2.1. Pompia Is a Regular Diploid Citrus
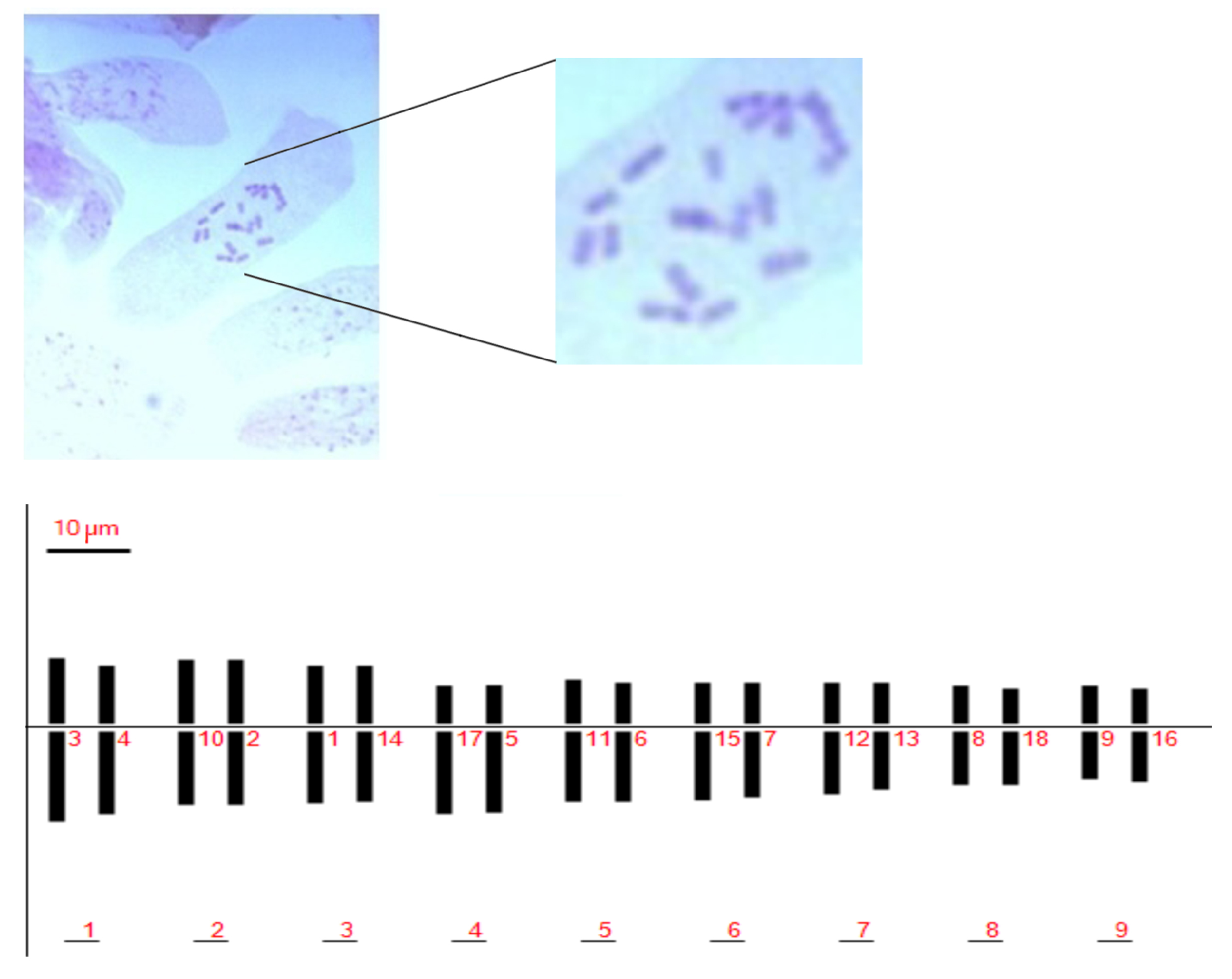
Karyo-morphometric features of Pompia’s chromosomes were visualized in squashes of colchicine treated root tips of several Pompia accessions.
No significant differences in measured karyological parameters were visualized among the analyzed accessions. Chromosome types were classified based on their arm ratio according to Levan et al. (1964) [
32]. We identified a total chromosome number of 2n = 2x = 18, sixteen of which were classified as metacentric and two as sub-metacentric (
Figure 1 and
Table 1). The Pompia karyogram, as reconstructed by the Karyotype software [
33], is shown in
Figure 1.
The ratio between the largest and the smallest chromosomes was found equal to 1.82. The long arm of the most asymmetric chromosome was more than twice longer than the short arm. According to these parameters, we assigned Pompia to the 2A class of karyotype asymmetry class [
34].
The Total Haploid Length of chromosome (THL) was estimated as 162.25
m. The centromeric asymmetry and mean centromeric asymmetry were 9.94 and 18.54, respectively. The chromosome lengths showed a rather uniform distribution and the coefficient of variation of chromosome length (CVCL) was estimated in 18.85. For a complete list of estimated karyotype asymmetric indices see
Supplementary File 1.
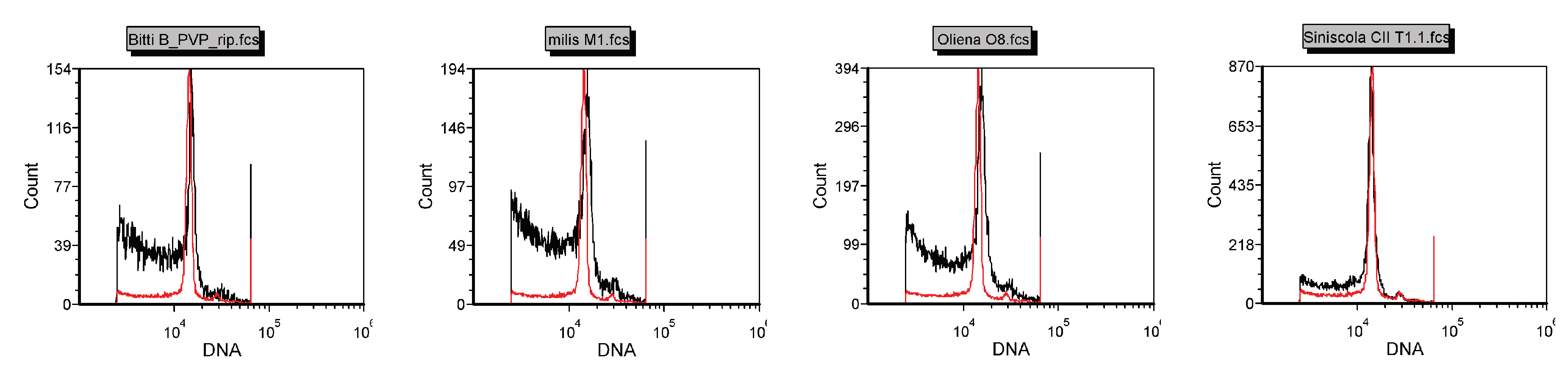
The nuclear DNA content of Pompia accessions was measured by flow cytometry analysis using
C. limon (1C = 0.40; pg [
35]) as diploid reference. No significant differences between the DNA content estimates of
C. limon and Pompia accessions were detected (
Figure 2).
Altogether, karyological and flow cytometrical data suggest that Pompia is a regular diploid citrus fruit tree.
2.2. Evolutionary Divergence of Pompia and Citrus Species Based on Single Nucleotide Polymorphisms at Nuclear or Cytoplasmic Loci
Molecular investigations were carried out on 10 Pompia accessions (
Table 2) along with accessions of the three basic Citrus species:
C. medica,
C. maxima,
C. reticulata and five secondary species, including
C. bergamia,
C. aurantium,
C. micrantha,
C. sinensis and
C. limon (
Table 2), which based on available morphological data [
18], could be considered among the most informative for Pompia taxonomic investigations (
Table 2). The citron Rhobs el arsa (ICVN0110244) is a
Citrus spp. accession morphologically similar to Pompia.
Preliminary investigations performed using the cpDNA sequences available in BOLD v4 [
37] for the two canonical barcodes rbcL and matK revealed a very low nucleotide variation and polymorphism rate between
C. medica,
C. limon and
C. aurantium [
27,
28,
29,
30]. We therefore focused our investigations on the intergenic (psbA-trnH) and intronic (trnL) cpDNA regions, together with the nuclear ITS locus, chosen as an nuclear marker for studying nucleotide diversity. The cpDNA amplicons ranged from 442 bp to 468 bp and from 530 bp to 542 bp, for the psbA-trnH and trnL-intron barcodes, respectively. The size of ITS amplicons ranged from 533 bp to 564 bp. Overall, the mean genetic distance was equal to 0.007 for psbA-trnH and 0.002 for trnL-intron sequences. The mean genetic distance for the ITS marker sequences was 0.010. The alignments of nucleotide sequences for the nuclear ITS and cpDNA regions are provided in the
Supplementary Files 2 and 3, respectively. Pairwise estimates of evolutionary divergence between taxa are shown in
Table 3.
As expected, the three Citrus basic species scored high levels of evolutionary divergence [
38].
C. medica (p: 0.004) was the closest to Pompia, followed by
C. maxima (p: 0.010) and
C. reticulata (p: 0.022). Since ITS sequences in several Citrus spp. accessions were associated with heterozygous loci, we subtracted the mean distance within groups from the average distance between groups (
Table 3). By doing so, the average net distance of Pompia was estimated as 0.019 from
C. reticulata and 0.007 from
C. maxima, while the net distance between Pompia and
C. medica was equal to 0.001. In relation to secondary Citrus species, Pompia showed the lowest level of ITS sequence divergence from the citron Rhobs el arsa (average net distance: −0.003) and
C. bergamia (average net distance: −0.003). The cp-DNA markers did not resolve univocally the genetic relatedness between Pompia and the basic species. The trnL marker identified
C. medica as the closest basic species to Pompia and based on the psbA-trnH marker this species scored the highest distance from Pompia.
C. maxima was the closest basic species to Pompia based on psbA-trnH marker while based on the trnL marker this species together with
C. reticulata proved the most divergent from Pompia among the basic species.
Regarding the pairwise comparisons involving Pompia and secondary Citrus species, both cpDNA marker loci indicated complete identity between Pompia and Rhobs el arsa,
C. bergamia,
C. aurantium and
C. limon and an average distance of 0.002 with
C. sinensis. The distance between Pompia and
C. micrantha was 0.002 for trnL and 0.014 for psbA-trnH (
Table 4). The ITS multiple sequence alignment displayed a stretch of 29 contiguous gaps starting from the nucleotide position 213 in sequences amplified from Pompia,
C. limon,
C. medica and
C. bergamia (see
Supplemental File 2). These observations prompted us to look for haplotypes discriminating the analysed species. The region corresponding to the ITS1 provided 44 SNVs (Single Nucleotide Variants), 29 of which were In/Dels whereas the region corresponding to the ITS2 revealed 5 SNPs and a single In/Del at position 451 of the alignment (
Table 5).
A total of nine haplotypes were identified, including six species-specific haplotypes and two haplotypes common to multiple accessions. More in detail, Hap1 was identified in all Pompia accessions, Rhobs el arsa and
C. aurantium, while Hap8 was found in Pompia accessions, Rhobs el arsa,
C. medica and
C. bergamia (
Table 5). Noteworthy, Hap1 was not identified in basic Citrus species. Furthermore, it is considered that
C. bergamia (Accession: SRA 212) is heterozygous for the two alleles best described by Hap5 and Hap8, while Rhobs el arsa (Accession: ICVN0110033) share the haplotypes Hap1 and Hap8 with all investigated accessions of Pompia. The occurrence of the observed In/Del in position 213–241 of the sequence alignment, was confirmed in all investigated Citrus spp. accessions, by using specific primers overlapping this In/Del (
Table 5). With this respect, it is worth noting that all Pompia accessions were confirmed heterozygous for this locus. Furthermore, consistent amplification profiles were observed for the accessions sharing one or multiple haplotypes with Pompia, namely:
C. medica (ACC: SRA540, SRA101, Hap8) and Rhobs el arsa (Acc. ICVN0110244; Hap8) ,
C. bergamia (Acc. SRA 612; Hap 5,8) and
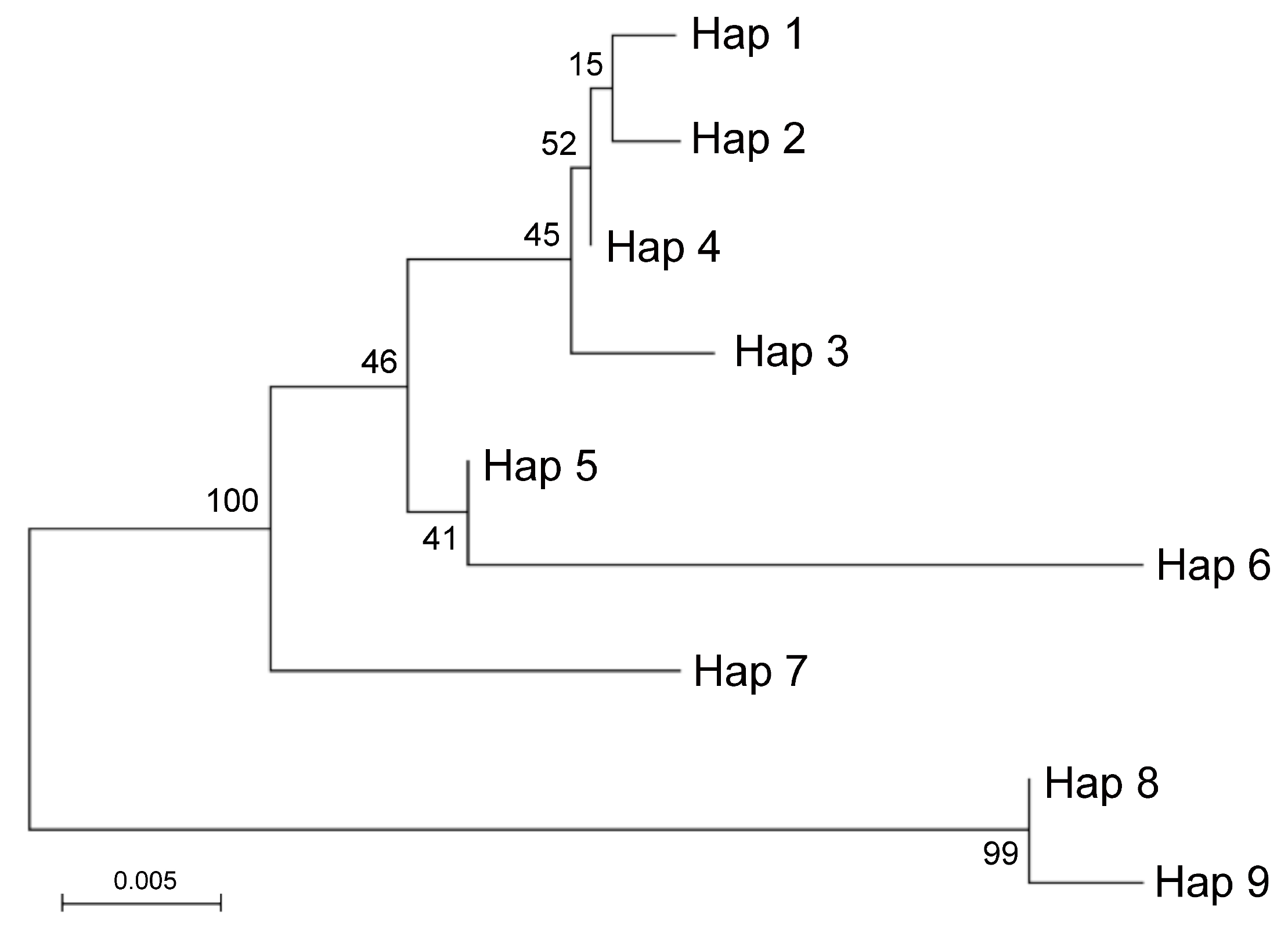
C. aurantium (Acc. ICVN0110033; Hap1) and Rhobs el arsa (Acc. ICVN0110244; Hap1) . The phylogenetic relationships between haplotypes of the ITS locus were summarized in a Maximum likelihood tree (
Figure 3).
Two main groups including highly similar haplotypes were identified. One group included Hap8 identified in
C. medica and Pompia and Hap9 identified in
C. limon. The other included: Hap1 identified in Pompia, Rhobs el arsa and
C. aurantium, which clustered in proximity to Hap2 (
C. sinensis), Hap4 (
C. maxima) and Hap3 (
C. sinensis). Hap5 identified
C. bergamia and Hap6 identified in
C.reticulata formed a subgroup with lower similarity. Finally, Hap7 specific of
C. micrantha was in intermediate position between this latter groups. This picture was in substantial agreement with Curk et al. [
38]. Based on ITS data it is likely that Pompia inherited the Hap8 from
C. medica or
C. bergamia and that Hap1 found in Pompia accessions derived from
C. aurantium or one of its interspecific hybrids. Moreover, at the level of resolution allowed by these molecular analyses, Pompia and Rhobs el arsa seem to be a case of synonymy.
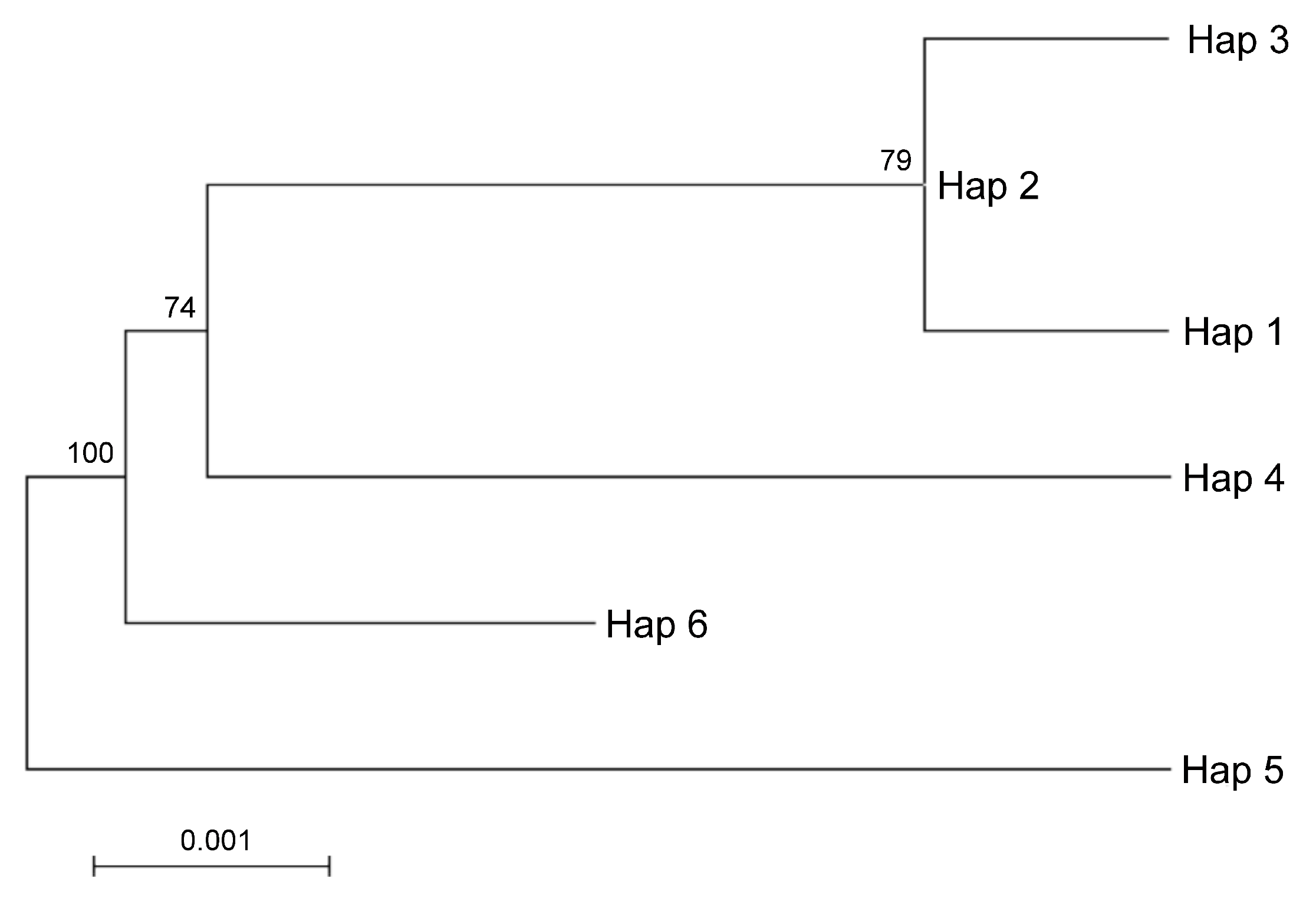
Taking into account the maternal inheritance of the chloroplast genome, we integrated the above nuclear DNA findings with chloroplast DNA polymorphisms in order to distinguish between parental species contributions. The analysis of the intergenic region psbA-trnH revealed 51 SNVs, 40 of which were In/Dels. The trnL-intron was less informative as it provided 4 SNPs and 12 In/Dels (for a total of 16 SNVs). Interestingly, the SNVs analysis of the merged chloroplast data set revealed six haplotypes (
Appendix A and
Supplementary File 3). It is worth noting that a single haplotype, namely Hap1, contained the sequences found in Pompia, together with Rhobs el arsa,
C. aurantium and the interspecific hybrids originated from this latter secondary species:
C. limon,
C. bergamia. Unique haplotypes were found for the remaining species:
C. maxima (Hap3),
C. sinensis (Hap2),
C. medica (Hap6),
C. reticulata (Hap5) and
C. micrantha (Hap4). Regarding the relationships among cpDNA haplotypes, the Maximum likelihood tree clustered Hap1 in proximity of Hap3 and Hap2 and far more distantly from Hap 4 (
Figure 4). The haplotypes displaying higher evolutionary divergence from Pompia (Hap1) accessions were Hap6 and Hap5, respectively from
C. medica and
C. reticulata.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}



