The Power of Genomic in situ Hybridization (GISH) in Interspecific Breeding of Bulb Onion (Allium cepa L.) Resistant to Downy Mildew (Peronospora destructor [Berk.] Casp.)

 , ,
, ,
Abstract
:1. Introduction
2. Results
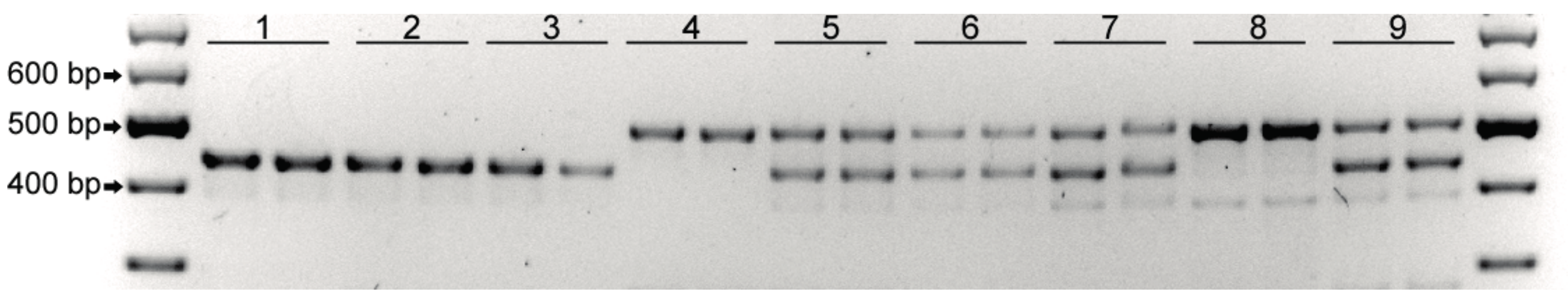
2.1. DMR1 marker-assisted screening
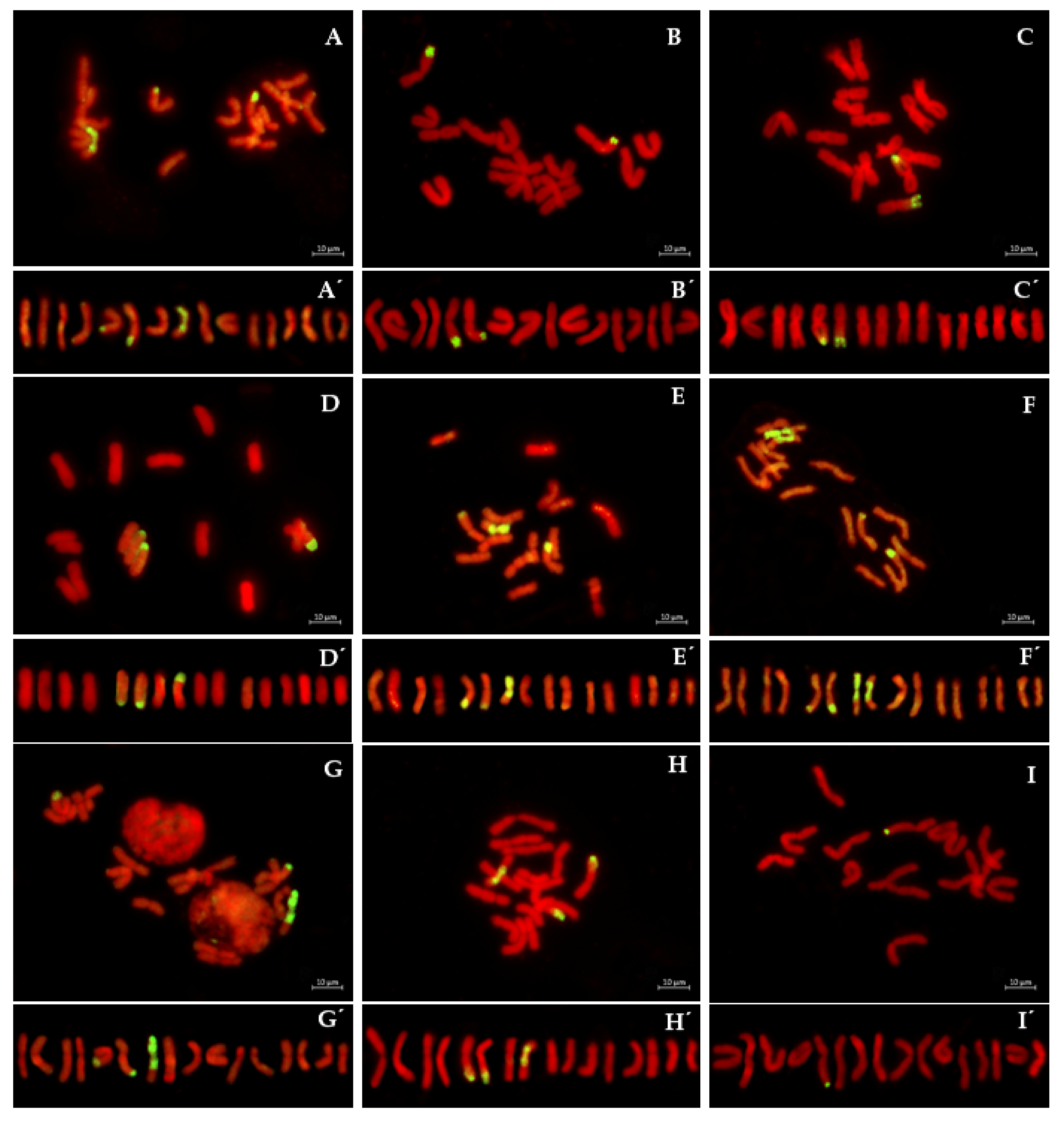
2.2. GISH Analysis
2.2.1. S1BC2
2.2.2. S2BC2
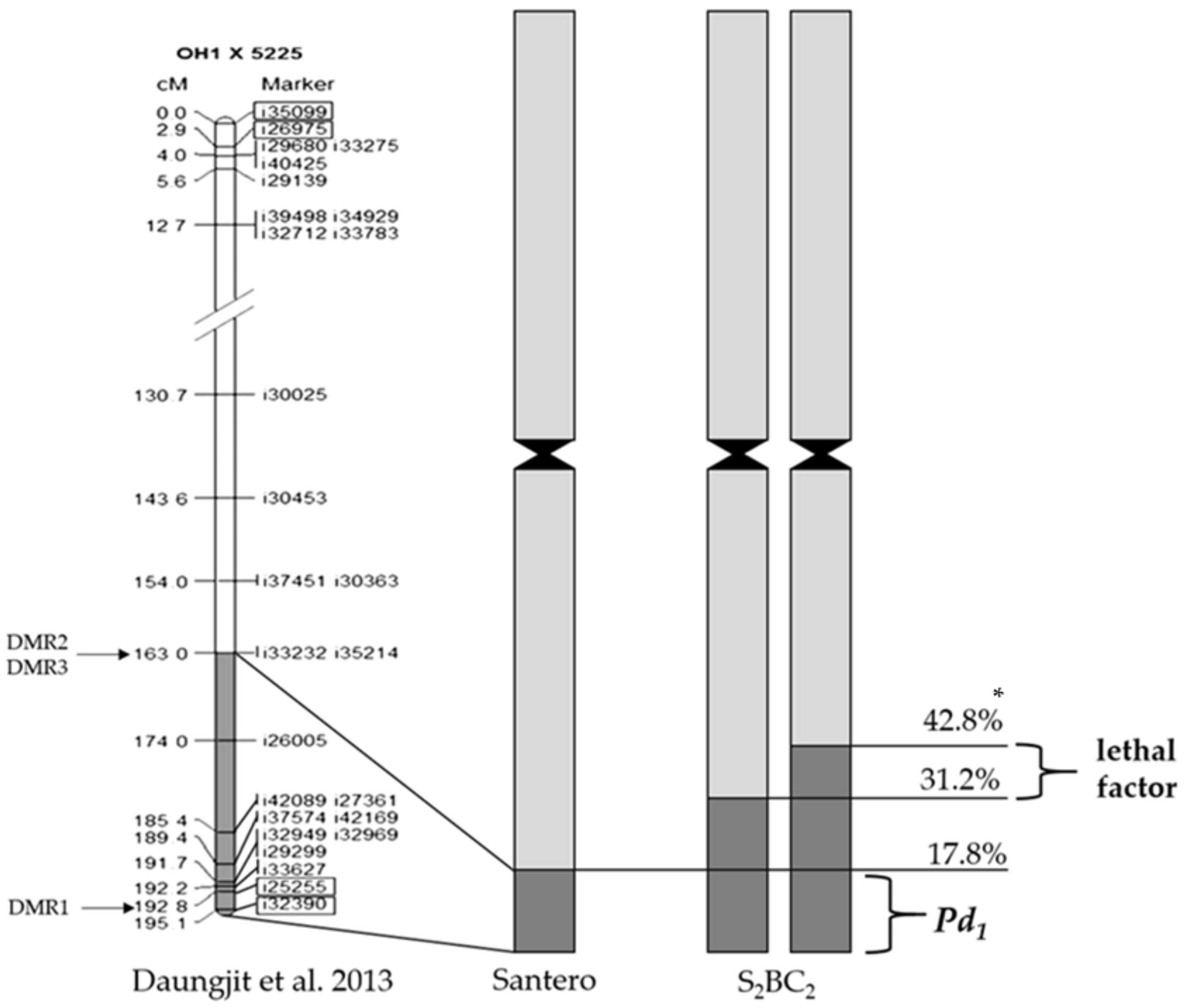
2.2.3. Monitoring of Crossing over Events between the Recombinant and A. cepa Chromosomes
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. PCR Analysis with DMR1
4.3. Preparation of Mitotic Chromosomes
4.4. Genomic in Situ Hybridization (GISH)
4.5. Microscopy and Image Analysis and Karyotype Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schwarzacher, T.; Leitch, A.R.; Bennett, M.D.; Heslop-Harrison, J.S. In-situ localization of parental genomes in a wide hybrid. Ann. Bot. 1989, 64, 315–324. [Google Scholar] [CrossRef]
- Dong, F.; Novy, R.G.; Helgeson, J.P.; Jiang, J. Cytological characterization of potato-Solanum etuberosum somatic hybrids and their backcross progenies by genomic in situ hybridization. Genome 1999, 42, 987–992. [Google Scholar] [CrossRef]
- Schwarzacher, T. DNA, chromosomes and in situ hybridization. Genome 2003, 46, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Khrustaleva, L.I.; de Melo, P.E.; van Heusden, A.W.; Kik, C. The integration of recombination and physical maps in a large-genome monocots using haploid genome analysis in a trihybrid Allium population. Genetics 2005, 169, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Budylin, M.V.; Kan, L.Y.; Romanov, V.S.; Khrustaleva, L.I. GISH Study of Advanced Generation of the Interspecific Hybrids Between Allium cepa L. and Allium fistulosum L. with Relative Resistance to Downy Mildew. Russ. J. Genet. 2014, 50, 387–394. [Google Scholar] [CrossRef]
- Ramzan, F.; Younis, A.; Lim, K.B. Application of genomic in situ hybridization in horticultural science. Int. J. Genomics 2017, 2017, 7561909. [Google Scholar] [CrossRef] [PubMed]
- Hizume, M. Allodiploid nature of Allium wakegi Araki revealed by genomic in situ hybridization and localization of 5S and 18S rDNAs. Jpn. J. Genet. 1994, 69, 407–415. [Google Scholar] [CrossRef]
- Khrustaleva, L.I.; Kik, C. Cytogenetical studies in the bridge cross Allium cepa × (A. fistulosum × A. roylei). Theor. Appl. Genet. 1998, 96, 8–14. [Google Scholar] [CrossRef]
- Khrustaleva, L.I.; Kik, C. Introgression of Allium fistulosum into A. cepa mediated by A. roylei. Theor. Appl. Genet. 2000, 10, 17–26. [Google Scholar] [CrossRef]
- Zwierzykowski, Z.; Zwierzykowska, E.; Taciak, M.; Jones, N.; Kosmala, A.; Krajewski, P. Chromosome pairing in allotetraploid hybrids of Festuca pratensis × Lolium perenne revealed by genomic in situ hybridization (GISH). Chromosome Res. 2008, 16, 575–585. [Google Scholar] [CrossRef]
- Silva, G.S.; Souza, M.M. Genomic in situ hybridization in plants. Genet. Mol. Res. 2013, 12, 2953–2965. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef]
- Schwartz, H.F.; Mohan, S.K. Compendium of Onion and Garlic Diseases and Pests, 2nd ed.; APS Press: St. Paul, MN, USA, 2008; pp. 32–35. [Google Scholar]
- Scholten, O.E.; Van Heusden, A.W.; Khrustaleva, L.I.; Burger-Meijer, K.; Mank, R.A.; Antonise, R.G.C.; Harrewijn, J.L.; Van haecke, W.; Oocst, E.H.; Peters, R.J.; et al. The long and winding road leading to the successful introgression of downy mildew resistance into onion. Euphytica 2007, 156, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.N. Cytogenetic evolution in the genus Allium. In Cytogenetics of Crop Plants; Swaminathan, M.S., Gupta, P.K., Sinha, U., Eds.; MacMillan: New York, NY, USA, 1983; pp. 516–554. [Google Scholar]
- Kofoet, A.; Zinkernagel, V. Resistance to downy mildew (Peronospora destructor (Berk.) Casp.) in Allium species. J. Plant Dis. Protect. 1990, 97, 13–23. [Google Scholar]
- Kofoet, A.; Kik, C.; Wietsma, W.A.; De Vries, J.N. Inheritance of resistance to downy mildew (Peronospora destructor [Berk.] Casp.) from Allium roylei Stearn in the backcross Allium cepa L. × (A. roylei × A. cepa). Plant Breed. 1990, 105, 144–149. [Google Scholar] [CrossRef]
- De Vries, J.N.; Jongerius, R.; Sandbrink, H.; Lindhout, P. RAPD markers assist in resistance breeding. Prophyta 1992, 2, 50–51. [Google Scholar] [CrossRef]
- Kik, C.; Buiteveld, J.; Verbeek, W.H.J. Biotechnological aspects of Allium breeding. Acta. Hortic. 1997, 433, 291–297. [Google Scholar] [CrossRef]
- Van Heusden, A.W.; Van Ooijen, J.W.; Vrielink-van Ginkel, R.; Verbeek, W.H.J.; Wietsma, W.A.; Kik, C. A genetic map of an interspecific cross in Allium based on amplified fragment length polymorphism (AFLPTM) markers. Theor. Appl. Genet. 2000, 100, 118–126. [Google Scholar] [CrossRef]
- Van Heusden, A.W.; Shigyo, M.; Tashiro, Y.; Vrielink-van Ginkel, R.; Kik, C. AFLP linkage group assignment to the chromosomes of Allium cepa L. via monosomic addition lines. Theor. Appl. Genet. 2000, 100, 480–486. [Google Scholar] [CrossRef]
- Kim, S.; Kim, C.W.; Choi, M.S.; Kim, S. Development of a simple PCR marker tagging the Allium roylei fragment harboring resistance to downy mildew (Peronospora destructor) in onion (Allium cepa L.). Euphytica 2016, 208, 561–569. [Google Scholar] [CrossRef]
- Duangjit, J.; Bohanec, B.; Chan, A.P.; Town, C.D.; Havey, M.J. Transcriptome sequencing to produce SNP-based genetic maps of onion. Theor. Appl. Genet. 2013, 126, 2093–2101. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, M.; Kim, Y.; Yeom, S.; Cheong, K.; Kim, K.; Jeon, J.; Kim, S.; Kim, D.; Sohn, S.; et al. Integrative structural annotation of de novo RNA-Seq provides an accurate reference gene set of the enormous genome of the onion (Allium cepa L.). DNA Res. 2015, 22, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Breton, C.; Serieys, H.; Bervillé, A. Gene transfer from wild Helianthus to sunflower: Topicalities and limits. Ol. Corps Gras Lipides 2010, 17, 104–114. [Google Scholar] [CrossRef]
- Kumar, N.V.M. Introgression of genes by wide hybridization: A crop improvement. Int. J. Appl. Pure Sci. Agric. 2016, 2, 33–38. [Google Scholar]
- Niranjana, M. Gametocidal genes of Aegilops: Segregation distorters in wheat–Aegilops wide hybridization. Genome 2017, 60, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Eshed, Y.; Zamir, D. A genomic library of Lycopersicon pennellii in L. esculentum: A tool for fine mapping of genes. Euphytica 1994, 79, 175–179. [Google Scholar] [CrossRef]
- Monforte, A.J.; Tanksley, S.D. Fine mapping of a quantitative trait locus (QTL) from Lycopersicon hirsutum chromosome 1 affecting fruit characteristics and agronomic traits: Breaking linkage among QTLs affecting different traits and dissection of heterosis for yield. Theor. Appl. Genet. 2000, 100, 471–479. [Google Scholar] [CrossRef]
- Francis, D.M.; Kabelka, E.; Bell, J.; Franchino, B.; St-Clair, D. Resistance to bacterial canker in tomato (Lycopersicon hirsutum LA407) and its progeny derived from crosses to L. esculentum. Plant Dis. 2001, 85, 1171–1176. [Google Scholar] [CrossRef]
- Canady, M.A.; Meglic, V.; Chetleat, R.T. A library of Solanum lycopersicoides introgression lines in cultivated tomato. Genome 2005, 48, 685–697. [Google Scholar] [CrossRef]
- Emsweller, S.L.; Jones, H.A. An interspecific hybrid in Allium. Hilgardia 1935, 9, 265–273. [Google Scholar] [CrossRef] [Green Version]
- De Vries, J.N.; Wietsma, W.A.; Jongerius, M.C. Linkage of downy mildew resistance genes Pd1 and Pd2 from Allium roylei Stearn in progeny of its interspecific hybrid with onion (A. cepa). Euphytica 1992, 64, 131–137. [Google Scholar] [CrossRef]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol. 1985, 5, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Schwarzacher, T.; Heslop-Harrison, P. Practical In Situ Hybridization; BIOS Scientific Ltd.: Oxford, UK, 2000; p. 203. [Google Scholar]
- De Vries, J.N. Onion chromosome nomenclature and homoeology relationships—Workshop report. Euphytica 1990, 49, 1–3. [Google Scholar] [CrossRef]
- De Vries, J.N.; Jongerius, M.C. Interstitial C-bands on the chromosomes of Allium-species from section Cepa. In Proceedings of the 4th Eucarpia Allium Symposium, Wellesbourne, UK, 6–9 September 1988; Institute of Horticultural Research: Bengaluru, India, 1988; pp. 71–78. [Google Scholar]
- Kirov, I.; Khrustaleva, L.; Van Laere, K.; Soloviev, A.; Meeus, S.; Romanov, D.; Fesenko, I. DRAWID: User-friendly java software for chromosome measurements and idiogram drawing. Comp. Cytogenet. 2017, 11, 747–757. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Position of recombination site*, % | ||||
|---|---|---|---|---|---|
| Chromosome 3 | Chromosome 4 | ||||
| Homologous 1 (large fragment) Mean ± SD | Homologous 2 (small fragment) Mean ± SD | Homologous 1 Mean ± SD | Homologous 2 Mean ± SD | ||
| S1BC2-8 (parental form) | 42.8 ± 2.25 | 22.3 ± 1.73 | A. cepa** | 54.0L ± 1.73 | |
| S2BC2 | 8-5 | 38.6 ± 1.38 | 20.3 ± 0.69 | A. cepa | 54.0L ± 1.73 |
| 8-20 | 24.2 ± 1.73 | 20.9 ± 1.21 | A. cepa | 44.5S ± 2.76 | |
| 8-32 | 41.5 ± 1.20 | 31.2 ± 1.00 | A. cepa | 54.0L ± 2.00 | |
| 8-33 | 39.7 ± 1.56 | 22.5 ± 4.02 | A. cepa | A. cepa | |
| 8-37 | 42.4 ± 1.56 | 22.5 ± 0.69 | A. cepa | 54.0L ± 1.73 | |
| 8-38 | 39.1 ± 1.00 | 27.0 ± 1.40 | 54.0L ± 2.00 | 46.2S ± 2.60 52.4L ± 2.40 | |
| 8-40 | 38.2 ± 2.25 | 24.7 ± 1.56 | A. cepa | 54.0L ± 1.73 | |
| 8-45 | 39.4 ± 1.60 | 24.1 ± 1.40 | A. cepa | 54.0L ± 2.00 | |
| 8-53 | 42.3 ± 1.56 | 24.1 ± 1.65 | A. cepa | A. cepa | |
| 8-56 | 39.0 ±1.00 | 24.2 ± 2.00 | A. cepa | 51.0S ± 2.00 54.6L ± 2.00 | |
| 8-59 | 41.8 ± 1.56 | 23.0 ± 1.38 | A. cepa | A. cepa | |
| Santero | A. cepa | 17.8 ± 1.90 | A. cepa | A. cepa | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khrustaleva, L.; Mardini, M.; Kudryavtseva, N.; Alizhanova, R.; Romanov, D.; Sokolov, P.; Monakhos, G. The Power of Genomic in situ Hybridization (GISH) in Interspecific Breeding of Bulb Onion (Allium cepa L.) Resistant to Downy Mildew (Peronospora destructor [Berk.] Casp.). Plants 2019, 8, 36. https://doi.org/10.3390/plants8020036
Khrustaleva L, Mardini M, Kudryavtseva N, Alizhanova R, Romanov D, Sokolov P, Monakhos G. The Power of Genomic in situ Hybridization (GISH) in Interspecific Breeding of Bulb Onion (Allium cepa L.) Resistant to Downy Mildew (Peronospora destructor [Berk.] Casp.). Plants. 2019; 8(2):36. https://doi.org/10.3390/plants8020036
Chicago/Turabian StyleKhrustaleva, Ludmila, Majd Mardini, Natalia Kudryavtseva, Rada Alizhanova, Dmitry Romanov, Pavel Sokolov, and Grigory Monakhos. 2019. "The Power of Genomic in situ Hybridization (GISH) in Interspecific Breeding of Bulb Onion (Allium cepa L.) Resistant to Downy Mildew (Peronospora destructor [Berk.] Casp.)" Plants 8, no. 2: 36. https://doi.org/10.3390/plants8020036