Blue Light added with Red LEDs Enhance Growth Characteristics, Pigments Content, and Antioxidant Capacity in Lettuce, Spinach, Kale, Basil, and Sweet Pepper in a Controlled Environment



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. LED Light Treatments
2.3. Plant Growth and Biomass Measurement
2.4. Chlorophyll and Carotenoid Analysis
2.5. Antioxidant Capacity Measurement
2.6. Statistical Analysis
3. Results and Discussion
3.1. Effect of Different Combinations of Red and Blue Light on the Plant Growth Characteristics
3.1.1. Plant Height and Growth Features
3.1.2. Leaf Number
3.1.3. Shoot Fresh and Dry Mass
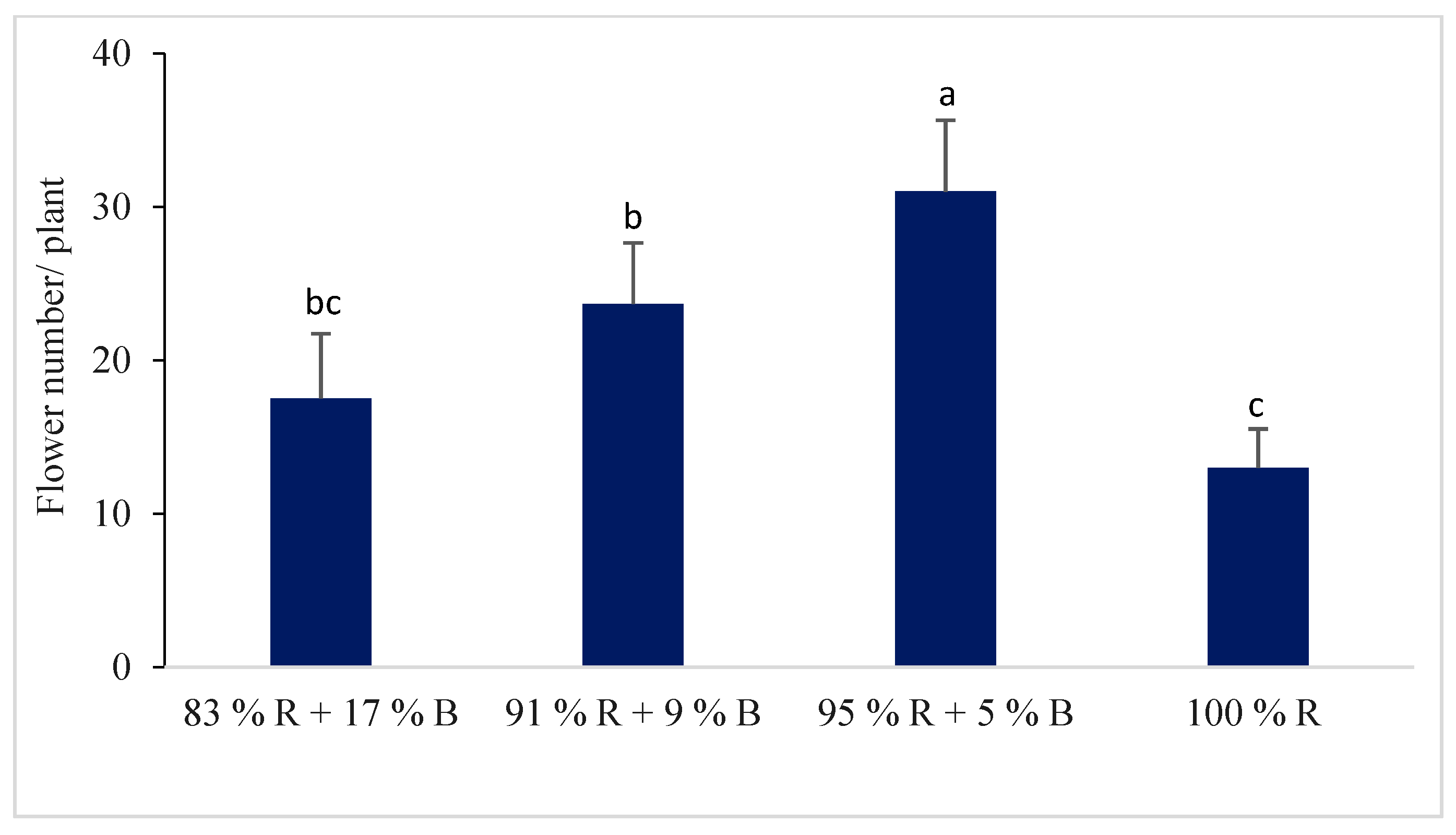
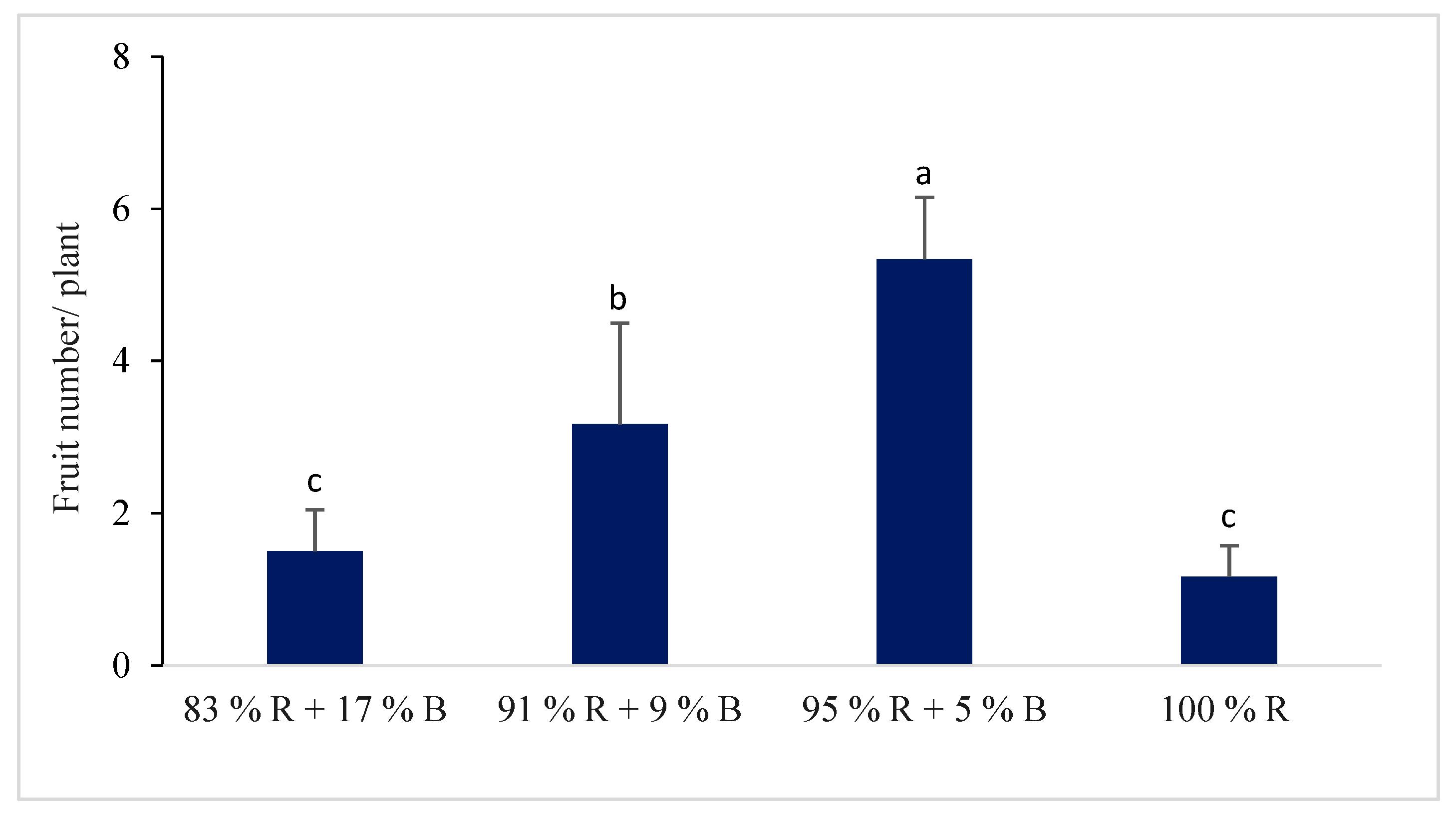
3.1.4. Flower and Fruit of the Pepper Plant
3.2. Effect of Different Combinations of Red and Blue Light on the Accumulation of Plants’ Pigment Content, and Antioxidant Capacity
3.2.1. Chlorophyll (chl) and Carotenoid Content
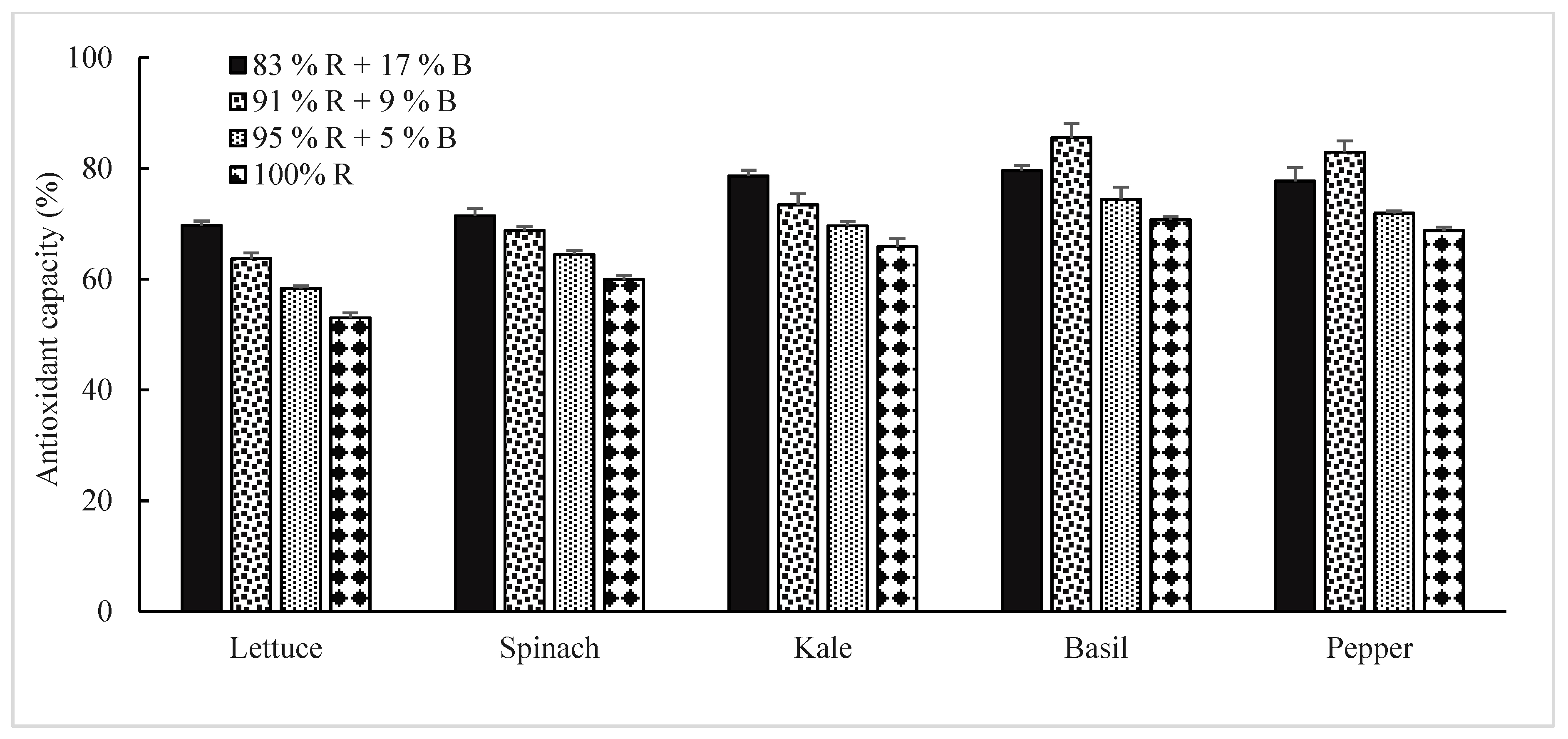
3.2.2. Antioxidant Capacity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kopsell, D.A.; Kopsell, D.E. Genetic and environmental factors affecting plant lutein/zeaxanthin. Agro Food Ind. Hi-Tech 2008, 19, 44–46. [Google Scholar]
- Hopkins, W.G.; Huner, N.P.A. Introduction to Plant Physiology, 3rd ed.; John Wiley and Sons: Hoboken, NJ, USA, 2004. [Google Scholar]
- Ramalho, J.C.; Marques, N.C.; Semedo, J.N.; Matos, M.C.; Quartin, V.L. Photosynthetic performance and pigment composition of leaves from two tropical species is determined by light quality. Plant Biol. 2002, 4, 112–120. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z.; Liu, X.; Han, X. Effects of different light sources on thegrowth of non-heading Chinese cabbage (Brassica campestris L.). J. Agric. Sci. 2012, 4, 262–270. [Google Scholar]
- Olle, M.; Virsilė, A. The effects of light-emitting diode lighting on greenhouse plant growth and quality. Agric. Food Sci. 2013, 22, 223–234. [Google Scholar] [CrossRef]
- Morrow, R.C. LED lighting in horticulture. HortScience 2008, 43, 1947–1950. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Cetner, M.D.; Samborska, I.A.; Kalaji, M.H. Measuring light spectrum as a main indicator of artificial sources quality. J. Coast. Life Med. 2015, 308, 400–406. [Google Scholar]
- Yeh, N.; Chung, J.P. High-brightness LEDs- Energy efficient lighting sources and their potential in indoor plant cultivation. Renew. Sustain. Energy Rev. 2009, 13, 2175–2180. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose–responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Matsuda, R.; Ohashi-kaneko, K.; Fujiwara, K.; Kurata, K. Analysis of the relationship between blue-light photon flux density and the photosynthetic properties of spinach (Spinacia oleracea L.) leaves with regard to the acclimation of photosynthesis to growth irradiance. Soil Sci. Plant Nutr. 2007, 53, 459–465. [Google Scholar] [CrossRef]
- Savvides, A.D.; Fanourakis, D.; van Ieperen, W. Co-ordination of hydraulic and stomatal conductances across light qualities in cucumber leaves. J. Exp. Bot. 2012, 63, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Stutte, G.W.; Edney, S.; Skerritt, T. Photoregulation of bioprotectant content of red leaf lettuce with light-emitting diodes. HortScience 2009, 44, 79–82. [Google Scholar] [CrossRef]
- Heo, J.W.; Kang, D.H.; Bang, H.S.; Hong, S.G.; Chun, C.; Kang, K.K. Early growth, pigmentation, protein content, and phenylalanine ammonia-lyase activity of red curled lettuces grown under different lighting conditions. Korean J. Hortic. Sci. Technol. 2012, 30, 6–12. [Google Scholar] [CrossRef]
- Johkan, M.K.; Shoji, F.; Goto, S.; Hashida, S.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience 2010, 45, 1809–1814. [Google Scholar] [CrossRef]
- Son, K.H.; Oh, M.M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Piovene, C.; Orsini, F.; Bosi, S.; Sanoubar, R.; Bregola, V.; Dinelli, G.; Gianquinto, G. Optimal red:blue ratio in led lighting for nutraceutical indoor horticulture. Sci. Hortic. 2015, 193, 202–208. [Google Scholar] [CrossRef]
- Gangadhar, B.H.; Mishra, R.K.; Pandian, G.; Park, S.W. Comparative study of color, pungency, and biochemical composition in chili pepper (Capsicum annuum L.) under different light-emitting diode treatments. HortScience 2012, 47, 1729–1735. [Google Scholar] [CrossRef]
- Lisiewska, Z.; Kmiecik, W.; Gębczyński, P.; Sobczyńska, L. Amino acid profile of raw and as-eaten products of spinach (Spinacia oleracea L.). Food Chem. 2011, 126, 460–465. [Google Scholar] [CrossRef]
- Becker, C.; Klaering, H.-P.; Kroh, L.W.; Krumbein, A. Cool-cultivated red leaf lettuce accumulates cyanidin-3-O-(6″-O-malonyl)-glucoside and caffeoylmalic acid. Food Chem. 2014, 146, 404–411. [Google Scholar] [CrossRef]
- Schmidt, S.; Zietz, M.; Schreiner, M.; Rohn, S.; Kroh, L.W.; Krumbein, A. Genotypic and climatic influences on the concentration and composition of flavonoids in kale (Brassica oleracea var. sabellica). Food Chem. 2010, 119, 1293–1299. [Google Scholar] [CrossRef]
- Lefsrud, M.G.; Kopsell, D.A.; Sams, C.E. Irradiance from distinct wavelength lightemitting diodes affect secondary metabolites in kale. HortScience. 2008, 43, 2243–2244. [Google Scholar] [CrossRef]
- Singletary, K.W. Basil: A brief summary of potential health benefits. Nutr. Today 2018, 53, 92–97. [Google Scholar] [CrossRef]
- Jovicich, E.; VanSickle, J.J.; Cantliffe, D.J.; Stoffella, P.J. Greenhouse-grown colored peppers: A profitable alternative for vegetable production in Florida? HortTechnology 2005, 15, 355–369. [Google Scholar] [CrossRef]
- Massa, G.; Graham, T.; Haire, T.; Flemming, C., II; Newsham, G.; Wheeler, R. Light emitting diode light transmission through leaf tissue of seven different crops. HortScience 2015, 50, 501–506. [Google Scholar] [CrossRef]
- Muneer, S.; Kim, E.J.; Park, J.S.; Lee, J.H. Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca Sativa L.). Int. J. Mol. Sci. 2014, 15, 4657–4670. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Naznin, M.T.; Maeda, T.; Morita, N. Antioxidant function of E- and Z-ajoene derived from Japanese garlic. Int. J. Food Prop. 2010, 13, 821–829. [Google Scholar] [CrossRef]
- Sharma, O.P.; Bhat, T.K. Analytical methods DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Ward, J.M.; Cufr, C.M.; Denzel, M.A.; Neff, M.M. The dof transcription factor OBP3 modulates Phytochrome and cryptochrome signaling in arabidopsis. Plant Cell 2005, 17, 475–485. [Google Scholar] [CrossRef]
- Poudel, P.R.; Kataoka, I.; Mochioka, R. Effect of red- and blue-light-emitting diodes on growth and morphogenesis of grapes. Plant Cell Tissue Organ Cult. 2008, 92, 147–153. [Google Scholar] [CrossRef]
- Chatterjee, M.; Sharma, P.; Khurana, J.P. Cryptochrome 1 from Brassica napus is up-regulated by blue light and controls hypocotyl/stem growth and anthocyanin accumulation. Plant Physiol. 2006, 141, 61–74. [Google Scholar] [CrossRef]
- Brown, C.S.; Schuerger, A.C.; Sager, J.C. Growth and photomorphogenesis of pepper plants under red light-emitting diodes with supplemental blue or far-red lighting. J. Am. Soc. Hortic. Sci. 1995, 120, 808–813. [Google Scholar] [CrossRef]
- Liu, X.Y.; Chang, T.T.; Guo, S.R.; Xu, Z.G.; Li, J. Effect of different light quality of LED on growth and photosynthetic cheracter in cherry tomato seedling. Acta Hortic. 2011, 907, 325–330. [Google Scholar] [CrossRef]
- Ahmad, M.; Cashmore, A.R. The blue-light receptor cryptochrome 1 shows functional dependence on phytochrome A of ohytochrome B in Arabidopsis thaliana. Plant J. 1997, 11, 421–427. [Google Scholar] [CrossRef]
- Ahmad, M.; Grancher, N.; Heil, M.; Black, R.C.; Giovani, B.; Galland, P.; Lardemer, D. Action spectrum for cryptochrome-dependent hypocotyl growth inhibition in Arabidopsis. Plant Physiol. 2002, 129, 774–785. [Google Scholar] [CrossRef]
- Hernàndez, R.; Kubota, C. Tomato seedling growth and morphological responses to supplemental LED lighting red: Blue ratios under varied daily solar light integrals. Acta Hortic. 2012, 956, 187–194. [Google Scholar] [CrossRef]
- Wollaeger, H.M.; Runkle, E.S. Growth of impatiens, Petunia, Salvia, and tomato seedlings under blue, green, and red light-emitting diodes. HortScience 2014, 49, 734–740. [Google Scholar] [CrossRef]
- Lin, W.C.; Jolliffe, P.A. Light intensity and spectral quality affect fruit growth and shelf life of greenhouse-grown long english cucumber. J. Am. Soc. Hortic. Sci. 1996, 121, 1168–1173. [Google Scholar] [CrossRef]
- Opdam, J.G.; Schoonderbeek, G.G.; Heller, E.B.; Gelder, A. Closed green-house: A starting point for sustainable entrepreneurship in horticulture. Acta Hortic. 2005, 691, 517–524. [Google Scholar] [CrossRef]
- Trouwborst, G.; Hogewoning, S.W.; van Kooten, O.; Harbinson, J.; van Ieperen, W. Plasticity of photosynthesis after the red light syndrome’ in cucumber. Environ. Exp. Bot. 2016, 121, 75–82. [Google Scholar] [CrossRef]
- Saebo, A.; Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of brich plantlets in vitro. Plant Cell Tissue Organ Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Lee, J.G.; Oh, S.S.; Cha, S.H.; Jang, Y.A.; Kim, S.Y.; Um, Y.C.; Cheong, S.R. Effects of red/blue light ratio and short-term light quality conversion on growth and anthocyanin contents of baby leaf lettuce. J. Bio-Environ. Control 2010, 19, 351–359. [Google Scholar]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E. Increases in shoot tissue pigments, glucosinolates, and mineral elements in sprouting broccoli after exposure to short-duration blue light from light emitting diodes. J. Am. Soc. Hortic. Sci. 2013, 138, 31–37. [Google Scholar] [CrossRef]
- Briggs, W.R.; Christie, J.M. Phototropins 1 and 2: Versatile plant blue-light receptors. Trends Plant Sci. 2002, 7, 204–210. [Google Scholar] [CrossRef]
- Tanaka, M.; Takamura, T.; Watanabe, H.; Endo, M.; Yanagi, T.; Okamoto, K. In vitro growth of Cymbidium plantlets cultured under super bright red and blue light emitting diodes (LEDs). J. Hortic. Sci. Biotechnol. 1998, 73, 39–44. [Google Scholar] [CrossRef]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z. The effect of different light qualities on rape seed (Bassicanapus L.) plantlet growth and morphogenesis in vitro. Sci. Hortic. 2013, 150, 117–124. [Google Scholar] [CrossRef]
- Ma, G.; Zang, L.; Kato, M.; Yamawaki, K.; Kiriiwa, Y.; Yahata, M.; Ikoma, Y.; Matsumoto, H. Effect of blue light and red LED light irradiation on β-cryptoxanthin accumulation in the flavedo of citrus fruits. J. Agric. Food Chem. 2012, 60, 197–201. [Google Scholar] [CrossRef]
- Bukhov, N.G.; Drozdova, I.S.; Bondar, V.V.; Mokronosov, A.T. Blue, red and blueplus red light control of chlorophyll content and CO2 gas exchange in barleyleaves: Quantitative description of the effects of light quality and fluence rate. Physiol. Planta 1992, 85, 632–638. [Google Scholar] [CrossRef]
- Yu, X.; Liu, H.; Klejnot, J.; Lin, C. The cryptochrome blue light receptors. In The Arabidopsis Book; ASPB: Rockville, MD, USA, 2010. [Google Scholar]
- Wu, M.C.; Hou, C.Y.; Jiang, C.M.; Wang, C.Y.; Chen, H.H.; Chang, H.M. A novel approach of LED light radiation improves the antioxidant activity of pea seedlings. Food Chem. 2007, 101, 1753–1758. [Google Scholar] [CrossRef]
- Liu, Y.; Roof, S.; Ye, Z.; Barry, C.; van Tuinen, A.; Vrebalov, J.; Bowler, C.; Giovannoni, J. Manipulation of light signal transduction as a means of modifying fruit nutritional quality in tomato. Proc. Natl. Acad. Sci. USA 2004, 101, 9897–9902. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.Q.; Wu, Y.J.; Tang, R.H.; Liu, D.; Liu, Y.; Cashmore, A.R. The C termini of Arabidopsis cryptochromes mediate a constitutive light response. Cell 2000, 103, 815–827. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Treatments | Plant Height (cm) | Leaf No/Plant | Fresh Mass/Plant (g) | Dry Mass/Plant (g) |
|---|---|---|---|---|---|
| Lettuce | 83% R + 17% B | 19.52 ± 2.98 ab | 30.33 ± 5.54 a | 70.85 ± 5.83 ab | 10.45 ± 1.14 ab |
| 91% R + 9% B | 19.33 ± 4.23 ab | 26.67 ± 3.27 a | 78.45 ± 10.21 a | 12.28 ± 3.52 a | |
| 95% R + 5% B | 18.17 ± 4.62 b | 25.50 ± 3.01 a | 72.52 ± 7.80 ab | 9.15 ± 1.05 a | |
| 100% R | 24.33 ± 2.50 a | 24.67 ± 4.13 a | 64.82 ± 8.05 b | 6.95 ± 0.85 b | |
| LSD < 0.05 | 3.68 | 4.11 | 8.12 | 1.97 | |
| Spinach | 83% R + 17% B | 31.35 ± 3.59 a | 66.67 ± 9.71 a | 63.73 ± 6.36 ab | 9.88 ± 1.09 ab |
| 91% R + 9% B | 35.6 ± 3.32 a | 72.5 ± 8.06 a | 68.73 ± 6.87 a | 12.13 ± 2.7 a | |
| 95% R + 5% B | 29.83 ± 2.93 a | 69.17 ± 5.63 a | 65.25 ± 3.43 a | 11.35 ± 0.50 ab | |
| 100% R | 35.17 ± 6.88 a | 36.33 ± 9.09 b | 47.7 ± 12.09 b | 8.45 ± 2.9 b | |
| LSD < 0.05 | 4.47 | 8.27 | 7.84 | 2.01 | |
| Kale | 83% R + 17% B | 46.05 ± 1.79 b | 27.83 ± 2.92 a | 109.78 ± 9.99 b | 26.21 ± 3.34 a |
| 91% R + 9% B | 46.43 ± 2.82 b | 28.83 ± 2.04 a | 185.6 ± 6.56 a | 28.11 ± 4.12 a | |
| 95% R + 5% B | 47.38 ± 1.51 ab | 28.17 ± 2.14 a | 189.95 ± 7.32 a | 29.71 ± 3.24 a | |
| 100% R | 51.63 ± 4.95 a | 29.17 ± 1.94 a | 85.45 ± 9.98 c | 19.76 ± 3.67 b | |
| LSD < 0.05 | 3.08 | 2.29 | 8.61 | 3.61 | |
| Basil | 83% R + 17% B | 45.68 ± 3.99 a | 101.83 ± 2.78 ab | 41.2 ± 8.82 ab | 10.13 ± 0.73 ab |
| 91% R + 9% B | 47.33 ± 4.34 a | 107.5 ± 9.20 a | 44.23 ± 3.58 a | 13.45 ± 3.22 a | |
| 95% R + 5% B | 48.05 ± 5.76 a | 103.67 ± 7.34 a | 42.92 ± 3.22 ab | 11.95 ± 3.54 ab | |
| 100% R | 51.12 ± 4.48 a | 90.5 ± 10.42 b | 32.68 ± 9.63 b | 9.37 ± 1.19 b | |
| LSD < 0.05 | 4.69 | 7.98 | 6.96 | 2.49 | |
| Pepper | 83% R + 17% B | 467.48 ± 6.6 c | 183.39 ± 8.4 b | 650.88 ± 8.8 c | 218.37 ± 4.5 b |
| 91% R + 9% B | 492.27 ± 6.1 b | 191.61 ± 6.5 b | 683.89 ± 7.5 b | 305.49 ± 4.9 a | |
| 95% R + 5% B | 568.33 ± 6.7 a | 215.37 ± 3.6 a | 783.71 ± 6.1 a | 197.03 ± 4.5 c | |
| 100% R | 459.42 ± 4.7 c | 161.39 ± 5.0 c | 620.82 ± 7.2 d | 183.67 ± 4.9 d | |
| LSD < 0.05 | 6.1 | 6.1 | 7.5 | 4.7 |
| Plants | Treatments | chl a (µg g−1 DM) | chl b (µg g−1 DM) | Total chl (µg g−1 DM) | Carotenoids (µg g−1 DM) |
|---|---|---|---|---|---|
| Lettuce | 83% R + 17% B | 555.69 ± 6.4 b | 177.46 ± 6.2 c | 733.14 ± 8.1 b | 201.96 ± 4.8 a |
| 91% R + 9% B | 600.51 ± 5.8 a | 199.88 ± 6.8 a | 800.39 ± 9.6 a | 208.02 ± 5.6 a | |
| 95% R + 5% B | 554.67 ± 4.62 b | 188.86 ± 5.9 b | 743.53 ± 9.8 b | 181.85 ± 3.9 b | |
| 100% R | 515.89 ± 3.9 | 145.48 ± 6.1 d | 661.38 ± 3.9 c | 151.72 ± 7.5 c | |
| LSD < 0.05 | 5.3 | 6.2 | 8.2 | 5.5 | |
| Spinach | 83% R + 17% B | 566.61 ± 5.1 b | 164.46 ± 6.2 b | 730.14 ± 8.1 b | 221.96 ± 4.8 a |
| 91% R + 9% B | 589.35 ± 7.6 a | 188.88 ± 6.8 a | 777.39 ± 9.6 a | 219.96 ± 4.8 a | |
| 95% R + 5% B | 597.16 ± 7.8 a | 162.86 ± 5.9 b | 759.53 ± 9.8 b | 171.85 ± 3.9 b | |
| 100% R | 465.68 ± 6.9 c | 137.48 ± 6.1 d | 602.38 ± 3.9 c | 163.72 ± 7.5 c | |
| LSD < 0.05 | 6.9 | 6.2 | 8.2 | 5.5 | |
| Kale | 83% R + 17% B | 578.39 ± 8.3 c | 203.37 ±7.2 c | 781.77 ± 9.2 c | 293.13 ± 6.5 a |
| 91% R + 9% B | 671.45 ± 4.9 b | 226.65 ±8.0 b | 898.09 ± 7.1 b | 271.16 ± 5.4 b | |
| 95% R + 5% B | 702.74 ± 7.6 a | 265.65 ±7.1 a | 968.39 ± 8.8 a | 261.23 ± 4.9 b | |
| 100% R | 556.13 ± 4.2 d | 201.13 ±5.6 c | 757.26 ± 5.4 d | 223.28 ± 7.9 c | |
| LSD < 0.05 | 6.5 | 7.0 | 7.8 | 6.3 | |
| Basil | 83% R + 17% B | 573.27 ± 5.1 c | 190.06 ± 4.1 b | 763.34 ± 5.3 c | 316.67 ± 9.1 a |
| 91% R + 9% B | 644.42 ± 5.6 a | 215.35 ± 6.7 a | 859.77 ± 3.3 a | 295.92 ± 3.9 b | |
| 95% R + 5% B | 618.73 ± 7.1 b | 201.51 ± 10.7 b | 820.24 ± 8.3 b | 269.96 ± 6.1 c | |
| 100% R | 524.52 ± 5.4 d | 162.32 ± 5.3 c | 686.85 ± 9.3 d | 171.42 ± 8.1 d | |
| LSD < 0.05 | 5.8 | 7.1 | 6.9 | 7.3 | |
| Pepper | 83% R + 17% B | 573.27 ± 5.1 c | 190.06 ± 4.1 b | 763.34 ± 5.3 c | 316.67 ± 9.1 a |
| 91% R + 9% B | 644.42 ± 5.6 a | 215.35 ± 6.7 a | 859.77 ± 3.3 a | 295.92 ± 3.9 b | |
| 95% R + 5% B | 618.73 ± 7.1 b | 201.51 ± 10.7 b | 820.24 ± 8.3 b | 269.96 ± 6.1 c | |
| 100% R | 524.52 ± 5.4 d | 162.32 ± 5.3 c | 686.85 ± 9.3 d | 171.42 ± 8.1 d | |
| LSD < 0.05 | 5.8 | 7.1 | 6.9 | 7.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, M.O.K. Blue Light added with Red LEDs Enhance Growth Characteristics, Pigments Content, and Antioxidant Capacity in Lettuce, Spinach, Kale, Basil, and Sweet Pepper in a Controlled Environment. Plants 2019, 8, 93. https://doi.org/10.3390/plants8040093
Naznin MT, Lefsrud M, Gravel V, Azad MOK. Blue Light added with Red LEDs Enhance Growth Characteristics, Pigments Content, and Antioxidant Capacity in Lettuce, Spinach, Kale, Basil, and Sweet Pepper in a Controlled Environment. Plants. 2019; 8(4):93. https://doi.org/10.3390/plants8040093
Chicago/Turabian StyleNaznin, Most Tahera, Mark Lefsrud, Valerie Gravel, and Md Obyedul Kalam Azad. 2019. "Blue Light added with Red LEDs Enhance Growth Characteristics, Pigments Content, and Antioxidant Capacity in Lettuce, Spinach, Kale, Basil, and Sweet Pepper in a Controlled Environment" Plants 8, no. 4: 93. https://doi.org/10.3390/plants8040093