–339. [Google Scholar] [CrossRef]
- Candresse, T.; Filloux, D.; Muhire, B.; Julian, C.; Galzi, S.; Fort, G.; Bernardo, P.; Daugrois, J.-H.; Fernandez, E.; Martin, D.P.; et al. Appearances Can Be Deceptive: Revealing a Hidden Viral Infection with Deep Sequencing in a Plant Quarantine Context. PLoS ONE 2014, 9, e102945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, I.P.; Miano, D.W.; Kinyua, Z.M.; Wangai, A.; Kimani, E.; Phiri, N.; Reeder, R.; Harju, V.; Glover, R.; Hany, U.; et al. Use of next-generation sequencing for the identification and characterization ofMaize chlorotic mottle virusandSugarcane mosaic viruscausing maize lethal necrosis in Kenya. Plant Pathol. 2013, 62, 741–749. [Google Scholar] [CrossRef]
- Espach, Y.; Maree, H.J.; Burger, J.T. Complete Genome of a Novel Endornavirus Assembled from Next-Generation Sequence Data. J. Virol. 2012, 86, 13142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monger, W.A.; Alicai, T.; Ndunguru, J.; Kinyua, Z.M.; Potts, M.; Reeder, R.H.; Miano, D.W.; Adams, I.P.; Boonham, N.; Glover, R.; et al. The complete genome sequence of the Tanzanian strain of Cassava brown streak virus and comparison with the Ugandan strain sequence. Arch. Virol. 2010, 155, 429–433. [Google Scholar] [CrossRef]
- Coetzee, B.; Freeborough, M.-J.; Maree, H.J.; Celton, J.-M.; Rees, D.J.G.; Burger, J.T. Deep sequencing analysis of viruses infecting grapevines: Virome of a vineyard. Virology 2010, 400, 157–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badu-Apraku, B.; Fakorede, M.A.B.; Badu-Apraku, B.; Fakorede, M.A.B. Maize in Sub-Saharan Africa: Importance and Production Constraints. In Advances in Genetic Enhancement of Early and Extra-Early Maize for Sub-Saharan Africa; Badu-Apraku, B., Fakorede, M.A.B., Eds.; Springer International Publishing: Basel, Switzerland, 2017; pp. 3–10. [Google Scholar]
- VIB. VIB’s Facts Series. Available online: http://www.vib.be/en/about-vib/Documents/VIB_MaizeInAfrica_EN_2017.pdf (accessed on 21 May 2020).
- Stewart, L.R.; Willie, K.; Wijeratne, S.; Niblett, C.L.; Asiimwe, T.; Redinbaugh, M.G.; Massawe, D.P.; Kiggundu, A. Johnsongrass mosaic virus Contributes to Maize Lethal Necrosis in East Africa. Plant Dis. 2017, 101, 1455–1462. [Google Scholar] [CrossRef] [Green Version]
- Owade, J.O.; Abong’, G.; Okoth, M.; Mwang’Ombe, A.W. A review of the contribution of cowpea leaves to food and nutrition security in East Africa. Food Sci. Nutr. 2020, 8, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Mukankusi, C.M.; Raatz, B.; Nkalubo, S.; Berhanu, F.; Binagwa, P.; Kilango, M.; Williams, M.; Enid, K.; Chirwa, R.; Beebe, S. Genomics, genetics and breeding of common bean in Africa: A review of tropical legume project. Plant Breed. 2019, 138, 401–414. [Google Scholar] [CrossRef]
- Zheng, Y.; Gao, S.; Padmanabhan, C.; Li, R.; Galvez, M.; Gutierrez, D.; Fuentes, S.; Ling, K.-S.; Kreuze, J.F.; Fei, Z. VirusDetect: An automated pipeline for efficient virus discovery using deep sequencing of small RNAs. Virology 2017, 500, 130–138. [Google Scholar] [CrossRef]
- Menzel, P.; Ng, K.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, T.-H.; Li, P.-G. Sweet potato: Origin and production. In Sweet Potato: Chemistry, Processing and Nutrition; Mu, T.-H., Singh, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 5–25. ISBN 9780128136379. [Google Scholar]
- Kreuze, J.F.; Perez, A.; Gargurevich, M.G.; Cuellar, W.J. Badnaviruses of Sweet Potato: Symptomless Coinhabitants on a Global Scale. Front. Plant Sci. 2020, 11, 313. [Google Scholar] [CrossRef] [PubMed]
- Spencer, D.S.C.; Ezedinma, C. Cassava cultivation in sub-Saharan Africa. In Achieving Sustainable Cultivation of Cassava; Hershey, C.H., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2017; Volume 1, pp. 123–148. [Google Scholar]
- Tumwegamire, S.; Kanju, E.; Legg, J.; Shirima, R.; Kombo, S.; Mkamilo, G.; Mtunda, K.; Sichalwe, K.; Kulembeka, H.; Ndyetabura, I.; et al. Exchanging and managing in-vitro elite germplasm to combat Cassava Brown Streak Disease (CBSD) and Cassava Mosaic Disease (CMD) in Eastern and Southern Africa. Food Secur. 2018, 10, 351–368. [Google Scholar] [CrossRef] [Green Version]
- Abass, A.B.; Awoyale, W.; Alenkhe, B.; Malu, N.; Asiru, B.W.; Manyong, V.; Sanginga, N. Can food technology innovation change the status of a food security crop? A review of cassava transformation into “bread” in Africa. Food Rev. Int. 2018, 34, 87–102. [Google Scholar] [CrossRef]
- FAO. Save and Grow Cassava: A Guide to Sustainable Production and Identification; FAO: Rome, Italy, 2013. [Google Scholar]
- Food and Agriculture Organization of the United Nations FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 23 September 2020).
- Huson, D.H.; Auch, A.F.; Qi, J.; Schuster, S.C. MEGAN analysis of metagenomic data. Genome Res. 2007, 17, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomlinson, K.R.; Bailey, A.M.; Alicai, T.; Seal, S.; Foster, G.D. Cassava brown streak disease: Historical timeline, current knowledge and future prospects. Mol. Plant Pathol. 2018, 19, 1282–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddell, S.G.; Walker, P.J.; Lefkowitz, E.J.; Mushegian, A.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; Knowles, N.J.; Kropinski, A.M.; et al. Binomial nomenclature for virus species: A consultation. Arch. Virol. 2020, 165, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Fox, A. Reconsidering causal association in plant virology. Plant Pathol. 2020, ppa.13199. [Google Scholar] [CrossRef]
- Van Regenmortel, M.H.V. Solving the species problem in viral taxonomy: Recommendations on non-Latinized binomial species names and on abandoning attempts to assign metagenomic viral sequences to species taxa. Arch. Virol. 2019, 164, 2223–2229. [Google Scholar] [CrossRef]
- Jones, R.A.C.; Kehoe, M.A. A proposal to rationalize within-species plant virus nomenclature: Benefits and implications of inaction. Arch. Virol. 2016, 161, 2051–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P.; Valkonen, J.P.T.; Gray, S.M.; Boonham, N.; Jones, R.A.C.; Kerlan, C.; Schubert, J. Discussion paper: The naming of Potato virus Y strains infecting potato. Arch. Virol. 2008, 153, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, J.; Khan, J. The current naming of plant viruses: A critical appraisal. Arch. Virol. 2002, 147, 2251–2253. [Google Scholar] [CrossRef] [PubMed]
- Guy, P.L.; Gerard, P.J. First record of White clover cryptic virus-2 in New Zealand. Australas. Plant Dis. Notes 2018, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Subbarao, B.L.; Kumari, R.; Hallan, V. Molecular characterization of a novel cryptic virus infecting pigeonpea plants. PLoS ONE 2017, 12, e0181829. [Google Scholar] [CrossRef] [PubMed]
- Nerva, L.; Vigani, G.; Di Silvestre, D.; Ciuffo, M.; Forgia, M.; Chitarra, W.; Turina, M. Biological and Molecular Characterization of Chenopodium quinoa Mitovirus 1 Reveals a Distinct Small RNA Response Compared to Those of Cytoplasmic RNA Viruses. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuhara, T.; Tabara, M.; Koiwa, H.; Takahashi, H. Effect of asymptomatic infection with southern tomato virus on tomato plants. Arch. Virol. 2020, 165, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Adams, I.P.; Fox, A.; Boonham, N.; Massart, S.; De Jonghe, K. The impact of high throughput sequencing on plant health diagnostics. Eur. J. Plant Pathol. 2018, 152, 909–919. [Google Scholar] [CrossRef]
- Massart, S.; Candresse, T.; Gil, J.; Lacomme, C.; Predajna, L.; Ravnikar, M.; Reynard, J.-S.; Rumbou, A.; Saldarelli, P.; Škorić, D.; et al. A Framework for the Evaluation of Biosecurity, Commercial, Regulatory, and Scientific Impacts of Plant Viruses and Viroids Identified by NGS Technologies. Front. Microbiol. 2017, 8, 45. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Virus Name | Abbreviation | Genus | Family |
|---|---|---|---|
| African cassava mosaic virus | ACMV | Begomovirus | Geminiviridae |
| African eggplant yellowing virus | AeYV | Polerovirus | Luteoviridae |
| Arctopus echinatus-associated virus | AeaV | N/A | N/A |
| Barley virus G | BVG | Polerovirus | Luteoviridae |
| Barley yellow dwarf virus | BYDV | Luteovirus | Luteoviridae |
| Bean common mosaic virus | BCMV | Potyvirus | Potyviridae |
| Bean common mosaic virus strain blackeye lackeye cowpea mosaic | BCMV-BlCM | Potyvirus | Potyviridae |
| Bean common mosaic necrosis virus | BCMNV | Potyvirus | Potyviridae |
| Bean yellow disorder virus | BnYDV | Crinivirus | Closteroviridae |
| Cacao swollen shoot virus | CSSV | Badnavirus | Caulimoviridae |
| Cacao swollen shoot CD virus | CSSCDV | Badnavirus | Caulimoviridae |
| Cacao swollen shoot CE virus | CSSCEV | Badnavirus | Caulimoviridae |
| Cacao swollen shoot Ghana M virus | CSSGMV | Badnavirus | Caulimoviridae |
| Cacao swollen shoot Ghana N virus | CSSGNV | Badnavirus | Caulimoviridae |
| Cacao swollen shoot Ghana Q virus | CSSGQV | Badnavirus | Caulimoviridae |
| Cacao swollen shoot Ghana R virus | CSSGRV | Badnavirus | Caulimoviridae |
| Cacao swollen shoot Togo A virus | CSSTAV | Badnavirus | Caulimoviridae |
| Cassava brown streak virus | CBSV | Ipomovirus | Potyviridae |
| Chickpea chlorotic dwarf virus tomato strain | CpCDV-T | Mastrevirus | Geminiviridae |
| Coccinia mottle virus | CocMoV | Ipomovirus | Potyviridae |
| Cotton yellow mosaic virus | CYMV | Begomovirus | Geminiviridae |
| Cowpea aphid-borne mosaic virus | CABMV | Potyvirus | Potyviridae |
| Cowpea associated mycotymovirid 1 | CPaMV-1 | N/A | Tymoviridae |
| Cowpea mild mottle virus | CPMMV | Carlavirus | Betaflexiviridae |
| Cowpea mottle virus | CPMoV | Gammacarmovirus | Tombusviridae |
| Cowpea polerovirus 1 | CPPV-1 | Polerovirus | Luteoviridae |
| Cowpea polerovirus 2 | CPPV-2 | Polerovirus | Luteoviridae |
| Cowpea tombusvirid 1 | CPTV-1 | N/A | Tombusviridae |
| Cowpea tombusvirid 2 | CPTV-2 | N/A | Tombusviridae |
| Cucumber mosaic virus | CMV | Cucumovirus | Bromoviridae |
| Cucumber vein yellowing virus | CVYV | Ipomovirus | Potyviridae |
| Dioscorea bacilliform RT virus 3 | DBRTV3 | Badnavirus | Caulimoviridae |
| Dioscorea mosaic-associated virus | DMaV | Sadwavirus | Secoviridae |
| Grapevine endophyte alphaendornavirus | GEEV | Alphaendornavirus | Endornaviridae |
| Grapevine leafroll-associated virus 3 | GLRaV-3 | Ampelovirus | Closteroviridae |
| Grapevine rupestris stem pitting-associated virus | GRSPaV | Foveavirus | Betaflexiviridae |
| Grapevine virus A | GVA | Vitivirus | Betaflexiviridae |
| Grapevine virus E | GVE | Vitivirus | Betaflexiviridae |
| Grapevine virus F | GVF | Vitivirus | Betaflexiviridae |
| Groundnut rosette virus | GRV | Umbravirus | Tombusviridae |
| Groundnut rosette assistor virus | GRAV | N/A | Luteoviridae |
| Iris yellow spot virus | IYSV | Orthotospovirus | Tospoviridae |
| Ivy ringspot-associated virus | IRSaV | Badnavirus | Caulimoviridae |
| Johnsongrass mosaic virus | JGMV | Potyvirus | Potyviridae |
| Lolium perenne-associated virus | LpaV | N/A | N/A |
| Maize-associated betaflexivirus | MaBV | Chordovirus | Betaflexaviridae |
| Maize-associated pteridovirus | MaPV | Pteridovirus | Mayoviridae |
| Maize-associated totivirus | MATV | Totivirus | Totiviridae |
| Maize-associated totivirus-1-Tanz | MATV-1-Tanz | Totivirus | Totiviridae |
| Maize-associated totivirus-4-Tanz | MATV-4-Tanz | Totivirus | Totiviridae |
| Maize dwarf mosaic virus | MDMV | Potyvirus | Potyviridae |
| Maize chlorotic mottle virus | MCMV | Machlomovirus | Tombusviridae |
| Maize pteridovirus 1 | MPtV-1 | Pteridovirus | Mayoviridae |
| Maize streak virus | MSV | Mastrevirus | Geminiviridae |
| Maize yellow dwarf virus RMV2 | MYDV-RMV2 | Polerovirus | Luteoviridae |
| Maize yellow mosaic virus | MaYMV | Polerovirus | Luteoviridae |
| Moroccan watermelon mosaic virus | MWMV | Potyvirus | Potyviridae |
| Morogoro maize-associated virus | MMaV | Alphanucleorhabdovirus | Rhabdoviridae |
| Ornithogalum mosaic virus | OrMV | Potyvirus | Potyviridae |
| Ornithogalum virus 3 | OV-3 | Potyvirus | Potyviridae |
| Ornithogalum virus 5 | OV-5 | Polerovirus | Luteoviridae |
| Panicum ecklonii-associated virus | PeaV | N/A | N/A |
| Papaya mild mottle-associated virus | PaMMV | Carlavirus | Betaflexiviridae |
| Papaya mottle-associated virus | PaMV | Carlavirus | Betaflexiviridae |
| Papaya virus A | PaVA | Allexivirus | Alphaflexiviridae |
| Passiflora virus | PV | Potyvirus | Potyviridae |
| Peanut mottle virus | PeMoV | Potyvirus | Potyviridae |
| Pelargonium vein-banding virus | PVBV | Badnavirus | Caulimoviridae |
| Pepo aphid-borne yellows virus | PABYV | Polerovirus | Luteoviridae |
| Phaseolus vulgaris alphaendornavirus 1 | PvEV-1 | alphaendornavirus | Endornaviridae |
| Phaseolus vulgaris alphaendornavirus 2 | PvEV-2 | alphaendornavirus | Endornaviridae |
| Potato Virus Y | PVY | Potyvirus | Potyviridae |
| Plum bark necrosis stem-pitting-associated virus | PBNSPaV | Ampelovirus | Closteroviridae |
| Pumpkin polerovirus | PuPV | Polerovirus | Luteoviridae |
| Scallion mosaic virus | ScaMV | Potyvirus | Potyviridae |
| Sorghum mosaic virus | SrMV | Potyvirus | Potyviridae |
| Southern bean mosaic virus | SBMV | Sobemovirus | Solemoviridae |
| Southern cowpea mosaic virus | SCPMV | Sobemovirus | Solemoviridae |
| Soybean chlorotic blotch virus | SbCBV | Begomovirus | Geminiviridae |
| Squash chlorotic leaf spot virus | SCLSV | Torradovirus | Secoviridae |
| Stipagrostis-associated virus | SaV | N/A | N/A |
| Sudan watermelon mosaic virus | SuWMV | Potyvirus | Potyviridae |
| Sugarcane mosaic virus | SCM | Potyvirus | Potyviridae |
| Sugarcane streak Egypt Virus | SSEV | Mastrevirus | Geminiviridae |
| Sugarcane white streak Virus | SCWSV | Mastrevirus | Geminiviridae |
| Sweet potato chlorotic fleck virus | SPCFV | Carlavirus | Betaflexiviridae |
| Sweet potato chlorotic stunt virus | SPCSV | Crinivirus | Closteroviridae |
| Sweet potato feathery mottle virus | SPFMV | Potyvirus | Potyviridae |
| Sweet potato leaf curl virus | SPLCV | Begomovirus | Geminiviridae |
| Sweet potato leaf curl Sao Paulo virus | SPLCSPV | Begomovirus | Geminiviridae |
| Sweet potato leaf curl Uganda virus | SPLCUV | Begomovirus | Geminiviridae |
| Sweet potato mosaic virus | SPMV | Begomovirus | Geminiviridae |
| Sweet potato pakakuy virus | SPPV | Badnavirus | Caulimoviridae |
| Sweet potato virus 2 | SPV2 | Potyvirus | Potyviridae |
| Sweet potato virus C | SPVC | Potyvirus | Potyviridae |
| Sweet potato virus G | SPVG | Potyvirus | Potyviridae |
| Sweet potato symptomless virus 1 | SPSMV-1 | Mastrevirus | Geminiviridae |
| Telfairia mosaic virus | TelMV | Begomovirus | Geminiviridae |
| Tobacco mottle virus | TMoV | Umbravirus | Tombusviridae |
| Tomato chlorosis virus | ToCV | Crinivirus | Closteroviridae |
| Tomato leaf curl Uganda virus | ToLCUV | Begomovirus | Geminiviridae |
| Tomato mosaic virus | ToMV | Tobamovirus | Virgaviridae |
| Tomato spotted wilt orthotospovirus | TSWV | Orthotospovirus | Tospoviridae |
| Turnip yellows virus | TuYV | Polerovirus | Luteoviridae |
| Ugandan cassava brown streak virus | UCBSV | Ipomovirus | Potyviridae |
| Ugandan cassava brown streak virus Comoros | UCBSV-KM | Ipomovirus | Potyviridae |
| West African Asystasia virus 1 | WAAV1 | Begomovirus | Geminiviridae |
| Yam mosaic virus | YMV | Potyvirus | Potyviridae |
| Yam virus Y | YVY | N/A | Betaflexiviridae. |
| Zea mays chrysovirus 1 | ZMCV1 | Alphachrysovirus | Chrysoviridae |
| Zucchini shoestring virus | ZSSV | Potyvirus | Potyviridae |
| Year ! | Country # | Sequencing Platform | Country Where HTS Was Performed | Nucleic Acid Target | Host Plant | Plant-Infecting Virus Identified | References |
|---|---|---|---|---|---|---|---|
| 2020 | Zambia | Illumina MiSeq | South Africa | Total RNA | Common bean | SBMV*; BCMNV; BCMV; CABMV; CMV | [45] |
| 2020 | South Africa | Illumina NovaSeq | South Africa | Ribo-depleted RNA | Japanese plum (Prunus salicina) | PBNSPaV* | [46] |
| 2020 | South Africa | Illumina MiSeq | South Africa | Ribo-depleted RNA | Ornithogalum thyrsoides | OrMV; OV-3; OV-5+ | [47] |
| 2020 | South Africa | Illumina NextSeq | South Africa | Ribo-depleted RNA | ivy (Hedera helix) | IRSaV+ | [48] |
| 2020 | Zimbabwe | Illumina HiSeq | South Africa | Ribo-depleted RNA | Dendranthema morifolium | TSWV | [49] |
| 2020 | Zimbabwe | Illumina HiSeq | South Africa | Ribo-depleted RNA | Zucchini | ZSSV* | [50] |
| 2020 | Kenya | Illumina MiSeq | Kenya | Genomic DNA | Tomato | CpCDV-T+ | [51] |
| 2020 | Kenya | Illumina MiSeq | Kenya | mRNA | Papaya | MWMV*; CPMMV*; PaMV+ PMMaV+ | [52] |
| 2020 | Kenya | Illumina HiSeq | South Africa | Ribo-depleted RNA | Papaya | MWMV; PaVA+ | [53] |
| 2020 | Kenya; Rwanda; Tanzania; Uganda | Illumina MiSeq | Kenya | mRNA | Sweet potato | SPFMV | [54] |
| 2020 | Rwanda | Illumina MiSeq | USA | Ribo-depleted RNA | Maize | MCMV; SCMV; MSV; MYDV-RMV2; MYDV-like; MaYMV; BYDV; MATV+; MPtV-1+; ZMCV1*; MaBV+ | [55] |
| 2019 | Tanzania | Illumina MiSeq | Kenya | Total RNA | Maize | MCMV; SCMV; MSV; MYDV-RMV; BYDV; MDMV; SrMV | [56] |
| 2019 | Tanzania | Illumina HiSeq | South Africa | Ribo-depleted RNA | Maize | MaPV+; MCMV; MSV; MaYMV | [57] |
| 2019 | Tanzania | Illumina HiSeq | South Africa | Ribo-depleted RNA | Maize | MATV-1-Tanz*; MATV-4-Tanz+; | [58] |
| 2019 | Tanzania | Illumina HiSeq | South Africa | Ribo-depleted RNA | Maize | MMaV+; MCMV; MaYMV | [59] |
| 2019 | Tanzania | Illumina HiSeq | South Africa | Ribo-depleted RNA | Maize | MaYMV*; MCMV | [60] |
| 2019 | South Africa | Illumina HiSeq | South Africa | Ribo-depleted RNA | Tomato | ToCV; Other viruses not mentioned | [61] |
| 2019 | South Africa | Illumina MiSeq | South Africa | Ribo-depleted RNA; RCA dsDNA | Sweet potato | SPFMV; SPVG; SPVC; SPV2; SPLCSPV; SPMV | [62] |
| 2019 | South Africa | 454 pyrosequencing | USA | VANA | Arctopus echinatus; Lolium perenne; Panicum ecklonii; Stipagrostis sp. | AeaV; SaV; PeaV; LpaV | [63] |
| 2019 | Zimbabwe | Illumina HiSeq | South Africa | Ribo-depleted RNA | Garlic | IYSV | [64] |
| 2019 | Mozambique | Illumina MiSeq | South Africa | Ribo-depleted RNA | Cassava | CBSV; UCBSV | [65] |
| 2019 | Comoros | Illumina Hiseq | Switzerland | siRNAs; VANA; dsRNAs | Cassava | UCBSV-KM+; CBSV* | [66] |
| 2019 | Sudan | Illumina MiSeq | Switzerland | small RNAs | Cucurbit | CVYV | [67] |
| 2019 | Kenya | Illumina MiSeq | Kenya | Ribo-depleted RNA | Pumpkin | PuPV+; MWMV | [68] |
| 2019 | Kenya | Illumina MiSeq | Kenya | Total RNA | Groundnut | GRAV; Other viruses not mentioned | [69] |
| 2019 | Kenya | Illumina MiSeq | Kenya | mRNA | Common bean | BCMNV | [70] |
| 2019 | Kenya | Illumina HiSeq | South Korea | RCA dsDNA | Common bean | PVBV*; BCMNV | [71] |
| 2019 | Kenya | Illumina HiSeq | South Korea | Ribo-depleted RNA | Common bean; Cowpea | BCMNV; CABMV | [72] |
| 2019 | Kenya | Illumina MiSeq | Kenya | Ribo-depleted RNA | Passion fruit | CABMV | [73] |
| 2019 | Uganda | Illumina MiSeq | Kenya | Ribo-depleted RNA | Tannia (Xanthosoma sp.) | CMV* | [74] |
| 2019 | Uganda | Illumina MiSeq | Kenya | Total RNA | Passion fruit | PV+ | [75] |
| 2019 | Nigeria | Illumina HiSeq | UK | Total RNA | Dioscorea rotundata (Yam) | YMV+; DBRTV3-[2RT]+; DBRTV3-[3RT]+ | [76] |
| 2019 | Nigeria; Ghana | Illumina HiSeq | UK | Total RNA | Dioscorea rotundata | YVY+; YMV; DMaV; Yam badnavirus | [77] |
| 2018 | Zimbabwe | Illumina MiSeq | South Africa | Ribo-depleted RNA | Tomato | ToMV | [78] |
| 2018 | South Africa | Illumina MiSeq | South Africa | RCA dsDNA | Sweet Potato | SPLCV+; SPLCSPV; SPMV | [79] |
| 2018 | South Africa | Illumina MiSeq | South Africa | Ribo-depleted RNA; RCA dsDNA | Sweet potato | SPCSV; SPFMV; SPVC; SPVG; SPMV; SPLCSPV | [80] |
| 2018 | South Africa | Illumina MiSeq | South Africa | siRNAs | Sweet potato | SPPV* | [81] |
| 2018 | Kenya | Illumina Hiseq | South Korea | Ribo-depleted RNA | Sweet potato | SPCFV; SPFMV; SPVC; SPCSV | [82] |
| 2018 | Kenya | Illumina Hiseq | South Korea | Ribo-depleted RNA | Groundnut | GRV | [83] |
| 2018 | Kenya | Illumina MiSeq | Kenya | Total RNA | Common bean | PvEV-1+; PvEV-2+ BCMNV, CMV | [84] |
| 2018 | Uganda | Illumina MiSeq | Kenya | mRNA | Cowpea | CABMV | [85] |
| 2018 | Tanzania | Illumina HiSeq | Switzerland | siRNAs | Common Bean | Umbraviruses+; ToLCUV*; PeMoV; BnYDV; TMmV SBMV*; BCMNV; BCMV; PvEV-1; PvEV-2; CpMMV; CMV; CABMV | [86] |
| 2018 | Uganda; Kenya; Rwanda; Tanzania | Illumina Hiseq | USA | mRNA | Maize | MYDV-like polerovirus*; East African JGMV; SCMV; MCMV | [87] |
| 2018 | Kenya | Illumina MiSeq | Kenya | Total RNA | Maize; Sorghum; Napier grass | Hubei Poty-like virus 1; BVG; ScaMV; MCMV; SCMV; MSV; MYDV-RMV; JGMV | [88] |
| 2018 | Côte d’Ivoire; Ghana | Illumina HiSeq Illumina MiSeq | France | RCA dsDNA | Cacao | CSSCEV+; CSSGMV+; CSSGNV+; CSSGQV+; CSSGRV+; CSSV; CSSTAV; CSSCDV | [89] |
| 2017 | Benin; Mali | Illumina MiSeq | Germany | Ribo-depleted RNA | Solanum aethiopicum; Pepper (Capsicum spp.) | AeYV+ | [90] |
| 2017 | Malawi | Illumina MiSeq | Germany | Ribo-depleted RNA | Cassava | UCBSV | [91] |
| 2017 | Tanzania | Illumina HiSeq | Switzerland | siRNAs | Common bean | PvEV-1; PvEV-2; CPMMV | [92] |
| 2017 | Sudan | Illumina HiSeq | Switzerland. | siRNAs | Cucumber | SuWMV+ | [93] |
| 2017 | South Africa | Illumina HiSeq | South Africa | Ribo-depleted RNA | Zucchini | PABYV+ | [94] |
| 2017 | South Africa | Illumina HiSeq | South Africa | Ribo-depleted RNA | Potato | PVY | [95] |
| 2016 | South Africa | Illumina HiSeq | South Africa | Total RNA | Cabbage | TuYV* | [96] |
| 2016 | South Africa | Illumina HiSeq | South Africa | Ribo-depleted RNA | Pattypan; Zucchini | ZSSV+; MWMV | [97,98] |
| 2016 | Sudan | Illumina HiSeq | Switzerland | siRNAs | Squash | SCLSV+ | [99] |
| 2016 | Sudan | Illumina HiSeq | Switzerland | siRNAs | Wild cucurbit | CocMoV | [100] |
| 2016 | Cameroon; Togo | Illumina HiSeq | USA | RCA dsDNA | Cassava | SbCBV*; WAAV1*; ACMV | [101] |
| 2016 | Cameroon | Illumina MiSeq | USA | RCA dsDNA | Pumpkin | TelMV+ | [102] |
| 2016 | Benin | Illumina MiSeq | USA | RCA dsDNA | Cotton | CYMV+ | [103] |
| 2016 | Burkina Faso | 454 pyrosequencing | USA | VANA | Cowpea | CPPV-1+; CPPV-2+; CPTV-1+; CPTV-2+; CPaMV-1+; SCPMV*; BCMV-BlCM; CPMoV; CABMV | [104] |
| 2015 | South Africa | Illumina HiSeq | South Africa | dsRNAs | Grapevine | GLRaV-3 isolate GH24 | [105] |
| 2015 | South Africa | Illumina HiSeq | South Africa | dsRNAs | Grapevine | GVF; GLRaV-3 | [106] |
| 2015 | Tanzania | Illumina MiSeq | Kenya | Ribo-depleted RNA | Cassava | CBSV; UCBSV+ | [107] |
| 2015 | Ethiopia | Illumina MiSeq Illumina HiSeq | UK | Total RNA | Maize | MCMV*; SCMV | [108] |
| 2014 | Rwanda | Illumina MiSeq | UK | Total RNA | Maize | MCMV*; SCMV* | [109] |
| 2014 | Tanzania | Illumina HiSeq | Switzerland | siRNAs | Sweet potato | SPPV; SPSMV-1; SPLCUV; SPCSV; SPFMV; SPLCSPV+; SPCFV | [110] |
| 2014 | Egypt | Illumina HiSeq; Roche 454 GS FLX Titanium | USA | siRNAs; VANA | Sugarcane | SSEV; SCWSV+ | [111] |
| 2013 | Kenya | Roche 454 GS-FLX+ | UK | Total RNA; dsRNAs | Maize | MCMV*; SCMV | [112] |
| 2012 | South Africa | Illumina HiSeq | South Africa | dsRNAs | Grapevine. | GEEV+ | [113] |
| 2010 | Uganda; Tanzania | Roche GS-FLX | UK | Total RNA | Cassava | CBSV; UCBSV | [114] |
| 2010 | South Africa. | Illumina | South Africa | dsRNAs | Grapevine | GVE+; GRSPaV; GVA; GLRaV-3 | [115] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibaba, J.D.; Gubba, A. High-Throughput Sequencing Application in the Diagnosis and Discovery of Plant-Infecting Viruses in Africa, A Decade Later. Plants 2020, 9, 1376. https://doi.org/10.3390/plants9101376
Ibaba JD, Gubba A. High-Throughput Sequencing Application in the Diagnosis and Discovery of Plant-Infecting Viruses in Africa, A Decade Later. Plants. 2020; 9(10):1376. https://doi.org/10.3390/plants9101376
Chicago/Turabian StyleIbaba, Jacques Davy, and Augustine Gubba. 2020. "High-Throughput Sequencing Application in the Diagnosis and Discovery of Plant-Infecting Viruses in Africa, A Decade Later" Plants 9, no. 10: 1376. https://doi.org/10.3390/plants9101376