Silicification of Root Tissues

 , and
, and 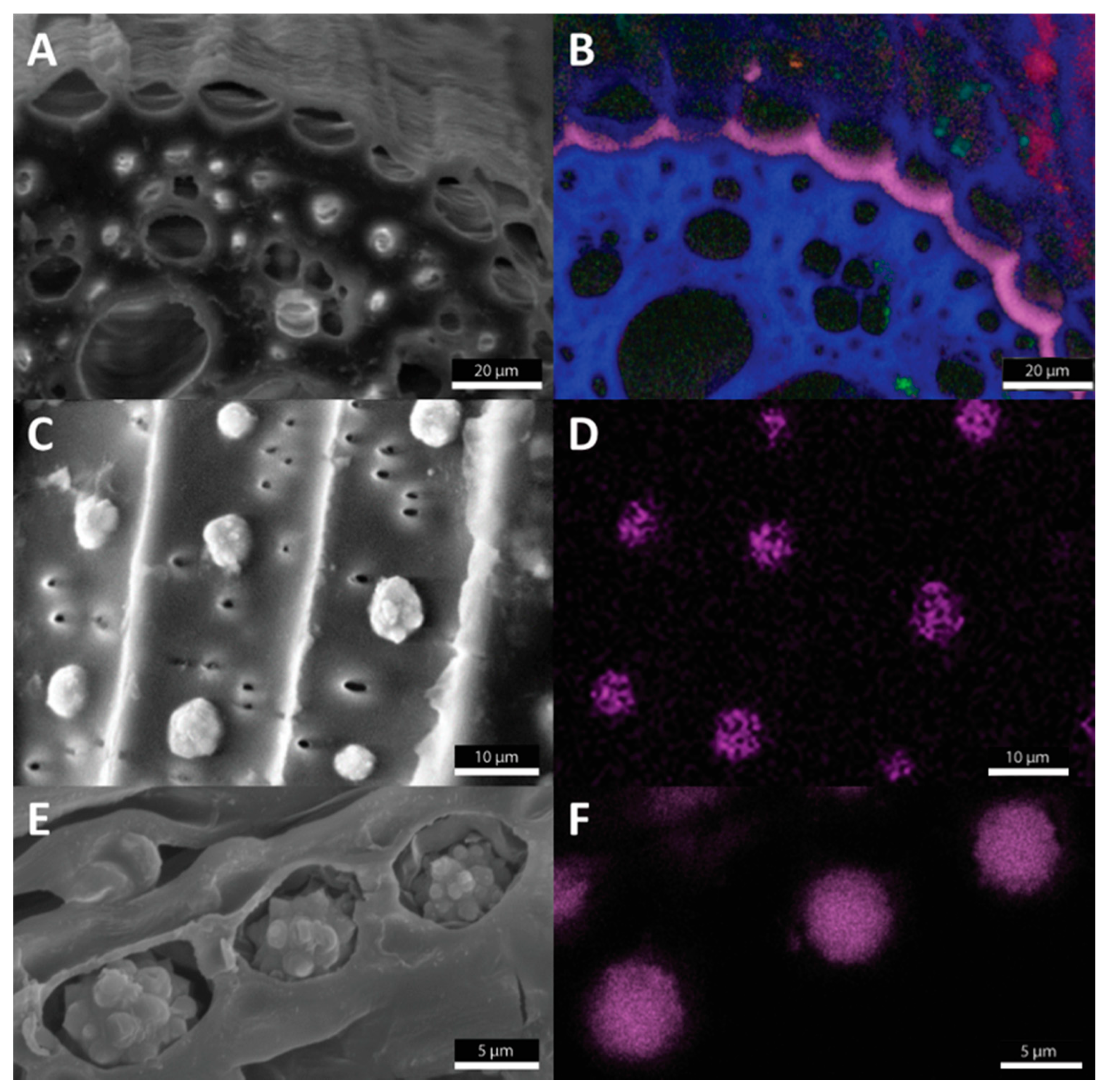{kind=link}
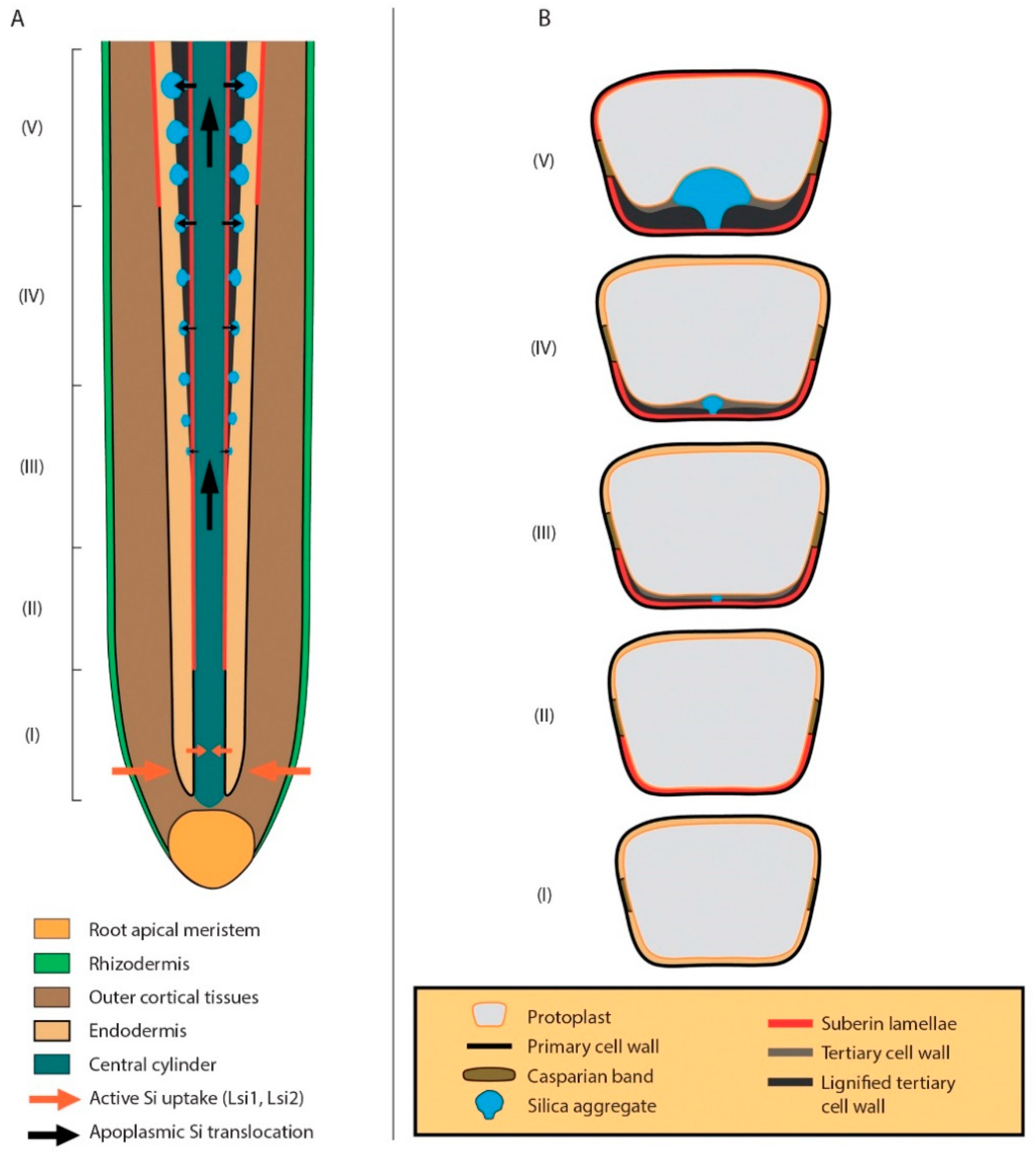{kind=link}
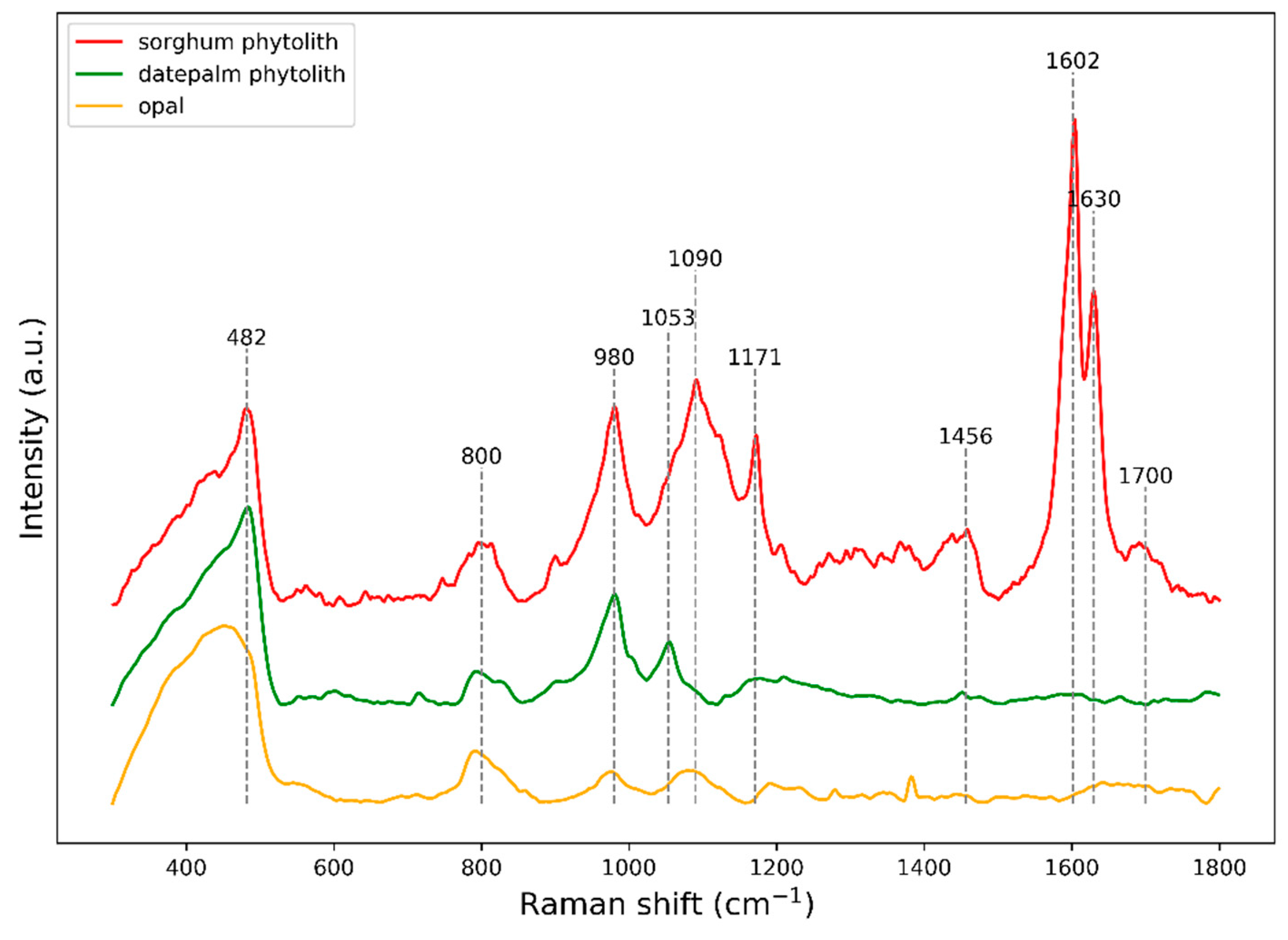{kind=link}
Abstract
:1. Introduction
2. History and Recent Knowledge about Si Deposits in Roots
3. The Root and Its Interaction with Silicon
4. Silicon Transport in Plants
5. Effects of Silicon in Plant Roots during Abiotic Stresses
6. Effects of Silicon in Plant Roots during Biotic Stresses
- (1)
- in plants by modification of gene expression and metabolome resulting in
- a:
- limitation of pathogens spreading via deposition of phenolics and suberin into the cell walls, and thus changed their composition and structure,
- b:
- reduction of oxidative stress via synthesis of enzymes decreasing the amounts of reactive oxygen species,
- c:
- decreased wounding caused by pathogen via increased production and directed secretion of plant enzymes degrading pathogens’ bodies, toxic and allelopathic substances,
- d:
- decreased secretion of effectors involved in compatible plant–pathogen interaction mimicking incompatible interaction and thus causing failure in pathogen attack,
- e:
- improvement of nutrient uptake and distribution via improved interaction with microorganisms involved in arbuscular mycorrhizas,
- f:
- increased biomass production and growth of a plant and in retardation of plant death via induction of pathways releasing energy and substances from its own resources.
- (2)
- in environment by changing the biotic and abiotic factors via plant secreted substances that promote reproduction of beneficial bacteria, actinomycetes, N2-fixing organisms, and silicate solubilizing microorganisms, and thus decreased the number of pathogens.
- (3)
- in pathogens by retardation of its reproduction speed, survival, lifespan, fertility, viability, and attack caused by a decreased amount of nutritious plant food and increased expenses to eliminate intoxication by allelopathic substances.
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ma, J.F.; Takahashi, E. Soil, Fertilizer, and Plant Silicon Research in Japan; Elsevier: Amsterdam, The Netherlands, 2002; p. 294. [Google Scholar]
- Ross, L.; Nababsing, P.; Cheong, Y.W.Y. Residual effect of calcium silicate applied to sugarcane soils. Proc. ISSCT 1974, 15, 539–542. [Google Scholar]
- Savant, N.K.; Snyder, G.H.; Datnoff, L.E. Silicon management and sustainable rice production. Adv. Agron. 1997, 58, 151–199. [Google Scholar]
- Guntzer, F.; Keller, C.; Meunier, J.D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Sistani, K.R.; Savant, N.K.; Reddy, K.C. Effect of rice hull ash silicon on rice seedling growth. J. Plant Nutr. 1997, 20, 195–201. [Google Scholar] [CrossRef]
- Mayland, H.F.; Shewmaker, G.E. Animal health problems caused by silicon and other mineral imbalances. J. Range Manag. 2001, 54, 441–446. [Google Scholar] [CrossRef]
- Jones, L.H.P.; Handreck, K.A. Silica in soils, plants, and animals. Adv. Agron. 1967, 19, 107–149. [Google Scholar]
- Epstein, E. The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. USA 1994, 9, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.A.; Dietz, K.J. Silicon as versatile player in plant and human biology: Overlooked and poorly understood. Front. Plant Sci. 2015, 6, 994. [Google Scholar] [CrossRef] [Green Version]
- Pati, S.; Pal, B.; Badole, S.; Hazra, G.C.; Mandal, B. Effect of silicon fertilization on growth, yield, and nutrient uptake of rice. Commun. Soil Sci. Plant Anal. 2016, 47, 284–290. [Google Scholar] [CrossRef]
- Tubana, B.S.; Babu, T.; Datnoff, L.E. A Review of silicon in soils and plants and its role in US agriculture. History and future perspectives. Soil Sci. 2016, 181, 393–411. [Google Scholar]
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s role in abiotic and biotic plant stresses. Annu. Rev. Phytopathol. 2017, 55, 85–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, H.; Tank, R.V.; Manoli, P. Significance of silicon in fruit crops—A review. Plant Arch. 2017, 17, 769–774. [Google Scholar]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and future prospects on the action mechanisms in alleviating biotic and abiotic stresses in plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef] [PubMed]
- Harrison, T. Paleontology and Geology of Laetoli: Human Evolution in Context: Volume 1: Geology, Geochronology, Paleoecology and Paleoenvironment; Springer Science & Business Media: Berlin, Germany, 2011; p. 402. [Google Scholar]
- Coe, H.H.G.; Osterrieth, M. Synthesis of Some Phytolith Studies in South America (Brazil and Argentina); Nova Science Publishers Inc.: New York, NY, USA, 2014; p. 280. [Google Scholar]
- Halligan, J.E. Soil Fertility and Fertilizers; Reprint. 2013; Forgotten Books: London, UK, 1912; pp. 4–5. [Google Scholar]
- Arnon, D.I.; Stout, P.R. The essenciality of certain elements in minute quantity for plants with special reference to copper. Plant Physiol. 1939, 14, 371–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Nishimura, K.; Takahashi, E. Effect of silicon on the growth of rice plant at different growth stages. Soil Sci. Plant Nutr. 1989, 35, 347–356. [Google Scholar] [CrossRef]
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef]
- Martin-Jézéquel, V.; Hildebrand, M.; Brzezinski, M.A. Silicon metabolism in diatoms: Implication for growth. J. Phycol. 2000, 36, 821–840. [Google Scholar] [CrossRef]
- Yoshida, S.; Ohnishi, Y.; Kitagishi, K. Chemical forms, mobility and deposition of silicon in rice plant. Soil Sci. Plant Nutr. 1962, 8, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A. The transport and function of silicon in plants. Biol. Rev. 1983, 58, 179–207. [Google Scholar] [CrossRef]
- Struve, G.A. De Silicia in Plantis Nonnullis. Ph.D. Thesis, Universitas Litteraria Friderica Guilelma, Berolini, Germany, 1835. [Google Scholar]
- Cruger, H. Westindische Fragmente. Cauto. Bot. Zeitung 1857, 15, 281–292. [Google Scholar]
- Von Mohl, I. über das Kieselskelett lebender Pflanzenzellen. Bot. Ztg 1861, 19, 209–215. [Google Scholar]
- Siesser, W.G. Christian Gottfried Ehrenberg: Founder of micropaleontology. Centaurus 1981, 2, 166–188. [Google Scholar] [CrossRef] [PubMed]
- Piperno, D.R. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists; Rowman & Littlefield Altamira Press: Lanham, MD, USA; New York, NY, USA; Toronto, ON, Canada; Oxford, UK, 2006; p. 238. [Google Scholar]
- Engel, W. Untersuchungen über die Kieselsäureverbindungen im Roggenhalm. Planta 1953, 41, 358–390. [Google Scholar] [CrossRef]
- Yoshida, S.; Ohnishi, Y.; Kitagishi, K. Histochemistry of silicon in rice plant: III. The presence of cuticle-silica double layer in the epidermal tissue. Soil Sci. Plant Nutr. 1962, 8, 1–5. [Google Scholar] [CrossRef]
- Kaufman, P.B.; Dayanandan, P.; Franklin, C.I.; Takeoka, Y. Structure and function of silica bodies in the epidermal system of grass shoots. Ann. Bot. 1985, 55, 487–507. [Google Scholar] [CrossRef]
- Dayanandan, P.; Kaufman, P.B.; Franklin, C.I. Detection of silica in plants. Am. J. Bot. 1983, 70, 1079–1084. [Google Scholar] [CrossRef]
- Sangster, A.G. Silicon in the roots of higher plants. Am. J. Bot. 1978, 65, 929–935. [Google Scholar] [CrossRef]
- Sangster, A.G.; Parry, D.W. Ultrastructure of silica deposits in higher plants. In Silicon and Siliceous Structures in Biological Systems; Simpson, T.L., Volcani, B.E., Eds.; Springer: New York, NY, USA, 1981; pp. 383–407. [Google Scholar]
- Parry, D.W.; Hodson, M.J.; Sangster, A.G. Some recent advances in studies of silicon in higher plants. Philos. Trallsactions R. Soc. Lond. Ser. B 1984, 304, 537–549. [Google Scholar] [CrossRef]
- Montgomery, D.J.; Parry, D.W. The ultrastructure and analytical microscopy of silicon deposition in the intercellular spaces of the roots of Molinia caerulea (L.) Moench. Ann. Bot. 1979, 44, 79–84. [Google Scholar] [CrossRef]
- Sangster, A.G.; Parry, D.W. Endodermal Silicification in Mature, Nodal Roots of Sorghum bicolor (L.) Moench. Ann. Bot. 1976, 40, 373–379. [Google Scholar] [CrossRef]
- Sangster, A.G.; Parry, D.W. Endodermal Silicon Deposits and Their Linear Distribution in Developing Roots of Sorghum bicolor (L.) Moench. Ann. Bot. 1976, 40, 361–371. [Google Scholar] [CrossRef]
- Sangster, A.G.; Parry, D.W. The Ultrastructure and Electron-probe Microassay of Silicon Deposits in the Endodermis of the Seminal Roots of Sorghum bicolor (L.) Moench. Ann. Bot. 1976, 40, 447–459. [Google Scholar] [CrossRef]
- Bennett, D.M. Silicon deposition in the roots of Hordeum sativum Jess, Avena sativa L. and Triticum aestivum L. Ann. Bot. 1982, 50, 239–245. [Google Scholar] [CrossRef]
- Soukup, M.; Martinka, M.; Bosnic, D.; Čaplovičová, M.; Elbaum, R.; Lux, A. Formation of silica aggregates in sorghum root endodermis is pre-determined by cell wall architecture and development. Ann. Bot. 2017, 120, 739–753. [Google Scholar] [CrossRef] [Green Version]
- Parry, D.W.; Soni, S.L. Electron-probe microanalysis of silicon in the roots of Oryza sativa L. Ann. Bot. 1972, 36, 781–783. [Google Scholar] [CrossRef]
- Hodson, M.J.; Sangster, A.G. X-ray microanalysis of the seminal root of Sorghum bicolor with particular reference to silicon. Ann. Bot. 1989, 64, 659–667. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, C.; Wang, H.; Zhang, F. Effect of Si on the distribution of Cd in rice seedlings. Plant Soil 2005, 272, 53–60. [Google Scholar] [CrossRef]
- Gong, H.J.; Randall, D.P.; Flowers, T.J. Silicon deposition in the root reduces sodium uptake in rice (Oryza sativa L.) seedlings by reducing bypass flow. Plant Cell Environ. 2006, 29, 1970–1979. [Google Scholar] [CrossRef]
- Hodson, M.J.; Sangster, A.G. Subcellular localization of mineral deposits in the roots of wheat (Triticum aestivum L.). Protoplasma 1989, 15, 19–32. [Google Scholar] [CrossRef]
- Moore, K.L.; Schroder, M.; Wu, Z.; Martin, B.G.; Hawes, C.R.; McGrath, S.P.; Hawkesford, M.J.; Ma, J.F.; Zhao, F.J.; Grovenor, C.R. High-resolution secondary ion mass spectrometry reveals the contrasting subcellular distribution of arsenic and silicon in rice roots. Plant Physiol. 2011, 156, 913–924. [Google Scholar] [CrossRef] [Green Version]
- Fleck, A.T.; Schulze, S.; Hinrichs, M.; Specht, A.; Waßmann, F.; Schreiber, L.; Schenk, M.K. Silicon promotes exodermal Casparian band formation in Si-accumulating and Si-excluding species by forming phenol complexes. PLoS ONE 2015, 10, e0138555. [Google Scholar] [CrossRef]
- Enstone, D.E.; Peterson, C.A.; Ma, F. Root endodermis and exodermis: Structure, function, and responses to the environment. J. Plant Growth Regul. 2003, 21, 335–351. [Google Scholar] [CrossRef]
- Lukačová, Z.; Švubová, R.; Kohanová, J.; Lux, A. Silicon mitigates the Cd toxicity in maize in relation to cadmium translocation, cell distribution, antioxidant enzymes stimulation and enhanced endodermal apoplasmic barrier development. Plant Growth Regul. 2013, 70, 89. [Google Scholar] [CrossRef]
- Vaculíková, M.; Vaculík, M.; Tandy, S.; Luxová, M.; Schulin, R. Alleviation of antimonate (SbV) toxicity in maize by silicon (Si). Environ. Exp. Bot. 2016, 128, 11–17. [Google Scholar] [CrossRef]
- Tomlinson, P.B. An anatomical approach to the classification of the Musaceae. J. Linn. Soc. Bot. 1959, 55, 779–809. [Google Scholar] [CrossRef]
- Tomlinson, P.B. Anatomy of the Monocotyledons, II. Palmae; Oxford at the Clarendon Press: London, UK, 1961; p. 477. [Google Scholar]
- Bokor, B.; Soukup, M.; Vaculík, M.; Vďačný, P.; Weidinger, M.; Lichtscheidl, I.; Vávrová, S.; Šoltys, K.; Sonah, H.; Deshmukh, R.; et al. Silicon uptake and localisation in Date palm (Phoenix dactylifera)—A unique association with sclerenchyma. Front. Plant Sci. 2019, 10, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtzmeier, M.A.; Stern, W.L.; Judd, W.S. Comparative anatomy and systematics of Senghas’s cushion species of Maxillaria (Orchidaceae). Bot. J. Linn. Soc. 1998, 127, 43–82. [Google Scholar] [CrossRef]
- Prychid, C.J.; Rudall, P.J.; Gregory, M. Systematics and biology of silica bodies in monocotyledons. Bot. Rev. 2003, 69, 377–440. [Google Scholar] [CrossRef]
- Hodson, M.J.; Wilkins, D.A. Localization of aluminium in the roots of Norway spruce (Picea abies (L.) Karst.) inoculated with Paxillus involutus Fr. New Phytol. 1991, 118, 273–278. [Google Scholar] [CrossRef]
- Sangster, A.G.; Hodson, M.J. Silica deposition in subterranean organs. In Phytolith Systematics, Advances in Archaeological and Museum Science; Rapp, G., Mulholland, S.C., Eds.; Springer: Boston, MA, USA, 1992; Volume 1, pp. 239–251. [Google Scholar]
- Watteau, F.; Villemin, G. Ultrastructural study of the biogeochemical cycle of silicon in the soil and litter of a temperate forest. Eur. J. Soil Sci. 2001, 52, 385–396. [Google Scholar] [CrossRef]
- Turpault, M.-P.; Calvaruso, C.; Kirchen, G.; Redon, P.-O.; Cochet, C. Contribution of fine tree roots to the silicon cycle in a temperate forest ecosystem developed on three soil types. Biogeosciences 2018, 15, 2231–2249. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, P.; Tripathi, R.D.; Singh, R.P.; Dwivedi, S.; Goutam, D.; Shri, M.; Trivedi, P.K.; Chakrabarty, D. Silicon mediates arsenic tolerance in rice (Oryza sativa L.) through lowering of arsenic uptake and improved antioxidant defence system. Ecol. Eng. 2013, 52, 96–103. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Hodson, M.; White, P.; Mead, A.; Broadley, M. Phylogenetic variation in the silicon composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lux, A.; Rost, T.L. Plant root research: The past, the present and the future. Ann. Bot. 2012, 110, 201–204. [Google Scholar] [CrossRef] [Green Version]
- Currie, H.A.; Perry, C.C. Chemical evidence for intrinsic ‘Si’ within Equisetum cell walls. Phytochemistry 2009, 70, 2089–2095. [Google Scholar] [CrossRef]
- Salmén, L. Wood morphology and properties from molecular perspectives. Ann. For. Sci. 2015, 72, 679–684. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Soukup, M.; Elbaum, R. Silicification in grasses: Variation between different cell types. Front. Plant Sci. 2017, 8, 438. [Google Scholar] [CrossRef] [Green Version]
- Soukup, M.; Zancajo, V.M.R.; Kneipp, J.; Elbaum, R. Formation of root silica aggregates in sorghum is an active process of the endodermis. J. Exp. Bot. 2019. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.Y.; Ma, X.L. In vitro simulation studies of silica deposition induced by lignin from rice. J. Zhejiang Univ. Sci. B 2006, 7, 267–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dove, P.M.; Han, N.; Wallace, A.F. Systematic dependence of kinetic and thermodynamic barriers to homogeneous silica nucleation on NaCl and amino acids. J. Mater. Res. 2019, 34, 442–455. [Google Scholar] [CrossRef]
- Kumar, S.; Milstein, Y.; Brami, Y.; Elbaum, M.; Elbaum, R. Mechanism of silica deposition in sorghum silica cells. New Phytol. 2017, 213, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.; Williams, R.; Fry, S.C. Cell wall biosynthesis during silicification of grass hairs. J. Plant Physiol. 1987, 126, 437–448. [Google Scholar] [CrossRef]
- He, C.; Ma, J.; Wang, L. A hemicellulose-bound form of silicon with potential to improve the mechanical properties and regeneration of the cell wall of rice. New Phytol. 2015, 206, 1051–1062. [Google Scholar] [CrossRef]
- Guerriero, G.; Hausman, J.F.; Legay, S. Silicon and the plant extracellular matrix. Front. Plant Sci. 2016, 7, 463. [Google Scholar] [CrossRef] [Green Version]
- Kulich, I.; Vojtíková, Z.; Sabol, P.; Ortmannová, J.; Neděla, V.; Tihlaříková, E.; Žárský, V. Exocyst subunit EXO70H4 has a specific role in callose synthase secretion and silica accumulation. Plant Physiol. 2018, 176, 2040–2051. [Google Scholar] [CrossRef] [Green Version]
- Guerriero, G.; Stokes, I.; Exley, C. Is callose required for silicification in plants? Biol. Lett. 2018, 14, 20180338. [Google Scholar] [CrossRef] [Green Version]
- Law, C.; Exley, C. New insight into silica deposition in horsetail (Equisetum arvense). BMC Plant Biol. 2011, 11, 112. [Google Scholar] [CrossRef] [Green Version]
- Perry, C.C.; Lu, Y. Preparation of silicas from silicon complexes: Role of cellulose in polymerisation and aggregation control. J. Chem. Soc. Faraday Trans. 1992, 88, 2915–2921. [Google Scholar] [CrossRef]
- Kido, N.; Yokoyama, R.; Yamamoto, T.; Furukawa, J.; Iwai, H.; Satoh, S.; Nishitani, K. The matrix polysaccharide (1;3, 1;4)-beta-d-glucan is involved in silicon-dependent strengthening of rice cell wall. Plant Cell Physiol. 2015, 56, 268–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zou, W.; Li, Y.; Feng, Y.; Zhang, H.; Wu, Z.; Tu, Y.; Wang, Y.; Cai, X.; Peng, L. Silica distinctively affects cell wall features and lignocellulosic saccharification with large enhancement on biomass production in rice. Plant Sci. 2015, 239, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Sangster, A.G.; Parry, D.W. Silicon deposition in the grass leaf in relation to transpiration and the effect of dinitrophenol. Ann. Bot. 1971, 35, 667–677. [Google Scholar] [CrossRef]
- Euliss, K.W.; Dorsey, B.L.; Benke, K.C.; Banks, M.K.; Schwab, A.P. The use of plant tissue silica content for estimating transpiration. Ecol. Eng. 2005, 25, 343–348. [Google Scholar] [CrossRef]
- Exley, C. A possible mechanism of biological silicification in plants. Front. Plant. Sci. 2015, 6, 853. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, P.B.; Bigelow, W.C.; Petering, L.B.; Drogosz, F.B. Silica in developing epidermal cells of avena internodes: Electron microprobe analysis. Science 1969, 166, 1015–1017. [Google Scholar] [CrossRef]
- Sangster, A.G.; Hodson, M.J.; Parry, D.W.; Rees, J.A. A developmental study of silicification in the trichomes and associated epidermal structures of the inflorescence bracts of the grass, Phalaris canariensis L. Ann. Bot. 1983, 52, 171–187. [Google Scholar] [CrossRef]
- He, H.; Veneklaas, E.J.; Kuo, J.; Lambers, H. Physiological and ecological significance of biomineralization in plants. Trends Plant Sci. 2014, 19, 166–174. [Google Scholar] [CrossRef]
- Bélanger, R.; Deshmukh, R.; Belzile, F.; Labbé, C.; Perumal, A.; Edwards, S.M. Plant with Increased Silicon Uptake. Patent No. WO/2016/183684, 24 November 2016. [Google Scholar]
- Takahashi, E.; Ma, J.F.; Miyake, Y. The possibility of silicon as an essential element for higher plants. Comments Agric. Food Chem. 1990, 2, 99–122. [Google Scholar]
- Mitani, N.; Ma, J.F. Uptake system of silicon in different plant species. J. Exp. Bot. 2005, 56, 1255–1261. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F.; Mitani, N.; Nagao, S.; Konishi, S.; Tamai, K.; Iwashita, T.; Yano, M. Characterization of the silicon uptake and molecular mapping of the silicon transporter gene in rice. Plant Physiol. 2004, 136, 3284–3289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, N.; Mitatni, N.; Ma, J.F. A transporter regulating silicon distribution in rice shoots. Plant Cell 2008, 20, 1381–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, I.S.; Roberts, D.M. Distinct transport selectivity of two structural subclasses of the nodulin-like intrinsic protein family of plant aquaglyceroporin channels. Biochemistry 2005, 44, 16826–16834. [Google Scholar] [CrossRef] [PubMed]
- Mitani-Ueno, N.; Yamaji, N.; Ma, J.F. Silicon efflux transporters isolated from two pumpkin cultivars contrasting in Si uptake. Plant Signal. Behav. 2011, 6, 991–994. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, R.K.; Vivancos, J.; Ramakrishnan, G.; Guérin, V.; Carpentier, G.; Sonah, H.; Labbé, C.; Isenring, P.; Belzile, F.J.; Bélanger, R.R. A precise spacing between the NPA domains of aquaporins is essential for silicon permeability in plants. Plant J. 2015, 83, 489–500. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. A cooperative system of silicon transport in plants. Trends Plant Sci. 2015, 20, 435–442. [Google Scholar] [CrossRef]
- Jung, J.S.; Preston, G.M.; Smith, B.L.; Guggino, W.B.; Agre, P. Molecular structure of the water channel through aquaporin CHIP. The hourglass model. J. Biol. Chem. 1994, 269, 14648–14654. [Google Scholar]
- Forrest, K.L.; Bhave, M. Major intrinsic proteins (MIPs) in plants: A complex gene family with major impacts on plant phenotype. Funct. Integr. Genom. 2007, 7, 263–289. [Google Scholar] [CrossRef]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Bélanger, R.R. The controversies of silicon’s role in plant biology. New Phytol. 2018, 221, 67–85. [Google Scholar] [CrossRef]
- Exley, C.; Guerriero, G.; Lopez, X. How is silicic acid transported in plants? Silicon 2020. [Google Scholar] [CrossRef] [Green Version]
- Mitani, N.; Chiba, Y.; Yamaji, N.; Ma, J.F. Identification and characterization of maize and barley Lsi2-like silicon efflux transporters reveals a distinct silicon uptake system from that in rice. Plant Cell 2009, 21, 2133–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, N.; Sakurai, G.; Mitani-Ueno, N.; Ma, J.F. Orchestration of three transporters and distinct vascular structures in node for intervascular transfer of silicon in rice. Proc. Natl. Acad. Sci. USA 2015, 112, 11401–11406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, N.; Ma, J.F. Spatial distribution and temporal variation of the rice silicon transporter Lsi1. Plant Physiol. 2007, 143, 1306–1313. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F. Silicon Transporters in Higher Plants. In MIPs and Their Role in the Exchange of Metalloids. Advances in Experimental Medicine and Biology; Jahn, T.P., Bienert, G.P., Eds.; Springer: New York, NY, USA, 2010; Volume 679, pp. 99–109. [Google Scholar]
- Mitani, N.; Yamaji, N.; Ma, J.F. Identification of maize silicon influx transporters. Plant Cell Physiol. 2009, 50, 5–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiba, Y.; Mitani, N.; Yamaji, N.; Ma, J.F. HvLsi1 is a silicon influx transporter in barley. Plant J. 2009, 57, 810–818. [Google Scholar] [CrossRef]
- Montpetit, J.; Vivancos, J.; Mitani-Ueno, N.; Yamaji, N.; Rémus-Borel, W.; Belzile, F.; Ma, J.F.; Bélanger, R.R. Cloning, functional characterization and heterologous expression of TaLsi1, a wheat silicon transporter gene. Plant Mol. Biol. 2012, 79, 35–46. [Google Scholar] [CrossRef]
- Bokor, B.; Ondoš, S.; Vaculík, M.; Bokorová, S.; Weidinger, M.; Lichtscheidl, I.; Turňa, J.; Lux, A. Expression of genes for Si uptake, accumulation, and correlation of Si with other elements in ionome of maize kernel. Front. Plant Sci. 2017, 8, 1063. [Google Scholar] [CrossRef] [Green Version]
- Mitani, N.; Yamaji, N.; Ago, Y.; Iwasaki, K.; Ma, J.F. Isolation and functional characterization of an influx silicon transporter in two pumpkin cultivars contrasting in silicon accumulation. Plant J. 2011, 66, 231–240. [Google Scholar] [CrossRef]
- Yamaji, N.; Chiba, Y.; Mitani-Ueno, N.; Ma, J.F. Functional characterization of a silicon transporter gene implicated in silicon distribution in barley. Plant Physiol. 2012, 160, 1491–1497. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, R.K.; Vivancos, J.; Guérin, V.; Sonah, H.; Labbé, C.; Belzile, F.; Bélanger, R.R. Identification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Guo, J.; Duan, Y.; Zhang, T.; Huo, H.; Gong, H. Isolation and functional characterization of CsLsi1, a silicon transporter gene in Cucumis sativus. Physiol Plant 2017, 159, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Zellner, W.; Lutz, L.; Khandekar, S.; Leisner, S. Identification of NtNIP2;1: An Lsi1 silicon transporter in Nicotiana tabacum. J. Plant Nutr. 2019, 42, 1028–1035. [Google Scholar] [CrossRef]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Shivaraj, S.M.; Frenette-Cotton, R.; Tremblay, L.; Isenring, P.; Bélanger, R.R. Si permeability of a deficient Lsi1 aquaporin in tobacco can be enhanced through a conserved residue substitution. Plant Direct 2019, 3, e00163. [Google Scholar] [CrossRef] [PubMed]
- Grégoire, C.; Rémus-Borel, W.; Vivancos, J.; Labbé, C.; Belzile, F.; Bélanger, R.R. Discovery of a multigene family of aquaporin silicon transporters in the primitive plant Equisetum arvense. Plant J. 2012, 72, 320–330. [Google Scholar] [CrossRef]
- Líška, D.; Soukup, M.; Lukačová, Z.; Bokor, B.; Vaculík, M. Mechanisms of silicon-mediated alleviation of abiotic stress in plants: Recent advances and future perspective. In Silicon in Plants: Advances and Future Prospects; Tripathi, D.K., Singh, V.P., Ahmad, P., Chauhan, D.K., Prasad, S.M., Eds.; CRC Press, Taylor & Francis: Boca Raton, FL, USA, 2017; pp. 1–28. [Google Scholar]
- Vaculík, M.; Vaculíková, M. Role of silicon under heavy metal and toxic element stress: An emphasis on root biology. In Silicon in Plants: Advances and Future Prospects; Tripathi, D.K., Singh, V.P., Ahmad, P., Chauhan, D.K., Prasad, S.M., Eds.; CRC Press, Taylor & Francis: Boca Raton, FL, USA, 2017; pp. 175–194. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press Ltd.: London, UK; San Diego, CA, USA, 1986; p. 461. [Google Scholar]
- Hattori, T.; Inanaga, S.; Tanimoto, E.; Lux, A.; Luxová, M.; Sugimoto, Y. Silicon-induced changes in viscoelastic properties of sorghum root cell walls. Plant Cell Physiol. 2003, 44, 743–749. [Google Scholar] [CrossRef] [Green Version]
- Hattori, T.; Inanaga, S.; Araki, H.; An, P.; Morita, S.; Luxová, M.; Lux, A. Application of silicon enhanced drought tolerance in Sorghum bicolor. Physiol. Plant 2005, 123, 459–466. [Google Scholar] [CrossRef]
- Vaculík, M.; Lux, A.; Luxová, M.; Tanimoto, E.; Lichtscheidl, I. Silicon mitigates cadmium inhibitory effects in young maize plants. Environ. Exp. Bot. 2009, 67, 52–58. [Google Scholar] [CrossRef]
- Hodson, M.J.; Sangster, A.G. The interaction between silicon and aluminium in Sorghum bicolor (L.) Moench: Growth analysis and X-ray microanalysis. Ann. Bot. 1993, 72, 389–400. [Google Scholar] [CrossRef]
- Kopittke, P.M.; Gianoncelli, A.; Kourousias, G.; Green, K.; McKenna, B.A. Alleviation of Al Toxicity by Si Is Associated with the Formation of Al–Si Complexes in Root Tissues of Sorghum. Front. Plant Sci. 2017, 8, 2189. [Google Scholar] [CrossRef] [Green Version]
- Hodson, M.J.; Sangster, A.G. Silicon and Abiotic Stress. In Proceedings of the Second Silicon in Agriculture Conference, Tsuruoka, Yamagata, Japan, 22–26 August 2002; pp. 99–104. [Google Scholar]
- Seregin, I.V.; Ivanov, V.B. Physiological aspects of cadmium and lead toxic effects on higher plants. Russ. J. Plant Physiol. 2001, 48, 523–544. [Google Scholar] [CrossRef]
- Da Cunha, K.P.V.; do Nascimento, C.W.A. Silicon effects on metal tolerance and structural changes in maize (Zea mays L.) gown on a cadmium and zinc enriched soil. Water Air Soil Pollut. 2009, 197, 323–330. [Google Scholar] [CrossRef]
- Gu, H.H.; Zhan, S.; Wang, S.H.; Tang, Y.T.; Chaney, R.L.; Fang, X.H.; Cai, X.D.; Qiu, R.L. Silicon-mediated amelioration of zinc toxicity in rice. Plant Soil 2012, 350, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Yan, C.; Liu, J.; Lu, H.; Liu, T.; Song, Z. Effects of silicon on the distribution of cadmium compartmentation in root tips of Kandelia obovata (S.; L.) Young. Environ. Pollut. 2012, 162, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Vaculík, M.; Landberg, T.; Greger, M.; Luxová, M.; Stoláriková, M.; Lux, A. Silicon modifies root anatomy, and uptake and subcellular distribution of cadmium in young maize plants. Ann. Bot. 2012, 110, 433–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocker, K.M.; Evans, D.E.; Hodson, M.J. The amelioration of aluminium toxicity by silicon in higher plants: Solution chemistry or an in planta mechanism? Physiol. Plant 1998, 104, 608–614. [Google Scholar] [CrossRef]
- Ma, J.; Cai, H.; He, C.; Zhang, W.; Wang, L. A hemicelluloses-bound form of silicon inhibits cadmium ion uptake in rice (Oryza sativa) cells. New Phytol. 2015, 206, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of silicon in mitigation of heavy metal stresses in crop plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, L.; Nie, Q.; Zhang, W.; Zhang, F. Long-term effects of exogenous silicon on cadmium translocation and toxicity in rice (Oryza sativa L.). Environ. Exp. Bot. 2008, 62, 300–307. [Google Scholar] [CrossRef]
- Lux, A.; Luxová, M.; Morita, S.; Abe, J.; Inanaga, S. Endodermal silicification in developing seminal roots of lowland and upland cultivars in rice (Oryza sativa L.). Can. J. Bot. 1999, 77, 955–960. [Google Scholar]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon improves water use efficiency in maize plants. J. Plant Nutr. 2004, 27, 1457–1470. [Google Scholar] [CrossRef]
- Choi, E.; Sukweenadhi, J.; Kim, Y.; Jung, K.H.; Koh, S.; Hoang, V.; Yang, D. The effects of rice dressing with Paenibacillus yonginensis and silicon on crop development on South Korea’s reclaimed tidal land. Field Crops Res. 2016, 188, 121–132. [Google Scholar] [CrossRef]
- Ullah, U.; Ashraf, M.; Shahzed, S.M.; Siddiqui, A.R.; Piracha, M.A.; Suleman, M. Growth behavior of tomato (Solanum lycopersicum L.) under drought stress in the presence of silicon and plant growth promoting rhizobacteria. Soil Environ. 2016, 35, 65–75. [Google Scholar]
- Diogo, R.V.C.; Wydra, K. Silicon-induced basal resistance in tomato against Ralstonia solanacearum is related to modification of pectic cell wall polysaccharide structure. Physiol. Mol. Plant Pathol. 2007, 70, 120–129. [Google Scholar] [CrossRef]
- Chen, Y.T.; Liu, M.; Wang, L.; Lin, W.P.; Fan, X.Y.; Cai, K.Z. Proteomic characterization of silicon-mediated resistance against Ralstonia solanacearum in tomato. Plant Soil 2015, 387, 425–440. [Google Scholar] [CrossRef]
- Jiang, N.H.; Fan, X.Y.; Lin, W.P.; Wang, G.P.; Cai, K.Z. Transcriptome analysis reveals new insights into the bacterial wilt resistance mechanism mediated by silicon in tomato. Int. J. Mol. Sci. 2019, 20, 761. [Google Scholar] [CrossRef] [Green Version]
- Cherif, M.; Belanger, R.R. Use of potassium silicate amendments in recirculating nutrient solutions to suppress Pythium ultimum on long English cucumber. Plant Dis. 1992, 76, 1008–1011. [Google Scholar] [CrossRef]
- Cherif, M.; Asselin, A.; Belanger, R.R. Defense responses induced by soluble silicon in cucumber roots infected by Pythium spp. Phytopathology 1994, 84, 236–242. [Google Scholar] [CrossRef]
- Bathoova, M.; Bokor, B.; Soukup, M.; Lux, A.; Martinka, M. Silicon-mediated cell wall modifications of sorghum root exodermis and suppression of invasion by fungus Alternaria alternata. Plant Pathol. 2018, 67, 1891–1900. [Google Scholar] [CrossRef]
- de Souza, E.A.; de Moraes, J.C.; do Amaral, J.L.; Liberato, R.D.; Bonelli, E.A.; Lima, L.R. Application effect of calcium silicate in Brachiaria brizantha cv. Marandu on the nymph population of the brown root stinkbug, soil chemical characteristics, and plant and dry matter production. Cienc. Agrotec. 2009, 33, 1518–1526. [Google Scholar]
- Keeping, M.G.; Kvedaras, O.L.; Bruton, A.G. Epidermal silicon in sugarcane: Cultivar differences and role in resistance to sugarcane borer Eldana saccharina. Environ. Exp. Bot. 2009, 66, 54–60. [Google Scholar] [CrossRef]
- Silva, R.V.; Oliveira, R.D.L.; Nascimento, K.J.T.; Rodrigues, F.A. Biochemical responses of coffee resistance against Meloidogyne exigua mediated by silicon. Plant Pathol. 2010, 59, 586–593. [Google Scholar] [CrossRef]
- Zhan, L.P.; Peng, D.L.; Wang, X.L.; Kong, L.A.; Peng, H.; Liu, S.M.; Liu, Y.; Huang, W.K. Priming effect of root-applied silicon on the enhancement of induced resistance to the root-knot nematode Meloidogyne graminicola in rice. BMC Plant Biol. 2018, 18, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labrousse, P.; Delmail, D.; Arnaud, M.; Thalouarn, P. Mineral nutrient concentration influences sunflower infection by broomrape (Orobanche cumana). Botany 2010, 88, 839–849. [Google Scholar] [CrossRef]
- Karimmojeni, H.; Ehtemam, M.H.; Javadimoghadam, S.; Shahbazi, S.; Bazrafshan, A.H. Egyptian broomrape (Phelipanche aegyptiaca) response to silicon nutrition in tomato (Solanum lycopersicum L.). Arch. Agron. Soil Sci. 2017, 63, 612–618. [Google Scholar] [CrossRef]
- Bu, R.; Xie, J.M.; Yu, J.H.; Liao, W.B.; Xiao, X.M.; Lv, J.; Wang, C.L.; Ye, J.; Calderon-Urrea, A. Autotoxicity in cucumber (Cucumis sativus L.) seedlings is alleviated by silicon through an increase in the activity of antioxidant enzymes and by mitigating lipid peroxidation. J. Plant Biol. 2016, 59, 247–259. [Google Scholar] [CrossRef]
- Anda, C.C.O.; Opfergelt, S.; Declerck, S. Silicon acquisition by bananas (cv. Grande Naine) is increased in presence of the arbuscular mycorrhizal fungus Rhizophagus irregularis MUCL 41833. Plant Soil 2016, 409, 77–85. [Google Scholar] [CrossRef]
- Hajiboland, R.; Moradtalab, N.; Aliasgharzad, N.; Eshaghi, Z.; Feizy, J. Silicon influences growth and mycorrhizal responsiveness in strawberry plants. Physiol. Molecul. Biol. Plants 2018, 24, 1103–1115. [Google Scholar] [CrossRef]
- Moradtalab, N.; Hajiboland, R.; Aliasgharzad, N.; Hartmann, T.E.; Neumann, G. Silicon and the association with an arbuscular-mycorrhizal fungus (Rhizophagus clarus) mitigate the adverse effects of drought stress on strawberry. Agronomy 2019, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Dannon, E.A.; Wydra, K. Interaction between silicon amendment, bacterial wilt development and phenotype of Ralstonia solanacearum in tomato genotypes. Physiol. Mol. Plant Pathol. 2004, 64, 233–243. [Google Scholar] [CrossRef]
- Heine, G.; Tikum, G.; Horst, W.J. The effect of silicon on the infection by and spread of Pythium aphanidermatum in single roots of tomato and bitter gourd. J. Exp. Bot. 2007, 58, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Vermeire, M.; Kablan, L.A.; Dorel, M.; Delvaux, B.; Risède, J.; Legrève, A. Protective role of silicon in the banana-Cylindrocladium spathiphylli pathosystem. Eur. J. Plant Pathol. 2011, 131, 621–630. [Google Scholar] [CrossRef]
- Fortunato, A.A.; Rodrigues, F.Á.; Baroni, J.C.; Soares, G.C.; Rodriguez, M.A.; Pereira, O.L. Silicon suppresses Fusarium wilt development in banana plants. J. Phytopathol. 2012, 160, 674–679. [Google Scholar] [CrossRef]
- Wang, L.; Cai, K.; Chen, Y.; Wang, G. Silicon-mediated tomato resistance against Ralstonia solanacearum is associated with modification of soil microbial community structure and activity. Biol. Trace Elem. Res. 2013, 152, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Mattei, D.; Dias-Arieira, C.R.; Lopes, A.P.M.; Miamoto, A. Influence of Rocksil (R), Silifort (R) and wollastonite on penetration and development of Meloidogyne javanica in Poaceae and Fabaceae. J. Phytopathol. 2017, 165, 91–97. [Google Scholar] [CrossRef]
- Gopal, M.; Gupta, A.; Nair, R. Variations in hosting beneficial plant-associated microorganisms by root (wilt)-diseased and field-tolerant coconut palms of West Coast tall variety. Curr. Sci. 2005, 59, 1922–1927. [Google Scholar]
- Cogliati, E.; Gilardi, G.; Garibaldi, A.; Gullino, M. Effects of increased conductivity of the nutrient solutions and of the addition of potassium silicate against Alternaria japonica on rocket (Eruca vesicaria) in soilless cultivation. J. Plant Pathol. 2013, 95, 25–31. [Google Scholar]
- Khoshgoftarmanesh, A.H.; Mohaghegh, P.; Sharifnabi, B.; Shirvani, M.; Khalili, B. Silicon nutrition and Phytophthora drechsleri infection effects on growth and mineral nutrients concentration, uptake, and relative translocation in hydroponic-grown cucumber. J. Plant Nutr. 2012, 35, 1168–1179. [Google Scholar] [CrossRef]
- Mohaghegh, P.; Khoshgoftarmanesh, A.H.; Shirvani, M.; Sharifnabi, B.; Nili, N. Effect of silicon nutrition on oxidative stress induced by Phytophthora melonis infection in cucumber. Plant Dis. 2011, 95, 455–460. [Google Scholar] [CrossRef]
- Huang, C.H.; Roberts, P.D.; Datnoff, L.E. Silicon suppresses Fusarium crown and root rot of tomato. J. Phytopathol. 2011, 159, 546–554. [Google Scholar] [CrossRef]
- Fortunato, A.A.; Rodrigues, F.Á.; Nascimento, K.J.T.D. Physiological and biochemical aspects of the resistance of banana plants to Fusarium wilt potentiated by silicon. Phytopathology 2012, 102, 957–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gent, M.N.P.; Elmer, W.H. Partial saturation ebb and flow irrigation and silicon to suppress Pythium root rot of poinsettia. Crop Prot. 2017, 101, 95–102. [Google Scholar] [CrossRef]
- Roldi, M.; Dias-Arieira, C.R.; da Silva, S.A.; Dorigo, O.F.; Machado, A.C. Control of Meloidogyne paranaensis in coffee plants mediated by silicon. Nematology 2017, 19, 245–250. [Google Scholar] [CrossRef]
- Oliveira, R.M.; Ribeiro, R.C.F.; Xavier, A.A.; Pimenta, L.; Korndorfer, G.H. Effect of calcium and magnesium silicate on Meloidogyne javanica reproduction and development of Banana Prata-A seedlings. Rev. Bras. Frutic. 2012, 34, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Korndorfer, A.P.; Grisoto, E.; Vendramim, J.D. Induction of insect plant resistance to the spittlebug mahanarva fimbriolata stal (Hemiptera: Cercopidae) in sugarcane by silicon application. Neotrop. Entomol. 2011, 40, 387–392. [Google Scholar]
- Frew, A.; Powell, J.R.; Hiltpold, I.; Allsopp, P.G.; Sallam, N.; Johnson, S.N. Host plant colonisation by arbuscular mycorrhizal fungi stimulates immune function whereas high root silicon concentrations diminish growth in a soil-dwelling herbivore. Soil Biol. Biochem. 2017, 112, 117–126. [Google Scholar] [CrossRef]
- Kvedaras, O.L.; Keeping, M.G.; Goebel, F.R.; Byrne, M.J. Larval performance of the pyralid borer Eldana saccharina Walker and stalk damage in sugarcane: Influence of plant silicon, cultivar and feeding site. Int. J. Pest. Manag. 2007, 53, 183–194. [Google Scholar] [CrossRef]
- Kvedaras, O.L.; Byrne, M.J.; Coombes, N.E.; Keeping, M.G. Influence of plant silicon and sugarcane cultivar on mandibular wear in the stalk borer Eldana saccharina. Agric. For. Entomol. 2009, 11, 301–306. [Google Scholar] [CrossRef]
- DoGramaci, M.; Arthurs, S.P.; Chen, J.J.; Osborne, L. Silicon applications have minimal effects on Scirtothrips dorsalis (Thysanoptera thripidae) populations on pepper plant, Capsicum annum L. Fla. Entomol. 2013, 96, 48–54. [Google Scholar] [CrossRef]
- Lukačová, Z.; Švubová, R.; Janikovičová, S.; Volajová, Z.; Lux, A. Tobacco plants (Nicotiana benthamiana) were influenced by silicon and were not infected by dodder (Cuscuta europaea). Plant Physiol. Biochem. 2019, 139, 179–190. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lux, A.; Lukačová, Z.; Vaculík, M.; Švubová, R.; Kohanová, J.; Soukup, M.; Martinka, M.; Bokor, B. Silicification of Root Tissues. Plants 2020, 9, 111. https://doi.org/10.3390/plants9010111
Lux A, Lukačová Z, Vaculík M, Švubová R, Kohanová J, Soukup M, Martinka M, Bokor B. Silicification of Root Tissues. Plants. 2020; 9(1):111. https://doi.org/10.3390/plants9010111
Chicago/Turabian StyleLux, Alexander, Zuzana Lukačová, Marek Vaculík, Renáta Švubová, Jana Kohanová, Milan Soukup, Michal Martinka, and Boris Bokor. 2020. "Silicification of Root Tissues" Plants 9, no. 1: 111. https://doi.org/10.3390/plants9010111