Can Inositol Pyrophosphates Inform Strategies for Developing Low Phytate Crops?


Abstract
:
1. Introduction

2. InsP6 Synthesis: The Lipid-Dependent Pathway
3. InsP6 Synthesis: The Lipid-Independent Pathway
4. PP-InsP Synthetic Pathway
5. How Do PP-InsPs Function in Plants?
6. Consequences for Targeting InsP6 for Reduction
7. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Livermore, T.; Azevedo, C.; Kolozsvari, B.; Wilson, M.; Saiardi, A. Phosphate, inositol and polyphosphates. Biochem. Soc. Trans. 2016, 44, 253–259. [Google Scholar] [CrossRef]
- Chakraborty, A. The inositol pyrophosphate pathway in health and diseases. Biol. Rev. Camb. Philos. Soc. 2019, 93, 1203–1227. [Google Scholar] [CrossRef]
- Shears, S.B. Intimate connections: Inositol pyrophosphates at the interface of metabolic regulation and cell signaling. J. Cell Physiol. 2018, 233, 1897–1912. [Google Scholar] [CrossRef]
- Nagy, R.; Grob, H.; Weder, B.; Green, P.; Klein, M.; Frelet-Barrand, A.; Schjoerring, J.K.; Brearley, C.; Martinoia, E. The Arabidopsis ATP-binding cassette protein AtMRP5/AtABCC5 is a high affinity inositol hexakisphosphate transporter involved in guard cell signaling and phytate storage. J. Biol. Chem. 2009, 284, 33614–33622. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.; Rangarajan, P.; Donahue, J.L.; Williams, S.P.; Land, E.S.; Mandal, M.K.; Phillippy, B.Q.; Perera, I.Y.; Raboy, V.; Gillaspy, G.E.; et al. Two inositol hexakisphosphate kinases drive inositol pyrophosphate synthesis in plants. Plant J. 2014, 80, 642–653. [Google Scholar] [CrossRef]
- Raboy, V. Seed Total Phosphate and Phytic Acid. In Molecular Genetic Approaches to Maize Improvement; Biotechnology in Agriculture and Forestry; Kriz, A.L., Larkins, B.A., Eds.; Springer: Berlin, Germany, 2009; pp. 41–53. [Google Scholar]
- Otegui, M.S.; Capp, R.; Staehelin, L.A. Developing Seeds of Arabidopsis Store Different Minerals in Two Types of Vacuoles and in the Endoplasmic Reticulum. Plant Cell 2002, 14, 1311–1327. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-Y.; Huang, T.-K.; Kuo, H.-F.; Chiou, T.-J. Role of vacuoles in phosphorus storage and remobilization. J. Exp. Bot. 2017, 68, 3045–3055. [Google Scholar] [CrossRef]
- Bohn, L.; Meyer, A.S.; Rasmussen Søren, K. Phytate: Impact on environment and human nutrition. A challenge for molecular breeding. J. Zhejiang Univ. Sci. B 2008, 9, 165–191. [Google Scholar] [CrossRef] [Green Version]
- Raboy, V. Seeds for a better future: ‘Low phytate’ grains help to overcome malnutrition and reduce pollution. Trends Plant Sci. 2001, 6, 458–462. [Google Scholar] [CrossRef]
- Cowieson, A.J.; Acamovic, T.; Bedford, M.R. Phytic acid and phytase: Implications for protein utilization by poultry. Poult. Sci. 2006, 85, 878–885. [Google Scholar] [CrossRef]
- Abelson, P.H. A Potential Phosphate Crisis. Science 1999, 283, 2015. [Google Scholar] [CrossRef]
- Sharpley, A.N.; Withers, P.J.A. The environmentally-sound management of agricultural phosphorus. Fertil. Res. 1994, 39, 133–146. [Google Scholar] [CrossRef]
- Raboy, V. Progress in Breeding Low Phytate Crops. J. Nutr. 2002, 132, 503S–505S. [Google Scholar] [CrossRef]
- Isayenkov, S. Plant vacuoles: Physiological roles and mechanisms of vacuolar sorting and vesicular trafficking. Cytol. Genet. 2014, 48, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Perera, I.; Fukushima, A.; Tatsuki, A.; Horiguchi, G.; Saman, S.; Naoki, H. Expression regulation of myo-inositol 3-phosphate synthase 1 (INO1) in determination of phytic acid accumulation in rice grain. Sci. Rep. 2019, 9, 14866. [Google Scholar] [CrossRef]
- O’Dell, B.L.; De Boland, A.R.; Koirtyohann, S.R. Distribution of phytate and nutritionally important elements among the morphological components of cereal grains. J. Agric. Food Chem. 1972, 20, 718–723. [Google Scholar] [CrossRef]
- Erdman, J. Oilseed phytates: Nutritional implications. J. Am. Oil Chem. Soc. 1979, 56, 736–741. [Google Scholar] [CrossRef]
- Shears, S.B. Inositol pyrophosphates: Why so many phosphates? Adv. Biol. Regul. 2015, 57, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Williams, S.P.; Gillaspy, G.E.; Perera, I.Y. Biosynthesis and possible functions of inositol pyrophosphates in plants. Front. Plant Sci. 2015, 6, 67. [Google Scholar] [CrossRef] [Green Version]
- Odom, A.R.; Stahlberg, A.; Wente, S.R.; York, J.D. A Role for Nuclear Inositol 1,4,5-Trisphosphate Kinase in Transcriptional Control. Science 2000, 287, 2026–2029. [Google Scholar] [CrossRef]
- Perera, N.M.; Michell, R.H.; Dove, S.K. Hypo-osmotic Stress Activates Plc1p-dependent Phosphatidylinositol 4,5-Bisphosphate Hydrolysis and Inositol Hexakisphosphate Accumulation in Yeast. J. Biol. Chem. 2004, 279, 5216–5226. [Google Scholar] [CrossRef] [Green Version]
- York, J.; Odom, A.R.; Murphy, R.; Ives, E.; Wente, S.R. A Phospholipase C-Dependent Inositol Polyphosphate Kinase Pathway Required for Efficient Messenger RNA Export. Science 1999, 285, 96–100. [Google Scholar] [CrossRef]
- Fujii, M.; York, J.D. A role for rat inositol polyphosphate kinases rIPK2 and rIPK1 in inositol pentakisphosphate and inositol hexakisphosphate production in rat-1 cells. J. Biol. Chem. 2005, 280, 1156–1164. [Google Scholar] [CrossRef] [Green Version]
- Seeds, A.M.; Sandquist, J.C.; Spana, E.P.; York, J.D. A molecular basis for inositol polyphosphate synthesis in Drosophila melanogaster. J. Biol. Chem. 2004, 279, 47222–47232. [Google Scholar] [CrossRef] [Green Version]
- Stephens, L.R.; Irvine, R.F. Stepwise phosphorylation of myo-inositol leading to myo-inositol hexakisphosphate in Dictyostelium. Nature 1990, 346, 580–583. [Google Scholar] [CrossRef]
- Brearley, C.A.; Hanke, D.E. Metabolic evidence for the order of addition of individual phosphate esters in the myo-inositol moiety of inositol hexakisphosphate in the duckweed Spirodela polyrhiza L. Biochem. J. 1996, 314, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Desfougères, Y.; Wilson, M.S.C.; Laha, D.; Miller, G.J.; Saiardi, A. ITPK1 mediates the lipid-independent synthesis of inositol phosphates controlled by metabolism. Proc. Natl. Acad. Sci. USA 2019, 116, 24551–24561. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.J. Inositol trisphosphate and calcium signalling. Nature 1993, 361, 315–325. [Google Scholar] [CrossRef]
- Stevenson-Paulik, J.; Bastidas, R.; Chiou, S.-T.; Frye, R.; York, J. Generation of phytate-free seeds in Arabidopsis through disruption of inositol polyphosphate kinases. Proc. Natl. Acad. Sci. USA 2005, 102, 12612–12617. [Google Scholar] [CrossRef] [Green Version]
- Stevenson-Paulik, J.; Odom, A.R.; York, J.D. Molecular and Biochemical Characterization of Two Plant Inositol Polyphosphate 6-/3-/5-Kinases. J. Biol. Chem. 2002, 277, 42711–42718. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; van Wijk, R.; Shahbaz, M.; Roels, W.; van Schooten, B.; Vermeer, J.E.M.; Zarza, X.; Guardia, A.; Scuffi, D.; García-Mata, C.; et al. Arabidopsis Phospholipase C3 is Involved in Lateral Root Initiation and ABA Responses in Seed Germination and Stomatal Closure. Plant Cell Physiol. 2018, 59, 469–486. [Google Scholar] [CrossRef]
- Kuo, H.-F.; Hsu, Y.-Y.; Lin, W.-C.; Chen, K.-Y.; Munnik, T.; Brearley, C. Arabidopsis inositol phosphate kinases IPK1 and ITPK1 constitute a metabolic pathway in maintaining phosphate homeostasis. Plant J. 2018, 95, 613–630. [Google Scholar] [CrossRef] [Green Version]
- Laha, D.; Johnen, P.; Azevedo, C.; Dynowski, M.; Weiß, M.; Capolicchio, S.; Mao, H.; Iven, T.; Steenbergen, M.; Freyer, M.; et al. VIH2 Regulates the Synthesis of Inositol Pyrophosphate InsP8 and Jasmonate-Dependent Defenses in Arabidopsis. Plant Cell 2015, 27, 1082–1097. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-I.; Tai, T.H. Identification of genes necessary for wild-type levels of seed phytic acid in Arabidopsis thaliana using a reverse genetics approach. Mol. Genet. Genom. 2011, 286, 119–133. [Google Scholar] [CrossRef]
- Laha, D.; Parvin, N.; Hofer, A.; Giehl, R.F.H.; Fernandez-Rebollo, N.; von Wirén, N.; Saiardi, A.; Jessen, H.J.; Schaaf, G. Arabidopsis ITPK1 and ITPK2 Have an Evolutionarily Conserved Phytic Acid Kinase Activity. ACS Chem. Biol. 2019, 14, 2127–2133. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Lau, K.; Puschmann, R.; Harmel, R.K.; Zhang, Y.; Pries, V.; Gaugler, P.; Broger, L.; Dutta, A.K.; Jessen, H.J.; et al. Two bifunctional inositol pyrophosphate kinases/phosphatases control plant phosphate homeostasis. eLife 2019, 8, e43582. [Google Scholar] [CrossRef]
- Adepoju, O.; Williams, S.P.; Craige, B.; Cridland, C.A.; Sharpe, A.K.; Brown, A.M.; Land, E.; Perera, I.Y.; Mena, D.; Sobrado, P.; et al. Inositol Trisphosphate Kinase and Diphosphoinositol Pentakisphosphate Kinase Enzymes Constitute the Inositol Pyrophosphate Synthesis Pathway in Plants (pre-print). bioRxiv 2019, 724914. [Google Scholar] [CrossRef] [Green Version]
- Sweetman, D.; Johnson, S.; Caddick, S.E.K.; Hanke, D.E.; Brearley, C.A. Characterization of an Arabidopsis inositol 1,3,4,5,6-pentakisphosphate 2-kinase (AtIPK1). Biochem. J. 2006, 394, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Wang, H.; Hazebroek, J.; Ertl, D.S.; Harp, T. The maize low-phytic acid 3 encodes a myo-inositol kinase that plays a role in phytic acid biosynthesis in developing seeds. Plant J. 2005, 42, 708–719. [Google Scholar] [CrossRef]
- Kim, S.-I.; Tai, T.H. Genetic analysis of two OsLpa1-like genes in Arabidopsis reveals that only one is required for wild-type seed phytic acid levels. Planta 2010, 232, 1241–1250. [Google Scholar] [CrossRef]
- Kishor, D.S.; Lee, C.; Lee, D.; Venkatesh, J.; Seo, J.; Chin, J.H.; Jin, Z.; Hong, S.-K.; Ham, J.-K.; Koh, H.J. Novel allelic variant of Lpa1 gene associated with a significant reduction in seed phytic acid content in rice (Oryza sativa L.). PLoS ONE 2019, 14, e0209636. [Google Scholar] [CrossRef] [Green Version]
- Stiles, A.R.; Qian, X.; Shears, S.B.; Grabau, E.A. Metabolic and signaling properties of an Itpk gene family in Glycine max. FEBS Lett. 2008, 582, 1853–1858. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Wang, H.; Wu, Y.; Hazebroek, J.; Meeley, R.B.; Ertl, D.S. The Maize Low-Phytic Acid Mutant lpa2 Is Caused by Mutation in an Inositol Phosphate Kinase Gene. Plant Physiol. 2003, 131, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Sweetman, D.; Stavridou, I.; Johnson, S.; Green, P.; Caddick, S.E.K.; Brearley, C.A. Arabidopsis thaliana inositol 1,3,4-trisphosphate 5/6-kinase 4 (AtITPK4) is an outlier to a family of ATP-grasp fold proteins from Arabidopsis. FEBS Lett. 2007, 581, 4165–4171. [Google Scholar] [CrossRef] [Green Version]
- Majerus, P.W.; Wilson, D.B.; Zhang, C.; Nicholas, P.J.; Wilson, M.P. Expression of inositol 1,3,4-trisphosphate 5/6-kinase (ITPK1) and its role in neural tube defects. Adv. Enzyme Regul. 2010, 50, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Couso, I.; Evans, B.S.; Li, J.; Liu, Y.; Ma, F.; Diamond, S.; Allen, D.K.; Umen, J.G. Synergism between Inositol Polyphosphates and TOR Kinase Signaling in Nutrient Sensing, Growth Control, and Lipid Metabolism in Chlamydomonas. Plant Cell 2016, 28, 2026–2042. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Williams, J.; Cho, J.; Falck, J.R.; Shears, S. Purification, Sequencing, and Molecular Identification of a Mammalian PP-InsP5 Kinase That Is Activated When Cells Are Exposed to Hyperosmotic Stress. J. Biol. Chem. 2007, 282, 30763–30775. [Google Scholar] [CrossRef] [Green Version]
- Fridy, P.C.; Otto, J.C.; Dollins, D.E.; York, J.D. Cloning and characterization of two human VIP1-like inositol hexakisphosphate and diphosphoinositol pentakisphosphate kinases. J. Biol. Chem. 2007, 282, 30754–30762. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Fridy, P.C.; Ribeiro, A.A.; Choi, J.H.; Barma, D.K.; Vogel, G.; Falck, J.R.; Shears, S.B.; York, J.D.; Mayr, G.W. Structural Analysis and Detection of Biological Inositol Pyrophosphates Reveal That the Family of VIP/Diphosphoinositol Pentakisphosphate Kinases Are 1/3-Kinases. J. Biol. Chem. 2009, 284, 1863–1872. [Google Scholar] [CrossRef] [Green Version]
- Mulugu, S.; Bai, W.; Fridy, P.C.; Bastidas, R.J.; Otto, J.C.; Dollins, D.E.; Haystead, T.A.; Ribeiro, A.A.; York, J.D. A conserved family of enzymes that phosphorylate inositol hexakisphosphate. Science 2007, 316, 106–109. [Google Scholar] [CrossRef]
- Weaver, J.D.; Wang, H.; Shears, S.B. The kinetic properties of a human PPIP5K reveal that its kinase activities are protected against the consequences of a deteriorating cellular bioenergetic environment. Biosci. Rep. 2013, 33, e00022. [Google Scholar] [CrossRef]
- Draškovič, P.; Saiardi, A.; Bhandari, R.; Burton, A.; Ilc, G.; Kovačevič, M.; Snyder, S.H.; Podobnik, M. Inositol Hexakisphosphate Kinase Products Contain Diphosphate and Triphosphate Groups. Chem. Biol. 2008, 15, 274–286. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Calderon-Villalobos, L.I.A.; Sharon, M.; Zheng, C.; Robinson, C.V.; Estelle, M.; Zheng, N. Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature 2007, 446, 640–645. [Google Scholar] [CrossRef]
- Wild, R.; Gerasimaite, R.; Jung, J.-Y.; Truffault, V.; Pavlovic, I.; Schmidt, A.; Saiardi, A.; Jessen, H.J.; Poirier, Y.; Hothorn, M.; et al. Control of eukaryotic phosphate homeostasis by inositol polyphosphate sensor domains. Science 2016, 352, 986–990. [Google Scholar] [CrossRef] [Green Version]
- Rouached, H.; Arpat, A.B.; Poirier, Y. Regulation of Phosphate Starvation Responses in Plants: Signaling Players and Cross-Talks. Mol. Plant 2010, 3, 288–299. [Google Scholar] [CrossRef]
- Chien, P.-S.; Chiang, C.-P.; Leong, S.J.; Chiou, T.-J. Sensing and Signaling of Phosphate Starvation—From Local to Long Distance. Plant Cell Physiol. 2018, 59, 1714–1722. [Google Scholar] [CrossRef]
- Azevedo, C.; Saiardi, A. Eukaryotic Phosphate Homeostasis: The Inositol Pyrophosphate Perspective. Trends Biochem. Sci. 2017, 42, 219–231. [Google Scholar] [CrossRef]
- Ried, M. Inositol pyrophosphates promote the interaction of SPX domains with the coiled-coil motif of PHR transcription factors to regulate plant phosphate homeostasis (pre-print). bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Puga, M.I.; Mateos, I.; Charukesi, R.; Wang, Z.; Franco-Zorrilla, J.M.; de Lorenzo, L.; Irigoyen, M.L.; Masiero, S.; Bustos, R.; Rodríguez, J.; et al. SPX1 is a phosphate-dependent inhibitor of PHOSPHATE STARVATION RESPONSE 1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 14947–14952. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Ma, G.; Sui, L.; Wei, M.; Satheesh, V.; Zhang, R.; Ge, S.; Li, J.; Zhang, T.-E.; Wittwer, C.; et al. Inositol Pyrophosphate InsP8 Acts as an Intracellular Phosphate Signal in Arabidopsis. Mol. Plant 2019, 12, 1463–1473. [Google Scholar] [CrossRef]
- Kuo, H.-F.; Chang, T.-Y.; Chiang, S.-F.; Wang, W.-D.; Charng, Y.; Chiou, T.-J. Arabidopsis inositol pentakisphosphate 2-kinase, AtIPK1, is required for growth and modulates phosphate homeostasis at the transcriptional level. Plant J. 2014, 80, 503–515. [Google Scholar] [CrossRef]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, Action, and Interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [Green Version]
- Gray, W.M.; Kepinski, S.; Rouse, D.; Leyser, O.; Estelle, M. Auxin regulates SCF TIR1-dependent degradation of AUX/IAA proteins. Nature 2001, 414, 271–276. [Google Scholar] [CrossRef]
- Tan, X.; Zheng, N. Hormone signaling through protein destruction: A lesson from plants. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E223–E227. [Google Scholar] [CrossRef]
- Sheard, L.B.; Tan, X.; Mao, H.; Withers, J.; Ben-Nissan, G.; Hinds, T.R.; Kobayashi, Y.; Hsu, F.-F.; Sharon, M.; Browse, J.; et al. Jasmonate perception by inositol phosphate-potentiated COI1-JAZ co-receptor. Nature 2010, 468, 400–405. [Google Scholar] [CrossRef]
- Browse, J. Jasmonate Passes Muster: A Receptor and Targets for the Defense Hormone. Ann. Rev. Plant Biol. 2009, 60, 183–205. [Google Scholar] [CrossRef]
- Chini, A.; Fonseca, S.; Fernández, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; García-Casado, G.; López-Vidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef]
- Cui, M.; Du, J.; Yao, X. The Binding Mechanism Between Inositol Phosphate (InsP) and the Jasmonate Receptor Complex: A Computational Study. Front. Plant Sci. 2018, 9, 963. [Google Scholar] [CrossRef]
- Mosblech, A.; Thurow, C.; Gatz, C.; Feussner, I.; Heilmann, I. Jasmonic acid perception by COI1 involves inositol polyphosphates in Arabidopsis thaliana. Plant J. 2011, 65, 949–957. [Google Scholar] [CrossRef]
- Yu, H.; Wu, J.; Xu, N.; Peng, M. Roles of F-box proteins in plant hormone responses. Acta Biochim. Biophys. Sin. Shanghai 2007, 39, 915–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Chu, C. Nitrogen-phosphorus interplay: Old story with molecular tale. New Phytol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Baek, D.; Chun, H.J.; Yun, D.-J.; Kim, M.C. Cross-talk between Phosphate Starvation and Other Environmental Stress Signaling Pathways in Plants. Mol. Cells 2017, 40, 697–705. [Google Scholar]
- Murphy, A.M.; Otto, B.; Brearley, C.A.; Carr, J.P.; Hanke, D.E. A role for inositol hexakisphosphate in the maintenance of basal resistance to plant pathogens. Plant J. 2008, 56, 638–652. [Google Scholar] [CrossRef] [PubMed]
- Reddy, C.S.; Kim, S.-C.; Kaul, T. Genetically modified phytase crops role in sustainable plant and animal nutrition and ecological development: A review. 3 Biotech 2017, 7, 195. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
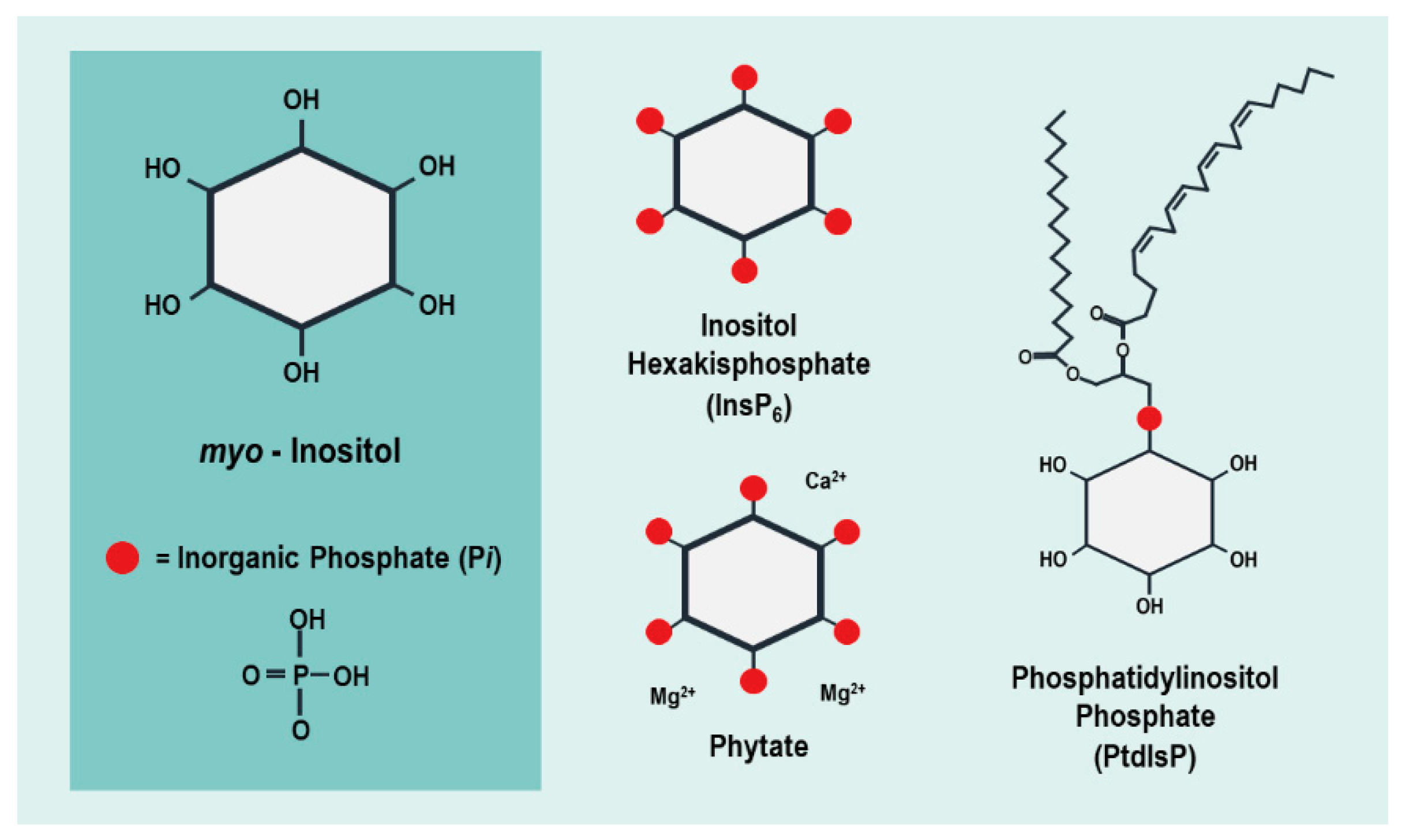{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Mutant | Impact on InsP6 | Impact on PP-InsPs |
|---|---|---|---|
| Lipid-Dependent Pathway | plc | Nine genes; characterized single mutants have no change in InsP6 [32] | No Change [32] |
| ipk2α * | Lethal Knock-Out [30] | Unknown | |
| ipk2β * | 35% reduction in mass seed InsP6; no change in seedling tissue as measured by radiolabeling [30] | Unknown | |
| ipk1 * | 83% reduction in mass seed InsP6; 93% reduction in seedlings as measured by radiolabeling [30] | InsP7 and InsP8 are reduced [33,34] | |
| Lipid-Independent Pathway | mik | 62–66% reduction in mass seed InsP6 [35] | Unknown |
| lpa1 | 47–57% reduction in mass seed InsP6 [35] | Unknown | |
| itpk1–4 | 46% reduction in mass seed InsP6 in Atitpk1 mutants; no changes in Atitpk2 or Atitpk3; 40–51% reduction in Atitpk4 [35] | Atitpk1 and Atitpk4 have reduced InsP7 and InsP8 [33,36] | |
| Phytate Storage | mrp5 | 73–80% reduction in mass seed InsP6 [35]; decreased InsP6 in seeds and vegetative tissue as measured by radiolabeling [4] | Elevated InsP7 and InsP8 [5] |
| PP-InsP Synthetic Pathway | vip1 | No change reported [34,37] | Increased InsP7 and Decreased InsP8 [34,37] |
| vip1/vip2 | No change reported [37] | Increased InsP7 and Decreased InsP8 [37,38] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freed, C.; Adepoju, O.; Gillaspy, G. Can Inositol Pyrophosphates Inform Strategies for Developing Low Phytate Crops? Plants 2020, 9, 115. https://doi.org/10.3390/plants9010115
Freed C, Adepoju O, Gillaspy G. Can Inositol Pyrophosphates Inform Strategies for Developing Low Phytate Crops? Plants. 2020; 9(1):115. https://doi.org/10.3390/plants9010115
Chicago/Turabian StyleFreed, Catherine, Olusegun Adepoju, and Glenda Gillaspy. 2020. "Can Inositol Pyrophosphates Inform Strategies for Developing Low Phytate Crops?" Plants 9, no. 1: 115. https://doi.org/10.3390/plants9010115